ДНК-полимераза бета - DNA polymerase beta





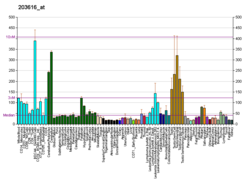
| Регуляторный элемент стволовой петли II в POLB | |
|---|---|
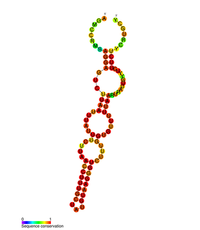 Прогнозируемая вторичная структура регуляторного элемента стволовой петли II (M2) в POLB | |
| Идентификаторы | |
| Символ | ПОЛБ |
| Рфам | RF01455 |
| NCBI Gene | 5423 |
| HGNC | 9174 |
| OMIM | 174760 |
| RefSeq | NM_002690 |
| Прочие данные | |
| РНК тип | Снг |
| Домен (ы) | Млекопитающие |
| Locus | Chr. 8 p11.2 |
| PDB структуры | PDBe |
ДНК-полимераза бета, также известен как ПОЛБ, является фермент присутствует в эукариоты. У человека он кодируется ПОЛБ ген.[5]
Функция
В эукариотический клетки, ДНК-полимераза бета (POLB) выполняет основная эксцизионная пластика (BER) требуется для ДНК поддержание, репликация, рекомбинация и лекарственная устойчивость.[5]
В митохондриальная ДНК клеток млекопитающих постоянно подвергается атакам со стороны кислородные радикалы выпущен во время Производство АТФ. Митохондрии клеток млекопитающих содержат эффективный основная эксцизионная пластика система, использующая POLB, которая удаляет некоторые частые окислительные повреждения ДНК.[6] Таким образом, POLB играет ключевую роль в поддержании стабильности митохондриальной геном.[6]
Анализ верности Репликация ДНК полимеразой бета в нейроны от молодых и очень старых мышей показали, что старение не оказывает значительного влияния на точность синтеза ДНК полимеразой бета.[7] Это открытие было сочтено доказательством против ошибки теории старения как катастрофы.[7][8]
Регулирование выражения
ДНК-полимераза бета поддерживает целостность генома, участвуя в основная эксцизионная пластика. Сверхэкспрессия POLB мРНК коррелирует с рядом типов рака, в то время как недостаток POLB приводит к гиперчувствительности к алкилирующие агенты, индуцированный апоптоз, и хромосомный разрыв. Следовательно, важно, чтобы экспрессия POLB строго регулировалась.[9][10][11][12]
Ген POLB активируется CREB1 фактор транскрипции привязан к элемент ответа cAMP (CRE) присутствует в промоторе гена POLB в ответ на воздействие алкилирующих агентов.[13][14] Экспрессия гена POLB также регулируется на посттранскрипционном уровне как 3 ’UTR мРНК POLB содержат три стебель-петля структуры, влияющие на экспрессию генов.[15] Эти трехстворчатые петлевые структуры известны как M1, M2 и M3, где M2 и M3 играют ключевую роль в регуляции генов. M3 способствует экспрессии генов, поскольку содержит полиаденилирование сигнал, за которым следует сайт расщепления и полиаденилирования, тем самым способствуя пре-мРНК обработка. Было показано, что M2 эволюционно консервативен, и с помощью мутагенеза было показано, что эта структура петли стебля действует как дестабилизирующий элемент РНК.
В дополнение к этим цис-регуляторные элементы присутствует в 3’UTR a трансакционный белок HAX1 считается, что способствует регуляции экспрессии генов. Трехгибридные анализы дрожжей показали, что этот белок связывается со стволовыми петлями в пределах 3’UTR мРНК POLB, однако точный механизм регуляции экспрессии генов этим белком еще предстоит определить.
