Каркасный белок - Scaffold protein
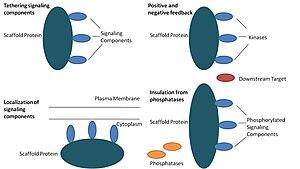
В биологии каркасные белки являются важными регуляторами многих ключевых сигнальные пути. Хотя функции каркасов строго не определены, известно, что они взаимодействуют и / или связываются с множеством членов сигнального пути, привязывая их к комплексы. В таких путях они регулируют передачу сигнала и помогают локализовать компоненты пути (организованные в комплексы) в определенных областях клетки, таких как плазматическая мембрана, то цитоплазма, то ядро, то Гольджи, эндосомы, а митохондрии.
История
Первым обнаруженным сигнальным каркасным белком был Ste5 белок из дрожжей Saccharomyces cerevisiae. Было показано, что три различных домена Ste5 связаны с протеинкиназы Ste11, Ste7, и Fus3 с образованием мультикиназного комплекса.[2]
Функция
Белки каркаса действуют по крайней мере четырьмя способами: связывая компоненты передачи сигналов, локализуя эти компоненты в определенных областях клетки, регулируя передачу сигналов путем координации положительный и негативный отзыв сигналы и изоляция правильных сигнальных белков от конкурирующих белков.[1]
Компоненты сигнализации привязки
Эта конкретная функция считается самой основной функцией каркаса. Эшафоты собирают сигнальные компоненты каскад в комплексы. Эта сборка может повышать специфичность передачи сигналов, предотвращая ненужные взаимодействия между белками передачи сигналов, и повышать эффективность передачи сигналов за счет увеличения близости и эффективной концентрации компонентов в комплексе каркаса. Типичным примером того, как каркасы повышают специфичность, является каркас, который связывает протеинкиназу и ее субстрат, обеспечивая тем самым специфическое фосфорилирование киназы. Кроме того, некоторые сигнальные белки требуют множественных взаимодействий для активации, и привязка каркаса может быть способна преобразовывать эти взаимодействия в одно взаимодействие, которое приводит к множественным модификациям.[3][4] Каркасы также могут быть каталитическими, поскольку взаимодействие с сигнальными белками может приводить к аллостерический изменения этих сигнальных компонентов.[5] Такие изменения могут усиливать или ингибировать активацию этих сигнальных белков. Примером является каркас Ste5 в митоген-активированной протеинкиназе (MAPK ) путь. Предполагается, что Ste5 направляет передачу сигналов спаривания через Fus3 MAPK путем каталитического разблокирования этой конкретной киназы для активации ее MAPKK Ste7.[6]
Локализация сигнальных компонентов в клетке
Каркасы локализуют сигнальную реакцию в определенной области клетки, и этот процесс может быть важным для местного производства сигнальных промежуточных соединений. Конкретным примером этого процесса является каркас, якорные белки A-киназы (AKAP), которые нацелены на циклическую AMP-зависимую протеинкиназу (PKA ) на различные сайты в ячейке.[7] Эта локализация способна локально регулировать PKA и приводит к локальному фосфорилированию PKA ее субстратов.
