Коннексин - Connexin
| Коннексин | |||||||||
|---|---|---|---|---|---|---|---|---|---|
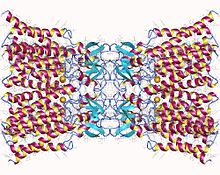 Додекамер коннексина-26. Щелевое соединение, состоящее из двенадцати идентичных белков коннексина, по шесть в мембране каждой клетки. Каждая из этих шести единиц представляет собой единственный полипептид, который четыре раза проходит через мембрану (называемые четырехпроходными трансмембранными белками). | |||||||||
| Идентификаторы | |||||||||
| Символ | Коннексин | ||||||||
| Pfam | PF00029 | ||||||||
| ИнтерПро | IPR013092 | ||||||||
| PROSITE | PDOC00341 | ||||||||
| TCDB | 1.A.24 | ||||||||
| OPM суперсемейство | 194 | ||||||||
| Белок OPM | 2zw3 | ||||||||
| |||||||||
Коннексины (Сх) (ТК № 1.A.24 ), или же щелевой переход белки, являются структурно родственными трансмембранными белками, которые собираются с образованием щелевых соединений позвоночных. Совершенно другое семейство белков, иннексины, образуют щелевые соединения в беспозвоночные.[1] Каждый щелевой переход состоит из двух полуканалов, или коннексоны, которые состоят из гомо- или гетерогексамерных массивов коннексинов, и коннексон в одной плазматической мембране стыкуется встык с коннексоном в мембране близко противоположной клетки. Гемиканал состоит из шести субъединиц коннексина, каждая из которых состоит из четырех трансмембранных сегментов. Щелевые соединения необходимы для многих физиологических процессов, таких как скоординированная деполяризация сердечная мышца, правильное эмбриональное развитие и проведенный ответ в микрососудистой сети. По этой причине мутации в генах, кодирующих коннексин, могут приводить к функциональным нарушениям и аномалиям развития.
Номенклатура
Коннексины обычно называют в соответствии с их молекулярной массой, например Cx26 представляет собой белок коннексин 26 кДа. Конкурирующая номенклатура - это белок щелевого соединения система, в которой коннексины сортируются по их формам α (GJA) и β (GJB), с дополнительными коннексинами, сгруппированными в группы C, D и E, за которыми следует идентификационный номер, например GJA1 соответствует Сх43. После голосования на конференции Gap Junction Conference (2007) в Эльсиноре сообщество согласилось использовать систему номенклатуры GJ для генов, кодирующих коннексины, но пожелало сохранить номенклатуру коннексинов для кодируемых белков, используя вес человеческого белка для нумерации. ортологичных белков.
Структура
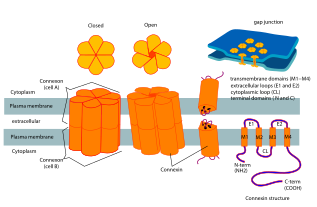
Коннексины содержат четыре высокоупорядоченных трансмембранные сегменты (TMS), в первую очередь неструктурированные цитоплазматические концы C и N, цитоплазматическая петля (CL) и две внеклеточные петли (EL-1) и (EL-2). Коннексины собираются в группы по шесть, чтобы сформировать полуканалы или коннексоны, а два полуканала затем объединяются, образуя щелевое соединение.
Доступна кристаллическая структура канала щелевого соединения, образованного Cx26 человека (также известного как GJB2) с разрешением 3,5 Å.[2] Карта плотности показывала два межмембранных полуканала и расположение четырех TMS шести протомеров, образующих каждый полуканал. Полуканалы имеют положительно заряженный вход в цитоплазму, воронку, отрицательно заряженный трансмембранный путь и внеклеточную полость. Пора сужается у воронки, которая образована шестью аминоконцевыми спиралями, выстилающими стенку канала, что, таким образом, определяет ограничение размера молекулы на входе в канал.
Семейство генов коннексина разнообразно: двадцать один идентифицированный член в секвенированном геноме человека и двадцать у мыши (девятнадцать из которых являются ортологичными парами). Обычно они весят от 25 до 60 кДа и имеют среднюю длину 380 аминокислот. Было обнаружено, что различные коннексины объединяются как в гомомерные, так и в гетеромерные щелевые переходы, каждый из которых может проявлять разные функциональные свойства, включая проводимость пор, селективность по размеру, селективность заряда, стробирование напряжения и химическое стробирование.[3]
Биосинтез и интернализация
Замечательный аспект коннексинов заключается в том, что у них относительно короткий период полураспада, составляющий всего несколько часов.[4] Результатом является наличие динамического цикла, в котором синтезируются и заменяются коннексины. Было высказано предположение, что такая короткая продолжительность жизни позволяет иметь более тонко регулируемые физиологические процессы, например, в миометрий.