Взаимодействия
Было показано, что бета-ДНК-полимераза взаимодействовать с участием ПНКП[16] и XRCC1.[17][18][19][20]
Смотрите также
Модельные организмы
Модельные организмы были использованы при изучении функции POLB. Условный нокаутирующая мышь линия называется Polbtm1a (КОМП) Wtsi был создан на Wellcome Trust Sanger Institute.[21] Самцы и самки животных прошли стандартизованный фенотипический скрининг[22] для определения последствий удаления.[23][24][25][26] Проведены дополнительные проверки: - Углубленное иммунологическое фенотипирование[27]
| Характеристика | Фенотип |
|---|---|
| Все данные доступны на сайте.[22][27]{ | |
| Лейкоциты периферической крови 6 недель | Нормальный |
| Гематология 6 недель | Нормальный |
| Гомозиготная жизнеспособность на P14 | Аномальный |
| Рецессивный смертельное исследование | Аномальный |
| Масса тела | Нормальный |
| Неврологический осмотр | Нормальный |
| Сила захвата | Нормальный |
| Дисморфология | Нормальный |
| Косвенная калориметрия | Нормальный |
| Тест толерантности к глюкозе | Нормальный |
| Слуховой ответ ствола мозга | Нормальный |
| DEXA | Нормальный |
| Морфология глаза | Нормальный |
| Клиническая химия | Нормальный |
| Гематология 16 недель | Нормальный |
| Лейкоциты периферической крови 16 недель | Нормальный |
| Вес сердца | Нормальный |
| Сальмонелла инфекция | Нормальный |
| Функция цитотоксических Т-клеток | Нормальный |
| Иммунофенотипирование селезенки | Нормальный |
| Иммунофенотипирование мезентериальных лимфатических узлов | Нормальный |
| Иммунофенотипирование костного мозга | Нормальный |
| Состав эпидермального иммунитета | Нормальный |
| Тричурис Вызов | Нормальный |
использованная литература
- ^ а б c ГРЧ38: Ансамбль выпуск 89: ENSG00000070501 - Ансамбль, Май 2017
- ^ а б c GRCm38: выпуск Ensembl 89: ENSMUSG00000031536 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ "Ссылка на Mouse PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ а б «Ген NCBI: ДНК-полимераза бета».
- ^ а б Прасад Р., Чаглаян М., Дай Д.П., Надалутти К.А., Чжао М.Л., Гассман Н.Р., Яношази А.К., Стефаник Д.Ф., Хортон Дж. К., Красич Р., Лонгли М.Дж., Коупленд В.К., Гриффит Д.Д., Уилсон С.Х. (декабрь 2017 г.). «ДНК-полимераза β: недостающее звено в механизме эксцизионной репарации оснований в митохондриях млекопитающих». Ремонт ДНК (Amst.). 60: 77–88. Дои:10.1016 / j.dnarep.2017.10.011. ЧВК 5919216. PMID 29100041.
- ^ а б Субба Рао К., Мартин Г.М., Леб Л.А. (октябрь 1985 г.). «Верность ДНК-полимеразы-бета в нейронах молодых и очень старых мышей». J. Neurochem. 45 (4): 1273–8. Дои:10.1111 / j.1471-4159.1985.tb05553.x. PMID 3161998. S2CID 84448241.
- ^ Оргель Л.Е. (июнь 1973 г.). «Старение клонов клеток млекопитающих». Природа. 243 (5408): 441–5. Дои:10.1038 / 243441a0. PMID 4591306. S2CID 4153800.
- ^ Canitrot Y, Cazaux C, Fréchet M, Bouayadi K, Lesca C, Salles B, Hoffmann JS (октябрь 1998 г.). «Сверхэкспрессия ДНК-полимеразы бета в клетке приводит к мутаторному фенотипу и снижению чувствительности к противоопухолевым препаратам». Труды Национальной академии наук Соединенных Штатов Америки. 95 (21): 12586–90. Дои:10.1073 / пнас.95.21.12586. ЧВК 22874. PMID 9770529.
- ^ Bergoglio V, Pillaire MJ, Lacroix-Triki M, Raynaud-Messina B, Canitrot Y, Bieth A, Garès M, Wright M, Delsol G, Loeb LA, Cazaux C, Hoffmann JS (июнь 2002 г.). «Дерегулированная ДНК-полимераза бета вызывает хромосомную нестабильность и онкогенез». Исследования рака. 62 (12): 3511–4. PMID 12067997.
- ^ Bergoglio V, Canitrot Y, Hogarth L, Minto L, Howell SB, Cazaux C, Hoffmann JS (сентябрь 2001 г.). «Повышенная экспрессия и активность ДНК-полимеразы бета в опухолевых клетках яичников человека: влияние на чувствительность к противоопухолевым агентам». Онкоген. 20 (43): 6181–7. Дои:10.1038 / sj.onc.1204743. PMID 11593426.
- ^ Шривастава Д.К., Хусейн И., Артеага С.Л., Уилсон С.Х. (июнь 1999 г.). «Различия в экспрессии ДНК-полимеразы бета в отдельных опухолях человека и клеточных линиях». Канцерогенез. 20 (6): 1049–54. Дои:10.1093 / carcin / 20.6.1049. PMID 10357787.