Согласование положительных и отрицательных отзывов
Многие гипотезы о том, как каркасы координируют положительную и отрицательную обратную связь, исходят от инженерных каркасов и математического моделирования. В сигнальных каскадах трех киназ каркасы связывают все три киназы, повышая специфичность киназы и ограничивая амплификацию сигнала за счет ограничения фосфорилирования киназы только одной нижерасположенной мишенью.[3][8][9] Эти способности могут быть связаны со стабильностью взаимодействия между каркасом и киназами, базальным фосфатаза активность в клетке, расположение каркаса и уровни экспрессии сигнальных компонентов.[3][8]
Изоляция правильных сигнальных белков от инактивации
Сигнальные пути часто инактивируются ферментами, которые меняют состояние активации и / или вызывают деградацию сигнальных компонентов. Были предложены каркасы для защиты активированных сигнальных молекул от инактивации и / или деградации. Математическое моделирование показало, что киназы в каскаде без каркасов имеют более высокую вероятность дефосфорилирования фосфатазами еще до того, как они смогут фосфорилировать нижестоящие мишени.[8] Кроме того, было показано, что каркасы изолируют киназы от субстратных и АТФ-конкурентных ингибиторов.[10]
Резюме белка каркаса
| Белки каркаса | Путь | Возможные функции | Описание |
|---|---|---|---|
| KSR | MAPK | Сборка и локализация пути RAS-ERK | Одним из наиболее изученных сигнальных путей в биологии является РАС-ЭРК путь, в котором G-белок RAS активирует MAPKKK РАФ, который активирует MAPKK MEK1 (MAPK / ERK киназа 1), которая затем активирует MAPK ERK. Было идентифицировано несколько каркасных белков, участвующих в этом пути и других подобных путях MAPK. Одним из таких каркасных белков является KSR, который является наиболее вероятным эквивалентом хорошо изученного дрожжевого каркасного белка Ste5 MAPK.[11] Он является положительным регулятором пути и связывает многие белки этого пути, включая все три киназы в каскаде.[6] Было показано, что KSR локализуется на плазматической мембране во время активации клетки, тем самым играя роль в сборке компонентов пути ERK и в локализации активированной ERK на плазматической мембране.[12] |
| MEKK1 | MAPK | Сборка и локализация сигнаносомы рецептора смерти | Другие белки каркаса включают В-клеточную лимфому 10 (BCL-10 ) и МЕК-киназа 1 (MEKK1 ), которые играют роль в N-концевой киназе JUN (JNK ) путь. |
| BCL-10 | MAPK | Сборка и специфика JNK | |
| AKAP | PKA Пути | Координация фосфорилирования PKA по нижестоящим мишеням | Это семейство белков связано только структурно по своей способности связывать регуляторную субъединицу PKA, но в остальном может связывать очень разнообразный набор ферментов и субстратов. |
| АХНАК-1 | Сигнализация кальция | Сборка и локализация кальциевых каналов | Передача сигналов кальция необходима для правильного функционирования иммунных клеток. Недавние исследования показали, что белок каркаса, AHNAK1, важен для эффективной передачи сигналов кальция и NFAT активация в Т-клетки благодаря его способности правильно локализовать кальциевые каналы на плазматической мембране [14]. В неиммунных клетках AHNAK1 также может связывать кальциевые каналы с фосфолипазой Cγ (PLC-γ ) и PKC.[1] Белки, связывающие кальций, часто подавляют большую часть поступающего кальция, поэтому связывание этих эффекторов кальция может быть особенно важным, когда сигналы индуцируются слабым притоком кальция. |
| ГОМЕР | Сигнализация кальция | Подавление активации NFAT | Другой пример белка каркаса, который модулирует передачу сигналов кальция, - это белки семейства HOMER. Было показано, что белки HOMER конкурируют с кальциневрин связываться с N-концом NFAT в активированных Т-клетках.[13] Благодаря этой конкуренции белки HOMER могут снижать активацию NFAT, что также снижает производство Ил-2 цитокин.[13] Напротив, белки HOMER также, как было показано, положительно регулируют передачу сигналов кальция в нейронах, связывая рецептор глутамата с рецепторами трифосфата в эндоплазматическом ретикулуме.[14] |
| Пеллино | Врожденная иммунная сигнализация | Сборка сигнаносомы TLR | Существуют доказательства того, что белки пеллино функционируют как каркасные белки в важном сигнальном пути врожденного иммунитета, Toll-подобном рецепторе (TLR ) путь. Во многом функция Пеллино является предположением; однако белки Pellino могут связываться с IRAK1, TRAF6 и TAK1 после активации IL-1R, указывая на то, что они могут собирать и локализовать компоненты пути TLR рядом с его рецептором.[15][16] |
