Сотовый переход - Cell junction
| Сотовый переход | |
|---|---|
| Подробности | |
| Идентификаторы | |
| латинский | межклеточные соединения |
| TH | H1.00.01.0.00012 |
| FMA | 67394 |
| Анатомическая терминология | |
Клеточные соединения (или же межклеточные мосты[1]) представляют собой класс ячеистых структур, состоящих из мультипротеиновые комплексы которые обеспечивают контакт или адгезию между соседними клетки или между ячейкой и внеклеточный матрикс у животных. Они также поддерживают межклеточный барьер эпителий и контроль парацеллюлярный транспорт. Клеточные соединения особенно многочисленны в эпителиальных тканях. В сочетании с молекулы клеточной адгезии и внеклеточный матрикс, соединения клеток помогают удерживать клетки животных вместе.
Клеточные соединения также особенно важны для обеспечения связи между соседними клетками через специализированные белковые комплексы, называемые сообщающиеся (щелевые) переходы. Клеточные соединения также важны для снижения нагрузки на клетки.
На заводах подобные каналы связи известны как плазмодесматы, И в грибы они называются септальные поры.[2]
Типы

В позвоночные, существует три основных типа соединений клеток:
- Адгезивные соединения, десмосомы и гемидесмосомы (анкерные соединения)
- Щелевые соединения[3] (сообщающийся узел)
- Узкие стыки (перекрывающие соединения)
Беспозвоночные есть несколько других типов конкретных соединений, например перегородки или C. elegans апикальное соединение.
В многоклеточных растения, структурные функции клеточных соединений вместо этого обеспечиваются клеточные стенки. Аналоги коммуникативных соединений клеток у растений получили название плазмодесматы.
Анкерные соединения
Клетки в тканях и органах должны быть прикреплены друг к другу и прикреплены к компонентам внеклеточный матрикс. Клетки разработали несколько типов соединительных комплексов для выполнения этих функций, и в каждом случае якорные белки проходят через плазматическую мембрану, связывая цитоскелетные белки в одной клетке с белками цитоскелета в соседних клетках, а также с белками внеклеточного матрикса.[4]
Наблюдаются три типа якорных соединений, которые отличаются друг от друга якорем цитоскелетного белка, а также трансмембранным линкерным белком, который проходит через мембрану:
| Соединение | Цитоскелетный якорь | Трансмембранный линкер | Связывает ячейку с: |
|---|---|---|---|
| Десмосомы | Промежуточные нити | Кадгерин | Другие клетки |
| Гемидесмосомы | Промежуточные нити | Интегрины | Матрица ЕС |
| Адгезивные соединения | Актин нити | Кадгерин / Интегрины | Другие ячейки / Матрица ЕС |
Соединения якорного типа не только удерживают клетки вместе, но и обеспечивают структурную сплоченность тканей. Эти соединения наиболее многочисленны в тканях, которые подвергаются постоянным механическим нагрузкам, например, в коже и сердце.[4]
Десмосомы

Десмосомы, также называемые прикрепленными пятнами, можно визуализировать как заклепки, проходящие через плазматическая мембрана соседних ячеек. Промежуточные нити состоит из кератин или же десмин прикрепляются к ассоциированным с мембраной белкам прикрепления, которые образуют плотную бляшку на цитоплазматической стороне мембраны. Молекулы кадгерина образуют фактический якорь, прикрепляясь к цитоплазматической бляшке, проходя через мембрану и прочно связываясь с кадгеринами, проходящими через мембрану соседней клетки.[5]
Гемидесмосомы
Гемидесмосомы образуют заклепочные связи между цитоскелетом и компонентами внеклеточного матрикса, такими как базальные пластинки которые лежат в основе эпителия. Подобно десмосомам, они связываются с промежуточными филаментами в цитоплазме, но в отличие от десмосом их трансмембранные якоря представляют собой интегрины, а не кадгерины.[6]
Адгезивные соединения