- ^ Хе Ф, Ян XP, Шривастава Д.К., Уилсон С.Х. (январь 2003 г.). «Экспрессия гена ДНК-полимеразы бета: активатор промотора CREB-1 активируется в клетках яичника китайского хомячка под действием стресса, вызванного ДНК-алкилирующим агентом». Биологическая химия. 384 (1): 19–23. Дои:10.1515 / BC.2003.003. PMID 12674496. S2CID 33798724.
- ^ Нараян С., Хе Ф, Уилсон С.Х. (август 1996 г.). «Активация промотора бета-ДНК-полимеразы человека ДНК-алкилирующим агентом посредством индуцированного фосфорилирования белка-1, связывающего элемент ответа цАМФ». Журнал биологической химии. 271 (31): 18508–13. Дои:10.1074 / jbc.271.31.18508. PMID 8702497.
- ^ Сарновска Э., Гжибовска Э.А., Собчак К., Конопинский Р., Вильчинска А., Шварц М., Сарновски Т.Дж., Кшизосяк В.Дж., Седлецкий Я.А. (2007). «Структура шпильки в 3'UTR мРНК ДНК-полимеразы бета действует как посттранскрипционный регуляторный элемент и взаимодействует с Hax-1». Исследования нуклеиновых кислот. 35 (16): 5499–510. Дои:10.1093 / нар / гкм502. ЧВК 2018635. PMID 17704138.
- ^ Уайтхаус CJ, Тейлор RM, Thistlethwaite A, Zhang H, Karimi-Busheri F, Lasko DD, Weinfeld M, Caldecott KW (январь 2001 г.). «XRCC1 стимулирует активность полинуклеотидкиназы человека на поврежденных концах ДНК и ускоряет восстановление однонитевых разрывов ДНК». Ячейка. 104 (1): 107–17. Дои:10.1016 / S0092-8674 (01) 00195-7. PMID 11163244. S2CID 1487128.
- ^ Ван Л., Бхаттачарья Н., Челси Д.М., Эскобар П.Ф., Банерджи С. (ноябрь 2004 г.). «Новый ядерный белок MGC5306 взаимодействует с ДНК-полимеразой бета и потенциально играет роль в клеточном фенотипе». Исследования рака. 64 (21): 7673–7. Дои:10.1158 / 0008-5472.CAN-04-2801. PMID 15520167.
- ^ Fan J, Otterlei M, Wong HK, Tomkinson AE, Wilson DM (2004). «XRCC1 совместно локализуется и физически взаимодействует с PCNA». Исследования нуклеиновых кислот. 32 (7): 2193–201. Дои:10.1093 / нар / гх556. ЧВК 407833. PMID 15107487.
- ^ Кубота Ю., Нэш Р.А., Клунгланд А., Шер П., Барнс Д.Э., Линдал Т. (декабрь 1996 г.). «Восстановление эксцизионной репарации оснований ДНК очищенными белками человека: взаимодействие между ДНК-полимеразой бета и белком XRCC1». Журнал EMBO. 15 (23): 6662–70. Дои:10.1002 / j.1460-2075.1996.tb01056.x. ЧВК 452490. PMID 8978692.
- ^ Бхаттачарья Н., Банерджи С. (июль 2001 г.). «Новая роль XRCC1 в функциях бета-варианта ДНК-полимеразы». Биохимия. 40 (30): 9005–13. Дои:10.1021 / bi0028789. PMID 11467963.
- ^ Гердин А.К. (2010). «Программа генетики Sanger Mouse: характеристика мышей с высокой пропускной способностью». Acta Ophthalmologica. 88: 925–7. Дои:10.1111 / j.1755-3768.2010.4142.x. S2CID 85911512.
- ^ а б «Международный консорциум по фенотипированию мышей».
- ^ Скарнес В.К., Розен Б., Вест А.П., Кутсуракис М., Бушелл В., Айер В., Мухика А.О., Томас М., Харроу Дж., Кокс Т., Джексон Д., Северин Дж., Биггс П., Фу Дж., Нефедов М., де Йонг П.Дж., Стюарт AF, Брэдли А. (июнь 2011 г.). «Ресурс условного нокаута для полногеномного исследования функции генов мыши». Природа. 474 (7351): 337–42. Дои:10.1038 / природа10163. ЧВК 3572410. PMID 21677750.
- ^ Долгин Э (июнь 2011 г.). "Библиотека мыши настроена на нокаут". Природа. 474 (7351): 262–3. Дои:10.1038 / 474262a. PMID 21677718.