| NLRP | Врожденная иммунная сигнализация | Сборка инфламмасомы | Семейство NLR - это высококонсервативное и большое семейство рецепторов, участвующих в врожденном иммунитете. Семейство рецепторов NLRP (семейство NLR, содержащие пириновый домен) функционируют как каркасы, собирая инфламмасому, комплекс, который приводит к секреции провоспалительных цитокинов, таких как IL-18 и IL-1β.[17] |
| DLG1 | Рецептор Т-клеток сигнализация | Сборка и локализация сигнальных молекул TCR, активация p38 | DLG1 является высококонсервативным в иммунных клетках и важен для активации Т-клеток на периферии. Он задействуется в иммунологическом синапсе и связывает ζ-цепь Т-клеточного рецептора (TCR ) в CBL, WASP, p38, LCK, VAV1 и ZAP70.[18][19][20][21] Эти данные предполагают, что DLG1 играет роль в связывании сигнального аппарата TCR с регуляторами цитоскелета, а также предполагает роль в альтернативной активации пути p38. Однако неясно, регулирует ли DLG1 положительно или отрицательно активацию Т-клеток. |
| Спинофилин | Дендритная клетка сигнализация | Сборка иммунологических белков синапсов DC | Спинофилин участвует в дендритный клеточная функция специфически участвует в формировании иммунологических синапсов. Спинофилин рекрутируется в синапс после контакта дендритных клеток с Т-клетками. Этот набор кажется важным, потому что без спинофилина дендритные клетки не могут активировать Т-клетки. in vitro или же in vivo.[22] Как спинофилин способствует презентации антигена в этом случае, все еще неизвестно, хотя возможно, что спинофилин регулирует продолжительность контакта клетки в синапсе или регулирует рециркуляцию костимулирующих молекул в клетке, таких как молекулы MHC.[1] |
| Регуляторный белок гриппа растений[23] | Согласование отрицательных отзывов во время протохлорофиллид биосинтез. | Сборка и локализация пути, который включает синтез высокотоксичных веществ. протохлорофиллид, предшественник хлорофилл. | Синтез протохлорофиллида должен строго регулироваться, поскольку для его превращения в хлорофилл требуется свет. Регуляторный белок гриппа находится в тилакоид мембрана и содержит только несколько сайтов белок-белкового взаимодействия без каталитической активности. Мутанты, лишенные этого белка, накапливают протохлорофиллид в темноте. Партнеры по взаимодействию неизвестны. В ходе эволюции белок претерпел упрощение. |
Белок Хантингтин
Хантингтин белок совмещен с Банкомат восстановить белок на участках Повреждение ДНК.[24] Хантингтин представляет собой каркасный белок в комплексе ответа на окислительное повреждение ДНК ATM.[24] Болезнь Хантингтона пациенты с аберрантным белком хантингтина испытывают дефицит в восстановлении окислительное повреждение ДНК. Окислительное повреждение ДНК, по-видимому, лежит в основе болезни Хантингтона патогенез.[25] Болезнь Хантингтона, вероятно, вызвана дисфункцией мутантного каркасного белка хантингтина в Ремонт ДНК что приводит к усилению окислительного повреждения ДНК в метаболически активных клетках.[24]
Другое использование термина "белок каркаса"
В некоторых других случаях в биологии (не обязательно в отношении передачи сигналов клеток) термин «каркасный белок» используется в более широком смысле, когда белок объединяет несколько вещей для любых целей.
- В сворачивании хромосом
- Хромосомный каркас играет важную роль в удержании хроматина в компактном хромосома. Хромосомный каркас состоит из белков, в том числе конденсин, топоизомераза IIα и член семейства кинезинов 4 (KIF4)[26] Белки, входящие в состав хромосомного каркаса, также называют каркасным белком.
- В ферментативной реакции
- Большие многофункциональные ферменты, которые выполняют ряд или цепочку реакций общим путем, иногда называемые каркасными белками.[27] такие как пируватдегидрогеназа.
- В формировании формы молекулы
- Фермент или структурный белок, который удерживает несколько молекул вместе, чтобы удерживать их в правильном пространственном расположении, например, каркасные белки кластера серы железа.[28][29]
- Строительные леса
- В цитоскелет и ECM молекулы обеспечивают механический каркас. Например, коллаген 4-го типа[30]
Рекомендации
- ^ а б c d Шоу, Андрей С .; Филберт, Эрин Л. (январь 2009 г.). «Белки каркаса и передача сигналов иммунных клеток». Nature Reviews Иммунология. 9 (1): 47–56. Дои:10.1038 / nri2473. PMID 19104498. S2CID 13443447.