От ядра к мембране
Поскольку они транслируются рибосомами, коннексины вставляются в мембрану эндоплазматический ретикулум (ER).[5] Именно в ER коннексины сворачиваются правильно, образуя две внеклеточные петли, EL-1 и EL-2. Также в ER олигомеризация молекул коннексина в полуканалы начинается, процесс, который может продолжаться также в промежуточном компартменте UR-Golgi.[4] Расположение этих гемиканалов может быть гомотипическим, гетеротипическим и комбинированным гетеротипическим / гетеромерным. После выхода из ER и проходя через ERGIC, свернутые коннексины обычно попадают в СНГ-Сеть Гольджи.[6] Однако некоторые коннексины, такие как Cx26, могут транспортироваться независимо от Гольджи.[7][8][9][10][11]
Сборка щелевого соединения
После внедрения в плазматическую мембрану клетки полуканалы свободно диффундируют в липидном бислое.[12] С помощью определенных белков, в основном кадгерины, полуканалы способны стыковаться с полуканалами соседних клеток, образуя щелевые соединения.[13] Недавние исследования показали наличие связи между адгезионными соединениями и щелевыми соединениями,[14] предполагая более высокий уровень координации, чем считалось ранее.
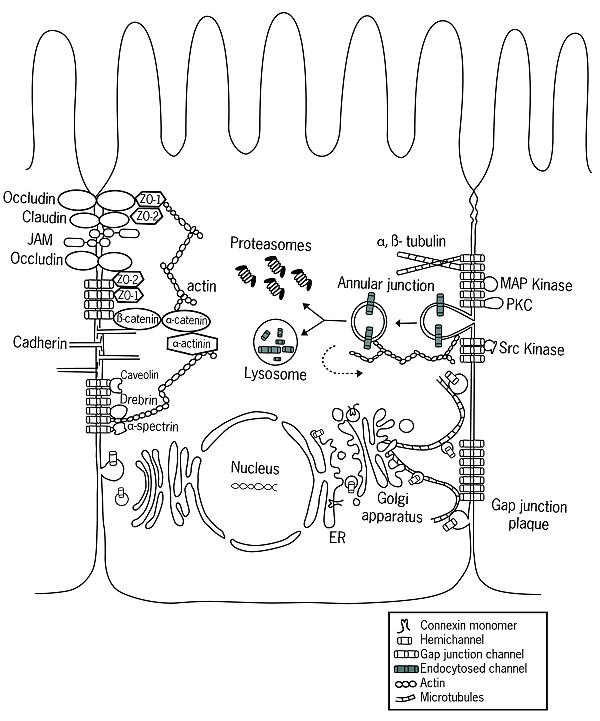
Функция
Щелевые соединения коннексинов встречаются только в позвоночные, в то время как функционально аналогичная (но генетически неродственная) группа белков, иннексины, отвечают за щелевые соединения в беспозвоночный разновидность. Иннексин ортологи также были идентифицированы в Хордовые, но они больше не могут образовывать щелевые контакты. Вместо этого каналы, образованные этими белками (называемые паннексины ) действуют как очень большие трансмембранные поры, которые соединяют внутри- и внеклеточные компартменты.