- ^ Коллинз Ф.С., Россант Дж., Вурст В. (январь 2007 г.). «Мышь по всем причинам». Ячейка. 128 (1): 9–13. Дои:10.1016 / j.cell.2006.12.018. PMID 17218247. S2CID 18872015.
- ^ Уайт Дж. К., Гердин А. К., Карп Н. А., Райдер Э., Бульян М., Басселл Дж. Н., Солсбери Дж., Клэр С., Ингем Нью-Джерси, Подрини С., Хоутон Р., Эстабель Дж., Боттомли Дж. , Logan DW, Macarthur DG, Flint J, Mahajan VB, Tsang SH, Smyth I, Watt FM, Skarnes WC, Dougan G, Adams DJ, Ramirez-Solis R, Bradley A, Steel KP (июль 2013 г.). «Полногеномное поколение и систематическое фенотипирование мышей с нокаутом открывает новые роли для многих генов». Ячейка. 154 (2): 452–64. Дои:10.1016 / j.cell.2013.06.022. ЧВК 3717207. PMID 23870131.
- ^ а б «Консорциум иммунофенотипирования инфекций и иммунитета (3i)».
дальнейшее чтение
- Датэ Т., Танихара К., Ямамото С., Номура Н., Мацукаге А. (сентябрь 1992 г.). «Две области в мРНК бета-полимеразы ДНК человека подавляют трансляцию в Escherichia coli». Исследования нуклеиновых кислот. 20 (18): 4859–64. Дои:10.1093 / nar / 20.18.4859. ЧВК 334243. PMID 1408801.
- Ван Л., Патель У., Гош Л., Банерджи С. (сентябрь 1992 г.). «Мутации бета-ДНК-полимеразы при колоректальном раке человека». Исследования рака. 52 (17): 4824–7. PMID 1511447.
- Токуи Т., Инагаки М., Нисидзава К., Ятани Р., Кусагава М., Аджиро К., Нисимото И., Дате Т., Мацукаге А. (июнь 1991 г.). «Инактивация ДНК-полимеразы бета путем фосфорилирования in vitro протеинкиназой C». Журнал биологической химии. 266 (17): 10820–4. PMID 2040602.
- SenGupta DN, Zmudzka BZ, Kumar P, Cobianchi F, Skowronski J, Wilson SH (апрель 1986). «Последовательность мРНК бета-ДНК-полимеразы человека, полученная путем клонирования кДНК». Сообщения о биохимических и биофизических исследованиях. 136 (1): 341–7. Дои:10.1016 / 0006-291X (86) 90916-2. PMID 2423078.
- Zmudzka BZ, Fornace A, Collins J, Wilson SH (октябрь 1988 г.). «Характеристика мРНК ДНК-полимеразы бета: клеточный цикл и ростовая реакция в культивируемых клетках человека». Исследования нуклеиновых кислот. 16 (20): 9587–96. Дои:10.1093 / nar / 16.20.9587. ЧВК 338765. PMID 2460824.
- Уайден С.Г., Кедар П., Уилсон С.Х. (ноябрь 1988 г.). «Ген бета-полимеразы человека. Структура 5'-фланкирующей области и активный промотор». Журнал биологической химии. 263 (32): 16992–8. PMID 3182828.
- Abbotts J, SenGupta DN, Zmudzka B, Widen SG, Notario V, Wilson SH (февраль 1988 г.). «Экспрессия бета-полимеразы ДНК человека в Escherichia coli и характеристика рекомбинантного фермента». Биохимия. 27 (3): 901–9. Дои:10.1021 / bi00403a010. PMID 3284575.
- Добаши Ю., Кубота Ю., Шуин Т., Торигоэ С., Яо М., Хосака М. (апрель 1995 г.). «Полиморфизмы в гене бета-полимеразы ДНК человека». Генетика человека. 95 (4): 389–90. Дои:10.1007 / bf00208961. PMID 7705833. S2CID 22658070.
- Chyan YJ, Ackerman S, Shepherd NS, McBride OW, Widen SG, Wilson SH, Wood TG (июль 1994 г.). «Структура гена ДНК-полимеразы бета человека. Доказательства альтернативного сплайсинга в экспрессии генов». Исследования нуклеиновых кислот. 22 (14): 2719–25. Дои:10.1093 / nar / 22.14.2719. ЧВК 308239. PMID 7914364.
- Маруяма К., Сугано С. (январь 1994 г.). «Олиго-кэппинг: простой метод замены кэп-структуры эукариотических мРНК олигорибонуклеотидами». Ген. 138 (1–2): 171–4. Дои:10.1016/0378-1119(94)90802-8. PMID 8125298.