- ^ Чой, Кан-Йелл; Саттерберг, Бретт; Lyons, David M .; Элион, Элейн А. (август 1994 г.). «Ste5 связывает несколько протеинкиназ в каскаде MAP-киназ, необходимых для спаривания в С. cerevisiae". Клетка. 78 (3): 499–512. Дои:10.1016/0092-8674(94)90427-8. PMID 8062390. S2CID 20541545.
- ^ а б c Левченко, Андре; Брук, Иошуа; Штернберг, Пол В. (23 мая 2000 г.). «Белки каркаса могут двухфазно влиять на уровни передачи сигналов митоген-активируемой протеинкиназы и снижать ее пороговые свойства». Труды Национальной академии наук. 97 (11): 5818–5823. Bibcode:2000PNAS ... 97.5818L. Дои:10.1073 / pnas.97.11.5818. ЧВК 18517. PMID 10823939.
- ^ Феррелл, Джеймс Э. (3 октября 2000 г.). «Что на самом деле делают белки каркаса?». Научная сигнализация. 2000 (52): pe1. Дои:10.1126 / стке.522000pe1. S2CID 219192522.
- ^ Бурак, У. Ричард; Шоу, Андрей С (апрель 2000). «Передача сигнала: висит на эшафоте». Текущее мнение в области клеточной биологии. 12 (2): 211–216. Дои:10.1016 / S0955-0674 (99) 00078-2. PMID 10712921.
- ^ а б Хорошо, Мэтью; Тан, Грейс; Синглтон, Джули; Ременьи, Аттила; Лим, Венделл А. (март 2009 г.). «Каркас Ste5 направляет передачу сигналов спаривания путем каталитической разблокировки киназы Fus3 MAP для активации». Клетка. 136 (6): 1085–1097. Дои:10.1016 / j.cell.2009.01.049. ЧВК 2777755. PMID 19303851.
- ^ Вонг, Вэй; Скотт, Джон Д. (декабрь 2004 г.). «Сигнальные комплексы АКАП: фокусы в пространстве и времени». Обзоры природы Молекулярная клеточная биология. 5 (12): 959–970. Дои:10.1038 / nrm1527. PMID 15573134. S2CID 15268680.
- ^ а б c Locasale, Джейсон У .; Шоу, Андрей С .; Чакраборти, Аруп К. (14 августа 2007 г.). «Белки каркаса придают каскады протеинкиназ разнообразные регуляторные свойства». Труды Национальной академии наук. 104 (33): 13307–13312. Bibcode:2007ПНАС..10413307Л. Дои:10.1073 / pnas.0706311104. PMID 17686969. S2CID 8907943.
- ^ Ухлик, Марк Т; Эйбелл, Эми Н; Куэвас, Брюс Д; Накамура, Казухиро; Джонсон, Гэри Л. (1 декабря 2004 г.). «Электросхемы регулирования МАПК по МЕКК1, 2 и 3». Биохимия и клеточная биология. 82 (6): 658–663. Дои:10.1139 / o04-114. PMID 15674433.
- ^ Гринвальд, Эрик С .; Редден, Джон М .; Dodge-Kafka, Kimberly L .; Соцерман, Джеффри Дж. (24 января 2014 г.). «Переключение состояния каркаса усиливает, ускоряет и изолирует передачу сигналов протеинкиназы C». Журнал биологической химии. 289 (4): 2353–2360. Дои:10.1074 / jbc.M113.497941. ЧВК 3900978. PMID 24302730.
- ^ Clapéron, A .; Терриен, М. (май 2007 г.). «KSR и CNK: два каркаса, регулирующие RAS-опосредованную активацию RAF». Онкоген. 26 (22): 3143–3158. Дои:10.1038 / sj.onc.1210408. PMID 17496912. S2CID 31061333.
- ^ Мюллер, Юрген; Ори, Стефан; Коупленд, Терри; Пивница-Вормс, Хелен; Моррисон, Дебора К. (ноябрь 2001 г.). «C-TAK1 регулирует передачу сигналов Ras путем фосфорилирования каркаса MAPK, KSR1». Молекулярная клетка. 8 (5): 983–993. Дои:10.1016 / S1097-2765 (01) 00383-5. PMID 11741534.