В рамках ЦНС щелевые соединения обеспечивают электрическую связь между клетками-предшественниками, нейронами и глиальными клетками. Используя специфический коннексин нокаутные мыши, исследования показали, что сцепление клеток важно для передачи визуальных сигналов. в сетчатка уровни внешнего освещения влияют на соединение ячеек, обеспечиваемое каналами щелевого соединения, адаптируя визуальную функцию к различным условиям освещения. Сцепление клеток регулируется несколькими механизмами, включая экспрессию коннексина.[16]
Decrock et al.. обсудили многоуровневую платформу, с помощью которой коннексины и паннексины могут влиять на следующие клеточные функции в ткани: (1) щелевые соединительные каналы коннексина (GJC) обеспечивают прямую межклеточную коммуникацию малых молекул, (2) гемиканалы коннексина и каналы паннексина могут способствовать к автокринный /паракринная передача сигналов путей, и (3) различные структурные домены этих белков позволяют выполнять независимые от каналов функции, такие как межклеточная адгезия, взаимодействие с цитоскелет и активация внутриклеточных сигнальных путей.[17] Таким образом, коннексины и паннексины вносят многогранный вклад в развитие мозга и специфические процессы в нейроглио-сосудистой единице, включая синаптическую передачу и пластичность, передачу глиальных сигналов, вазомоторный контроль и целостность гематоэнцефалического барьера в зрелой ЦНС.[17]
Специфичность субстрата
Различные коннексины могут проявлять разные специфичности в отношении растворенных веществ. Например, аденозин проходил примерно в 12 раз лучше через каналы, образованные Cx32, в то время как AMP и ADP прошел примерно в 8 раз лучше, и АТФ более чем в 300 раз лучше, через каналы, образованные Сх43. Таким образом, добавление фосфата к аденозину, по-видимому, смещает его относительную проницаемость с каналов, образованных Cx32, на каналы, образованные Cx43. Это может иметь функциональные последствия, поскольку энергетический статус клетки может контролироваться посредством экспрессии коннексина и образования каналов.[18]
Транспортная реакция
Транспортная реакция, катализируемая щелевыми контактами коннексина:
- Малые молекулы (цитоплазма клетки 1) ⇌ малые молекулы (цитоплазма клетки 2)
Коннексины человека и их влияние на болезнь
| Коннексин | Ген | Расположение и функции |
|---|---|---|
| Сх43 | GJA1 | Экспрессируется на поверхности сосудистой сети атеросклеротической бляшкой и активируется во время атеросклероза у мышей. Может иметь патологические эффекты. Также выражено между клетки гранулезы, который необходим для распространения. Обычно экспрессируется в астроцитах, также обнаруживается в большинстве астроцитом человека и в астроглиальном компоненте глионейрональных опухолей.[19] Это также главный кардиальный коннексин, обнаруживаемый в основном в миокарде желудочков.[20] Связана с окулодентодигитальная дисплазия. |
| Сх46 | GJA3 | |
| Сх37 | GJA4 | Индуцируется в гладких мышцах сосудов во время коронарного артериогенеза. Мутации Сх37 не смертельны. Образует щелевые контакты между ооцитами и клетками гранулезы и необходимы для выживания ооцитов. |
| Сх40 | GJA5 | Избирательно экспрессируется в миоцитах предсердий. Отвечает за координированную электрическую активацию предсердий.[21] |
| Сх33 | GJA6 (GJA6P) | Псевдоген в людях |
| Сх50 | GJA8 | Разрывные соединения между горизонтальными ячейками A-типа в сетчатке мыши и кролика[22] |
| Сх59 | GJA10 | |
| Сх62 | GJA10 | Cx62 человека соответствует Cx57 (мышь). Расположение в несущей аксон горизонтальной клетке B-типа в сетчатке кролика[23] |
| Сх32 | GJB1 | Основной компонент периферического миелина. Мутации в гене человека вызывают Х-сцепление Болезнь Шарко-Мари-Тута, наследственная невропатия. В нормальном мозге человека CX32 экспрессируется в нейронах и олигодендроцитах.[19] |
| Сх26 | GJB2 | Мутировал в Синдром Фохвинкеля а также Синдром кератита-иктиоза-глухоты (KID). |