- Чанг М., Бурмер Г.К., Суизи Дж., Лоеб Л.А., Эдельхофф С., Disteche CM, Ю. CE, Андерсон Л., Осима Дж., Накура Дж. (Май 1994 г.). «Доказательства против ДНК-полимеразы бета как гена-кандидата на синдром Вернера». Генетика человека. 93 (5): 507–12. Дои:10.1007 / bf00202813. PMID 8168825. S2CID 34824853.
- Чиан Ю.Дж., Штраус ПР, Вуд Т.Г., Уилсон С.Х. (август 1996 г.). «Идентификация новых изоформ мРНК для бета-полимеразы ДНК человека». ДНК и клеточная биология. 15 (8): 653–9. Дои:10.1089 / dna.1996.15.653. PMID 8769567.
- Пеллетье Х, Савая М.Р., Вольфле В., Уилсон С.Х., Краут Дж. (Октябрь 1996 г.). «Кристаллические структуры бета-полимеразы ДНК человека в комплексе с ДНК: значение для каталитического механизма, процессивности и точности». Биохимия. 35 (39): 12742–61. Дои:10.1021 / bi952955d. PMID 8841118.
- Пеллетье Х, Савая М.Р., Вольфле В., Уилсон С.Х., Краут Дж. (Октябрь 1996 г.). «Структурная основа мутагенности иона металлов и нуклеотидной селективности в человеческой ДНК-полимеразе бета». Биохимия. 35 (39): 12762–77. Дои:10.1021 / bi9529566. PMID 8841119.
- Пеллетье Х., Савая М.Р. (октябрь 1996 г.). «Характеристика мотивов спирали-шпильки-спирали, связывающих ионы металла, в человеческой ДНК-полимеразе бета с помощью рентгеноструктурного анализа». Биохимия. 35 (39): 12778–87. Дои:10.1021 / bi960790i. PMID 8841120.
- Кубота Ю., Нэш Р.А., Клунгланд А., Шер П., Барнс Д.Э., Линдал Т. (декабрь 1996 г.). «Восстановление эксцизионной репарации оснований ДНК очищенными белками человека: взаимодействие между ДНК-полимеразой бета и белком XRCC1». Журнал EMBO. 15 (23): 6662–70. Дои:10.1002 / j.1460-2075.1996.tb01056.x. ЧВК 452490. PMID 8978692.
- Беннетт Р.А., Уилсон Д.М., Вонг Д., Демпл Б. (июль 1997 г.). «Взаимодействие апуриновой эндонуклеазы человека и ДНК-полимеразы бета в пути эксцизионной репарации оснований». Труды Национальной академии наук Соединенных Штатов Америки. 94 (14): 7166–9. Дои:10.1073 / пнас.94.14.7166. ЧВК 23779. PMID 9207062.
- Sawaya MR, Prasad R, Wilson SH, Kraut J, Pelletier H (сентябрь 1997 г.). «Кристаллические структуры бета-полимеразы ДНК человека в комплексе с ДНК с разрывом и разрывом: свидетельство механизма индуцированной подгонки». Биохимия. 36 (37): 11205–15. Дои:10.1021 / bi9703812. PMID 9287163.
- Бхаттачарья Н., Банерджи С. (сентябрь 1997 г.). «Вариант бета-полимеразы ДНК действует как доминантно-негативный мутант». Труды Национальной академии наук Соединенных Штатов Америки. 94 (19): 10324–9. Дои:10.1073 / пнас.94.19.10324. ЧВК 23361. PMID 9294209.
- Судзуки Ю., Ёситомо-Накагава К., Маруяма К., Суяма А., Сугано С. (октябрь 1997 г.). «Создание и характеристика полноразмерной библиотеки кДНК, обогащенной по 5'-концу». Ген. 200 (1–2): 149–56. Дои:10.1016 / S0378-1119 (97) 00411-3. PMID 9373149.
- Тан XH, Чжао М., Пан К.Ф., Донг Й, Донг Б., Фэн Г.Дж., Цзя Г., Лу Й.Й. (март 2005 г.). «Частая мутация, связанная со сверхэкспрессией ДНК-полимеразы бета в первичных опухолях и предраковых поражениях желудка человека». Письма о раке. 220 (1): 101–14. Дои:10.1016 / j.canlet.2004.07.049. PMID 15737693.
внешние ссылки
Галерея PDB | |
|---|---|
|