- ^ а б Huang, Guo N .; Хусо, Дэвид Л .; Буяен, Самуэль; Ту, Цзяньчэнь; McCorkell, Kelly A .; Мэй, Майкл Дж .; Чжу, Юйвэнь; Лутц, Майкл; Коллинз, Сэмюэл; Дехофф, Марлин; Канг, Шин; Уортенби, Кэтрин; Пауэлл, Джонатан; Лихи, Дэниел; Уорли, Пол Ф. (25 января 2008 г.). «Связывание NFAT и регуляция активации Т-клеток цитоплазматическими белками Гомера». Наука. 319 (5862): 476–481. Bibcode:2008Sci ... 319..476H. Дои:10.1126 / science.1151227. ЧВК 3602998. PMID 18218901.
- ^ Сяо, Бо; Ченг Ту, Цзянь; Уорли, Пол Ф (июнь 2000 г.). «Гомер: связь между нервной активностью и функцией рецептора глутамата». Текущее мнение в нейробиологии. 10 (3): 370–374. Дои:10.1016 / S0959-4388 (00) 00087-8. PMID 10851183. S2CID 8699597.
- ^ Цзян, Чжэнфань; Джонсон, Х. Ян; Не, Хуэйцин; Цинь, Цзиньчжун; Птица, Тимоти А .; Ли, Сяося (28 марта 2003 г.). «Пеллино 1 необходим для передачи сигналов, опосредованной интерлейкином-1 (IL-1), благодаря его взаимодействию с комплексом киназы 4, ассоциированной с рецептором IL-1 (IRAK4) -IRAK-рецептор фактора некроза опухоли, фактора 6 (TRAF6)». Журнал биологической химии. 278 (13): 10952–10956. Дои:10.1074 / jbc.M212112200. PMID 12496252. S2CID 10165785.
- ^ Ю, Кан-Ёль; Квон, Хён-Джу; Норман, Дэвид А. М .; Виг, Ева; Goebl, Mark G .; Харрингтон, Морин А. (15 октября 2002 г.). "На переднем крае: мышь Pellino-2 модулирует передачу сигналов IL-1 и липополисахаридов". Журнал иммунологии. 169 (8): 4075–4078. Дои:10.4049 / jimmunol.169.8.4075. PMID 12370331. S2CID 25317655.
- ^ Петрилли, Вирджиния; Достерт, Кэтрин; Муруве, Даниэль А; Цхопп, Юрг (декабрь 2007 г.). «Инфламмасома: чувствительный к опасности комплекс, запускающий врожденный иммунитет». Текущее мнение в иммунологии. 19 (6): 615–622. Дои:10.1016 / j.coi.2007.09.002. PMID 17977705.
- ^ Ксавье, Рамник; Рабизаде, Шахруз; Исигуро, Казухиро; Андре, Нико; Ортис, Дж. Бернабе; Вахтель, Хизер; Моррис, Дэвид Дж .; Лопес-Иласака, Марко; Шоу, Альберт С .; Сват, Войцех; Сид, Брайан (19 июля 2004 г.). «Диски больших (Dlg1) комплексов в активации лимфоцитов». Журнал клеточной биологии. 166 (2): 173–178. Дои:10.1083 / jcb.200309044. ЧВК 2172307. PMID 15263016.
- ^ Ханада, Тошихико; Линь, Луньхуэй; Чанди, К. Джордж; О, С. Стивен; Чишти, Атар Х. (24 октября 1997 г.). «Человеческий гомолог дисков дрозофилы, супрессор большой опухоли связывается с p56 lck тирозинкиназой и калиевым каналом Kv1.3 шейкерного типа в Т-лимфоцитах». Журнал биологической химии. 272 (43): 26899–26904. Дои:10.1074 / jbc.272.43.26899. PMID 9341123. S2CID 23446334.