| Сх31 | GJB3 | Может быть связан с Вариабильная эритрокератодермия. |
| Сх30.3 | GJB4 | Фонсека и другие. подтверждено выражение Cx30.3 в тимоциты.[24] Может быть связан с Вариабильная эритрокератодермия. |
| Сх31.1 | GJB5 | |
| Сх30 | GJB6 | Мутировал в Синдром Клустона (гидротическая эктодермальная дисплазия) |
| Сх25 | GJB7 | |
| Сх45 | GJC1 / GJA7 | Эпителиальные клетки протоков поджелудочной железы человека.[25] Атриовентрикулярный узел. |
| Сх47 | GJC2 / GJA12 | Экспрессируется в щелевых соединениях олигодентроцитов[26] |
| Сх31.3 | GJC3 | Человеческий ортолог мышиного Сх29. Не известно об образовании щелевых контактов.[27] |
| Cx36 | GJD2 / GJA9 | Функция бета-клеток поджелудочной железы, опосредующая высвобождение инсулина. Нейроны на протяжении Центральная нервная система где они синхронизируют нейронную активность.[28] |
| Сх31.9 | GJD3 / GJC1 | |
| Сх39 | GJD4 | |
| Сх40.1 | GJD4 | |
| Сх23 | GJE1 |
Щелевые соединения необходимы для многих физиологических процессов, таких как скоординированная деполяризация сердечная мышца, правильное эмбриональное развитие и проведенный ответ в микрососудистой сети. По этой причине делеция или мутация различных коннексинов изоформы производит отличительные фенотипы и патологии.[29] В то время как мутации в Сх43 в основном связаны с окулодентодигитальной дисплазией, мутации Сх47 связаны с Пелизей-Мерцбахер -подобное заболевание и лимфедема. Мутации Сх40 в основном связаны с фибрилляцией предсердий. Мутации в Cx37 еще не описаны, но полиморфизмы в гене Cx37 вовлечены в развитие артериальных заболеваний.[30]
Рекомендации
- ^ Лодиш, Харви Ф .; Арнольд Берк; Пол Мацудаира; Крис А. Кайзер; Монти Кригер; Мэтью П. Скотт; С. Лоуренс Зипурски; Джеймс Дарнелл (2004). Молекулярная клеточная биология (5-е изд.). Нью-Йорк: W.H. Фримен и компания. стр.230–1. ISBN 0-7167-4366-3.
- ^ Маэда С., Накагава С., Суга М., Ямасита Е., Осима А., Фудзиёси Ю., Цукихара Т. (апрель 2009 г.). «Структура канала щелевого соединения коннексина 26 при разрешении 3,5 А». Природа. 458 (7238): 597–602. Дои:10.1038 / природа07869. ISSN 1476-4687. PMID 19340074. S2CID 4431769.
- ^ Аяд В.А., Локк Д., Корин IV, Харрис А.Л. (июнь 2006 г.). «Гетеромерные, но не гомомерные каналы коннексина избирательно проницаемы для инозитолфосфатов». J. Biol. Chem. 281 (24): 16727–39. Дои:10.1074 / jbc.M600136200. ISSN 0021-9258. PMID 16601118.
- ^ а б Laird DW (март 2006 г.). «Жизненный цикл коннексинов в здоровье и болезни». Biochem. J. 394 (Pt 3): 527–43. Дои:10.1042 / BJ20051922. ЧВК 1383703. PMID 16492141.
- ^ Беннетт М.В., Зукин Р.С. (февраль 2004 г.). «Электрическая связь и нейронная синхронизация в мозге млекопитающих». Нейрон. 41 (4): 495–511. Дои:10.1016 / s0896-6273 (04) 00043-1. PMID 14980200. S2CID 18566176.
- ^ Musil, LS; Гуденаф Д.А. (1993). «Сборка мультисубъединиц интегрального белка канала плазматической мембраны, коннексина 43 щелевого соединения, происходит после выхода из ER». Клетка. 74 (6): 1065–77. Дои:10.1016/0092-8674(93)90728-9. PMID 7691412. S2CID 12169415.
- ^ Эванс У.Х., Ахмад С., Диз Дж, Джордж Ч., Кендалл Дж. М., Мартин П.Е. (1999). «Пути торговли людьми, ведущие к образованию щелевых контактов». Новартис найден. Symp. Симпозиумы Фонда Новартис. 219: 44–54, обсуждение 54–9. Дои:10.1002 / 9780470515587.ch4. ISBN 9780470515587. PMID 10207897.
- ^ Джордж CH, Кендалл JM, Эванс WH (март 1999). "Пути внутриклеточного трафика в сборке коннексинов в щелевые соединения". J. Biol. Chem. 274 (13): 8678–85. Дои:10.1074 / jbc.274.13.8678. PMID 10085106.
- ^ Джордж CH, Кендалл JM, Кэмпбелл AK, Evans WH (ноябрь 1998 г.). «Химеры коннексина-акворина сообщают о цитоплазматической кальциевой среде вдоль путей переноса, ведущих к биогенезу щелевых соединений в живых клетках COS-7». J. Biol. Chem. 273 (45): 29822–9. Дои:10.1074 / jbc.273.45.29822. PMID 9792698.