Адгезивные соединения имеют общие характеристики якорения клеток через их цитоплазматические актиновые нити. Подобно десмосомам и гемидесмосомам, их трансмембранные якоря состоят из кадгеринов в тех, которые прикрепляются к другим клеткам, и интегринов в тех, которые прикрепляются к внеклеточному матриксу. Значительный морфологическое разнообразие среди стыковок. Те, которые связывают клетки друг с другом, видны как отдельные полосы или пятна, или как полосы, которые полностью окружают клетку. Связки адгезивного типа связаны с пучками актиновых филаментов, которые также окружают клетку чуть ниже плазматической мембраны. Пятнообразные сращения помогают клеткам прикрепляться к внеклеточному матриксу как in vivo и in vitro где они называются очаговые спайки. Актиновые филаменты цитоскелета, которые связываются с адгезивными соединениями, являются сократительными белками, и в дополнение к обеспечению якорной функции, адгезивные соединения, как полагают, участвуют в складывании и изгибании листов эпителиальных клеток. Представление о полосках актиновых филаментов как о `` завязках '' позволяет представить, как сокращение полос внутри группы клеток могло бы искажать лист в виде интересных узоров.[4]
Сообщающиеся (щелевые) переходы
Сообщающиеся узлы, или щелевые соединения обеспечивают прямую химическую связь между соседней клеточной цитоплазмой посредством диффузии без контакта с внеклеточной жидкостью.[7] Это возможно благодаря шести коннексин белки взаимодействуют, образуя цилиндр с порой в центре, называемой коннексон.[8] Комплексы коннексонов простираются через клеточную мембрану, и когда два соседних клеточных коннексона взаимодействуют, они образуют полный канал щелевого соединения.[7][8] Поры коннексона различаются по размеру, полярности и, следовательно, могут быть специфичными в зависимости от белков коннексина, которые составляют каждый отдельный коннексон.[7][8] Хотя изменения в каналах щелевых контактов действительно происходят, их структура остается относительно стандартной, и это взаимодействие обеспечивает эффективную связь без выхода молекул или ионов во внеклеточную жидкость.[8]
Щелевые соединения играют жизненно важную роль в организме человека,[9] включая их роль в равномерном сокращении сердечная мышца.[9] Они также важны при передаче сигналов в мозг, а их отсутствие указывает на снижение плотности клеток в головном мозге.[10] Сетчатка и клетки кожи также зависят от щелевых контактов при дифференцировке и пролиферации клеток.[9][10]
Узкие стыки
Найдено у позвоночных эпителий плотные контакты действуют как барьеры, регулирующие движение воды и растворенных веществ между слоями эпителия. Узкие стыки классифицируются как околеклеточный барьер, который определяется как не имеющий направленной дискриминации; однако движение растворенного вещества в значительной степени зависит от размера и заряда. Есть основания полагать, что структуры, через которые проходят растворенные вещества, чем-то напоминают поры.
Физиологический pH играет роль в селективности прохождения растворенных веществ через плотные контакты, при этом наиболее плотные контакты являются немного селективными для катионов. Плотные соединения, присутствующие в разных типах эпителия, являются селективными для растворенных веществ разного размера, заряда и полярности.
Белки
Было идентифицировано около 40 белков, участвующих в плотных контактах. Эти белки можно разделить на четыре основные категории; каркасные белки, сигнальные белки, регулирующие белки, и трансмембранные белки.
Роли
- Белки каркаса - организовать трансмембранные белки, соединить трансмембранные белки с другими цитоплазматическими белками, а также с актиновыми филаментами.
- Сигнальные белки - участвует в сборке соединений, регуляции барьера и транскрипции генов.
- Регулирующие белки - регулируют нацеливание на мембранные везикулы.
- Трансмембранные белки - включая соединительная молекула адгезии, окклюдин, и Клодин.
Считается, что клаудин - это молекула белка, отвечающая за избирательную проницаемость между слоями эпителия.
Трехмерное изображение еще предстоит получить, и как таковая конкретная информация о функции плотных контактов еще не определена.