- ^ Round, июнь L .; Хамфрис, Лиза А .; Томасян, Тамар; Миттельштадт, Пол; Чжан, Мин; Мичели, М. Кэрри (февраль 2007 г.). «Каркасный белок Dlgh1 координирует активацию альтернативной киназы p38, направляя сигналы Т-клеточного рецептора на NFAT, но не на факторы транскрипции NF-κB». Иммунология природы. 8 (2): 154–161. Дои:10.1038 / ni1422. PMID 17187070. S2CID 11906543.
- ^ Round, июнь L .; Томасян, Тамар; Чжан, Мин; Патель, Виреш; Schoenberger, Stephen P .; Мичели, М. Кэрри (7 февраля 2005 г.). «Dlgh1 координирует полимеризацию актина, синаптический Т-клеточный рецептор и агрегацию липидного рафта, а также эффекторную функцию Т-клеток». Журнал экспериментальной медицины. 201 (3): 419–430. Дои:10.1084 / jem.20041428. ЧВК 2213022. PMID 15699074.
- ^ Блум, Она; Unternaehrer, Julia J .; Цзян, Аймин; Шин, Чон-Сук; Деламар, Лелия; Аллен, Патрик; Меллман, Ира (21 апреля 2008 г.). «Спинофилин участвует в передаче информации в иммунологических синапсах». Журнал клеточной биологии. 181 (2): 203–211. Дои:10.1083 / jcb.200711149. PMID 18411312. S2CID 1717736.
- ^ Мескаускене, Раса; Натер, Мена; Гусят, Дэвид; Кесслер, Феликс; Лагерь, Роэль-оп-ден; Апель, Клаус (23 октября 2001 г.). «Грипп: негативный регулятор биосинтеза хлорофилла у Arabidopsis thaliana». Труды Национальной академии наук. 98 (22): 12826–12831. Bibcode:2001ПНАС ... 9812826М. Дои:10.1073 / pnas.221252798. ЧВК 60138. PMID 11606728.
- ^ а б c Майури, Тамара; Мокл, Эндрю Дж .; Hung, Claudia L .; Ся, Цзяньжун; van Roon-Mom, Willeke M.C .; Труант, Рэй (25 декабря 2016 г.). «Хантингтин представляет собой каркасный белок в комплексе ответа на окислительное повреждение ДНК ATM». Молекулярная генетика человека. 26 (2): 395–406. Дои:10.1093 / hmg / ddw395. PMID 28017939.
- ^ Аяла-Пенья, Сильветт (сентябрь 2013 г.). «Роль окислительного повреждения ДНК в митохондриальной дисфункции и патогенезе болезни Хантингтона». Свободная радикальная биология и медицина. 62: 102–110. Дои:10.1016 / j.freeradbiomed.2013.04.017. ЧВК 3722255. PMID 23602907.
- ^ Пунперм, Равин; Таката, Хидеаки; Хамано, Тору; Мацуда, Ацуши; Учияма, Сусуму; Хираока, Ясуши; Фукуи, Киичи (1 июля 2015 г.). «Хромосомный каркас представляет собой двухцепочечную сборку белков каркаса». Научные отчеты. 5 (1): 11916. Bibcode:2015НатСР ... 511916П. Дои:10.1038 / srep11916. ЧВК 4487240. PMID 26132639.
- ^ молекулярная клеточная биология от Lodish[требуется полная цитата ]
- ^ Аяла-Кастро, Карла; Шайни, Авниш; Ауттен, Ф. Уэйн (2008). «Пути сборки кластера Fe-S в бактериях». Обзоры микробиологии и молекулярной биологии. 72 (1): 110–125. Дои:10.1128 / MMBR.00034-07. ЧВК 2268281. PMID 18322036.
- ^ Адровер, Микель; Хоуз, Барри Д.; Яннуцци, Клара; Смулевич, Джульетта; Пасторе, Анналиса (1 июня 2015 г.). «Анатомия каркасного белка железо-серного кластера: понимание детерминант кластерной стабильности [2Fe – 2S] на IscU». Biochimica et Biophysica Acta (BBA) - Исследование молекулярных клеток. 1853 (6): 1448–1456. Дои:10.1016 / j.bbamcr.2014.10.023. PMID 25447544.
- ^ Молекулярная клеточная биология Lodish et al. выпуск 5[страница нужна ]