- ^ Мартин PE, Джордж CH, Кастро С., Кендалл Дж. М., Кейпел Дж., Кэмпбелл А. К., Ревилла А., Баррио Л. К., Эванс WH (январь 1998 г.). «Сборка химерных белков коннексин-экворин в функциональные каналы щелевых соединений. Отчетность о кальциевой среде внутриклеточной и плазматической мембраны». J. Biol. Chem. 273 (3): 1719–26. Дои:10.1074 / jbc.273.3.1719. PMID 9430718.
- ^ Мартин PE, Эррингтон RJ, Эванс WH (2001). «Сборка щелевого соединения: множественные флуорофоры коннексина идентифицируют сложные пути трафика». Cell Commun. Клеи. 8 (4–6): 243–8. Дои:10.3109/15419060109080731. PMID 12064596. S2CID 3029281.
- ^ Томас Т., Джордан К., Симек Дж., Шао К., Йедешко К., Уолтон П., Лэрд Д.В. (октябрь 2005 г.). «Механизмы транспорта Cx43 и Cx26 к плазматической мембране и регенерации щелевых контактов». J. Cell Sci. 118 (Pt 19): 4451–62. Дои:10.1242 / jcs.02569. PMID 16159960.
- ^ Джонген В. М., Фицджеральд Д. Д., Асамото М., Пикколи С., Слага Т. Джей, Грос Д., Такеичи М., Ямасаки Н. (август 1991 г.). «Регуляция межклеточной коммуникации через щелевые соединения, опосредованной коннексином 43, с помощью Ca2 + в эпидермальных клетках мышей контролируется E-кадгерином». J. Cell Biol. 114 (3): 545–55. Дои:10.1083 / jcb.114.3.545. ЧВК 2289094. PMID 1650371.
- ^ Вэй CJ, Фрэнсис Р., Сюй X, Ло CW (май 2005 г.). «Коннексин 43, связанный с N-кадгерин-содержащим мультибелковым комплексом, необходим для образования щелевых соединений в клетках NIH3T3» (PDF). J. Biol. Chem. 280 (20): 19925–36. Дои:10.1074 / jbc.M412921200. PMID 15741167. S2CID 770387.
- ^ Дбук Х.А., Мруе Р.М., Эль-Саббан М.Э., Талхук Р.С. (март 2009 г.). «Коннексины: множество функций, выходящих за рамки сборки каналов щелевых соединений». Сигнал сотовой связи. 7: 4. Дои:10.1186 / 1478-811X-7-4. ЧВК 2660342. PMID 19284610.
- ^ Кихара А.Х., де Кастро Л.М., Морискот А.С., Хамассаки, DE (май 2006 г.). «Длительная адаптация к темноте изменяет экспрессию коннексина в сетчатке мышей». J Neurosci Res. 83 (7): 1331–41. Дои:10.1002 / jnr.20815. PMID 16496335.
- ^ а б Decrock E, De Bock M, Wang N, Bultynck G, Giaume C, Naus CC, Green CR, Leybaert L (август 2015 г.). «Пути передачи сигналов коннексина и паннексина, архитектурный план физиологии и патологии ЦНС?». Клетка. Мол. Life Sci. 72 (15): 2823–51. Дои:10.1007 / s00018-015-1962-7. ISSN 1420-9071. PMID 26118660. S2CID 17170098.
- ^ Гольдберг Г.С., Морено А.П., Лампе П.Д. (сентябрь 2002 г.). «Щелевые соединения между клетками, экспрессирующими коннексин 43 или 32, демонстрируют обратную селективность по отношению к аденозину и АТФ». J. Biol. Chem. 277 (39): 36725–30. Дои:10.1074 / jbc.M109797200. ISSN 0021-9258. PMID 12119284.
- ^ а б Ароника Е., Гортер Дж. А., Янсен Г. Х., Леенстра С., Янкая Б., Трост Д. (май 2001 г.). «Экспрессия белков щелевых соединений коннексина 43 и коннексина 32 в опухолях мозга, связанных с эпилепсией, и в перилезионной эпилептической коре». Acta Neuropathol. 101 (5): 449–59. Дои:10.1007 / s004010000305. PMID 11484816. S2CID 6738913.
- ^ Verheule S, van Kempen MJ, te Welscher PH, Kwak BR, Jongsma HJ (май 1997 г.). «Характеристика каналов щелевого соединения в миокарде предсердий и желудочков взрослых кроликов». Circ. Res. 80 (5): 673–81. Дои:10.1161 / 01.res.80.5.673. PMID 9130448.