Трехклеточные соединения
Трехклеточные соединения запечатывают эпителий по углам трех клеток. Из-за геометрии вершин из трех ячеек, герметизация ячеек на этих участках требует специфической организации соединений, отличной от таковой в двухклеточных соединениях. У позвоночных компонентами трехклеточных соединений являются трицеллюлин и липопротеиновые рецепторы, стимулированные липолизом. У беспозвоночных компонентами являются глиотактин и анаконда.[11]
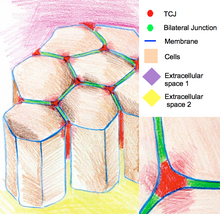
Трехклеточные соединения также участвуют в регуляции организации цитоскелета и клеточных делений. В частности, они обеспечивают деление клеток в соответствии с Правило Хертвига. В некоторых эпителиях дрозофилы во время деления клеток трехклеточные соединения устанавливают физический контакт с шпиндельный аппарат через астральные микротрубочки. Трехклеточные соединения оказывают тянущее усилие на аппарат веретена и служат геометрическими ключами для определения ориентации клеточных делений.[12]
Молекулы клеточного соединения
Молекулы, ответственные за создание клеточных соединений, включают различные молекулы клеточной адгезии. Выделяют четыре основных типа: селектины, кадгерины, интегрины, а суперсемейство иммуноглобулинов.[13]
Selectins представляют собой молекулы клеточной адгезии, которые играют важную роль в инициации воспалительных процессов.[14] Функциональная способность селектина ограничена взаимодействием лейкоцитов с эндотелием сосудов. У людей обнаружены три типа селектинов; L-селектин, P-селектин и E-селектин. L-селектин имеет дело с лимфоцитами, моноцитами и нейтрофилами, P-селектин работает с тромбоцитами и эндотелием, а E-селектин работает только с эндотелием. У них есть внеклеточные области, состоящие из аминоконцевого лектинового домена, присоединенного к углеводному лиганду, домена, подобного фактору роста, и коротких повторяющихся единиц (пронумерованные кружки), которые соответствуют доменам комплементарного связывающего белка.[15]
Кадгерины кальций-зависимые молекулы адгезии. Кадгерины чрезвычайно важны в процессе морфогенез – развитие плода. Вместе с альфа-бета катенин комплекса кадгерин может связываться с микрофиламентами цитоскелета клетки. Это обеспечивает гомофильную межклеточную адгезию.[16] В β-катенин –α-катенин связанный комплекс на стыках адгезивов позволяет формировать динамическую связь с актиновым цитоскелетом.[17]
Интегрины действуют как рецепторы адгезии, транспортируя сигналы через плазматическую мембрану во многих направлениях. Эти молекулы являются бесценной частью клеточной коммуникации, поскольку один лиганд может использоваться для многих интегринов. К сожалению, этим молекулам еще предстоит пройти долгий путь исследований.[18]
Суперсемейство иммуноглобулинов представляют собой группу кальций-независимых белков, способных к гомофильной и гетерофильной адгезии. Гомофильная адгезия включает связывание иммуноглобулин-подобных доменов на клеточной поверхности с иммуноглобулин-подобными доменами на противоположной клеточной поверхности, тогда как гетерофильная адгезия вместо этого относится к связыванию иммуноглобулин-подобных доменов с интегринами и углеводами.[19]
Адгезия клеток - жизненно важный компонент организма. Потеря этой адгезии влияет на клеточную структуру, клеточное функционирование и связь с другими клетками и внеклеточным матриксом и может привести к серьезным проблемам со здоровьем и заболеваниям.
Рекомендации
- ^ Митчелл, Ричард Шеппард; Кумар, Винай; Аббас, Абул К .; Фаусто, Нельсон (2007). «Глава 13: Вставка по морфологии плоскоклеточного рака». Базовая патология Роббинса (8-е изд.). Филадельфия: Сондерс. ISBN 978-1-4160-2973-1.
- ^ Блумендаль, S; Кюк, У (январь 2013 г.). «Межклеточная коммуникация у растений, животных и грибов: сравнительный обзор». Die Naturwissenschaften. 100 (1): 3–19. Bibcode:2013NW .... 100 .... 3B. Дои:10.1007 / s00114-012-0988-z. PMID 23128987. S2CID 11991859.
- ^ Эндрю Л. Харрис; Даррен Локк (2009). Коннексины, Путеводитель. Нью-Йорк: Спрингер. п. 574. ISBN 978-1-934115-46-6.
- ^ а б c Ян Х.Х., Мрук Д.Д., Ли В.М., Ченг CY (2008). Перекрестная связь между тугими и фиксирующими соединениями - урок из яичек. Достижения экспериментальной медицины и биологии. 636. Нью-Йорк, Нью-Йорк: Springer-Verlag New York. стр.234–54. Дои:10.1007/978-0-387-09597-4_13. ISBN 978-0-387-79990-2. ЧВК 4080640. PMID 19856171.
- ^ Ли П.П., Ченг CY, Мрук Д.Д. (2011). Биология десмосомоподобного соединения - универсального якорного соединения и преобразователя сигнала в семенном эпителии.. Международный обзор клеточной и молекулярной биологии. 286. С. 223–69. Дои:10.1016 / B978-0-12-385859-7.00005-7. ISBN 9780123858597. ЧВК 4381909. PMID 21199783.
- ^ Гипсон И.К., Спурр-Мишо С.Дж., Тисдейл А.С. (апрель 1988 г.). «Гемидесмосомы и фиксирующие фибриллы коллагена появляются синхронно во время развития и заживления ран». Биология развития. 126 (2): 253–62. Дои:10.1016/0012-1606(88)90136-4. PMID 3350210.