- ^ Голлоб М.Х., Джонс Д.Л., Кран А.Д., Дэнис Л., Гонг XQ, Шао К. и др. (Июнь 2006 г.). «Соматические мутации в гене коннексина 40 (GJA5) при фибрилляции предсердий». N. Engl. J. Med. 354 (25): 2677–88. Дои:10.1056 / NEJMoa052800. PMID 16790700.
- ^ Мэсси, Стивен (16 января 2009 г.). Коннексины: руководство (1-е изд.). Springer-Verlag Gmbh. С. 3– ?. ISBN 978-1-934115-46-6.
- ^ Бейер, Эрик С .; Причал, Вивиана М. (16 января 2009 г.). Коннексины: руководство (1-е изд.). Springer-Verlag Gmbh. С. 387–417. ISBN 978-1-934115-46-6.
- ^ Fonseca PC, Nihei OK, Urban-Maldonado M, Abreu S, de Carvalho AC, Spray DC, Savino W, Alves LA (июнь 2004 г.). «Характеристика коннексина 30.3 и 43 в тимоцитах». Иммунол. Латыш. 94 (1–2): 65–75. Дои:10.1016 / j.imlet.2004.03.019. PMID 15234537.
- ^ Тай М.Х., Олсон Л.К., Мадукар Б.В., Линнинг К.Д., Ван Кэмп Л., Цао М.С., Троско Д.Е. (январь 2003 г.). «Характеристика межклеточной коммуникации щелевого соединения в иммортализованных эпителиальных клетках протока поджелудочной железы человека с характеристиками стволовых клеток». Поджелудочная железа. 26 (1): e18–26. Дои:10.1097/00006676-200301000-00025. PMID 12499933. S2CID 34571252.
- ^ Камасава Н., Сик А., Морита М., Ясумура Т., Дэвидсон К.Г., Надь Джи, Раш Дж. Э. (2005). «Коннексин-47 и коннексин-32 в щелевых соединениях соматов олигодендроцитов, миелиновых оболочек, паранодальных петлях и резцах Шмидта-Лантермана: последствия для ионного гомеостаза и сифонирования калия». Неврология. 136 (1): 65–86. Дои:10.1016 / j.neuroscience.2005.08.027. ЧВК 1550704. PMID 16203097.
- ^ Саргианниду I, Ан М., Энрикес А.Д., Пейнадо А., Рейнольдс Р., Абрамс С., Шерер С.С., Клеопа К.А. (май 2008 г.). «Человеческие олигодендроциты экспрессируют Cx31.3: функция и взаимодействия с мутантами Cx32». Neurobiol. Дис. 30 (2): 221–33. Дои:10.1016 / j.nbd.2008.01.009. ЧВК 2704064. PMID 18353664.
- ^ Коннорс Б.В., Лонг Массачусетс (2004). «Электрические синапсы в мозгу млекопитающих». Анну. Преподобный Neurosci. 27: 393–418. Дои:10.1146 / annurev.neuro.26.041002.131128. PMID 15217338.
- ^ Pfenniger A, Wohlwend A, Kwak BR (январь 2011 г.). «Мутации в генах коннексинов и болезни». Евро. J. Clin. Вкладывать деньги. 41 (1): 103–16. Дои:10.1111 / j.1365-2362.2010.02378.x. ISSN 1365-2362. PMID 20840374.
- ^ Молика Ф., Минс М.Дж., Морел С., Квак Б.Р. (сентябрь 2014 г.). «Мутации в генах сердечно-сосудистых коннексинов». Биол. Клетка. 106 (9): 269–93. Дои:10.1111 / boc.201400038. PMID 24966059. S2CID 10070999.
внешняя ссылка
 СМИ, связанные с коннексины в Wikimedia Commons
СМИ, связанные с коннексины в Wikimedia Commons- Коннексины в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
По состоянию на это редактирование, в этой статье используется контент из «1.A.24 Семейство коннексинов (коннексинов), образующих щелевые соединения», который лицензирован таким образом, чтобы разрешить повторное использование в соответствии с Creative Commons Attribution-ShareAlike 3.0 Непортированная лицензия, но не под GFDL. Все соответствующие условия должны быть соблюдены.
| Авторитетный контроль |
|---|