- ^ а б c Эванс WH, Мартин PE (2002). «Щелевые соединения: структура и функции (Обзор)». Молекулярная мембранная биология. 19 (2): 121–36. Дои:10.1080/09687680210139839. PMID 12126230. S2CID 20806078.
- ^ а б c d Лампе П.Д., Лау А.Ф. (июль 2004 г.). «Влияние фосфорилирования коннексина на коммуникацию через щелевые контакты». Международный журнал биохимии и клеточной биологии. 36 (7): 1171–86. Дои:10.1016 / S1357-2725 (03) 00264-4. ЧВК 2878204. PMID 15109565.
- ^ а б c "Abstracts: Proceedings of the International Gap Junction Conference. 5–9 августа 2007 г. Эльсинор, Дания". Связь и адгезия клеток. 14 (6): 275–346. 2007. Дои:10.1080/15419060801891042. PMID 18392995.
- ^ а б Вэй CJ, Xu X, Lo CW (2004). «Коннексины и клеточная передача сигналов в развитии и болезни». Ежегодный обзор клеточной биологии и биологии развития. 20: 811–38. Дои:10.1146 / annurev.cellbio.19.111301.144309. PMID 15473861.
- ^ Байри С., Мисра Т., Сайед З.А., Батц Т., Шах Дж., Борил Л., Глашаузер Дж., Эгертер-Вильмсен Т., Мацат Т., Муссиан Б., Ув А., Лушниг С. (2015). «Белок с тремя повторами Anakonda контролирует образование эпителиальных трехклеточных соединений у дрозофилы». Клетка развития. 33 (5): 535–48. Дои:10.1016 / j.devcel.2015.03.023. PMID 25982676.
- ^ Босвельд Ф., Маркова О., Гирао Б., Мартин К., Ван З., Пьер А., Балакирева М., Гауг I, Эйнсли А., Кристофору Н., Любенский Д. К., Минк Н., Беллаиш И. (2016). «Эпителиальные трехклеточные соединения действуют как сенсоры формы межфазных клеток, ориентируя митоз». Природа. 530 (7591): 496–8. Bibcode:2016Натура.530..495Б. Дои:10.1038 / природа16970. ЧВК 5450930. PMID 26886796.
- ^ Лодиш; и другие. (2007). Молекулярная клеточная биология (6-е изд.). В. Х. Фриман и компания. п. 803. ISBN 978-1429203142.
- ^ Теддер Т.Ф., Стибер Д.А., Чен А., Энгель П. (июль 1995 г.). «Селектины: молекулы сосудистой адгезии». Журнал FASEB. 9 (10): 866–73. Дои:10.1096 / fasebj.9.10.7542213. PMID 7542213.
- ^ Bevilacqua MP, Нельсон RM (февраль 1993 г.). "Селекты". Журнал клинических исследований. 91 (2): 379–87. Дои:10.1172 / JCI116210. ЧВК 287934. PMID 7679406.
- ^ Rowlands TM, Symonds JM, Farookhi R, Blaschuk OW (январь 2000 г.). «Кадгерины: важнейшие регуляторы структуры и функции репродуктивных тканей». Отзывы о репродукции. 5 (1): 53–61. Дои:10.1530 / revreprod / 5.1.53. PMID 10711736.
- ^ Brembeck FH, Rosário M, Birchmeier W (февраль 2006 г.). «Баланс клеточной адгезии и передачи сигналов Wnt, ключевая роль β-катенина». Текущее мнение в области генетики и развития. 16 (1): 51–9. Дои:10.1016 / j.gde.2005.12.007. PMID 16377174.
- ^ Хайнс РО (сентябрь 2002 г.). «Интегрины: двунаправленные, аллостерические сигнальные машины». Клетка. 110 (6): 673–87. Дои:10.1016 / S0092-8674 (02) 00971-6. PMID 12297042. S2CID 30326350.
- ^ Вай Вонг К., Дай ДЭ, Кумб Д.Р. (2012). «Роль молекул адгезии клеток суперсемейства иммуноглобулинов в метастазировании рака». Международный журнал клеточной биологии. 2012: 1–9. Дои:10.1155/2012/340296. ЧВК 3261479. PMID 22272201.
3. Из CCH (2010)
внешняя ссылка
- Альбертс, Брюс; Джонсон, Александр; Льюис, Джулиан; Рафф, Мартин; Робертс, Кейт; Уолтер, Питер (2002). «Сотовые соединения». Молекулярная биология клетки (4-е изд.). Нью-Йорк: Наука о гирляндах. ISBN 978-0-8153-3218-3.
- Межклеточные + соединения в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- Ячейка-матрица + переходы в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
| Авторитетный контроль |
|---|