Нейронная цепь - Neural circuit - Wikipedia

А нейронная цепь это население нейроны связаны между собой синапсы для выполнения определенной функции при активации.[1] Нейронные цепи соединяются друг с другом, образуя крупномасштабные сети мозга.[2] Биологические нейронные сети вдохновили дизайн искусственные нейронные сети, но искусственные нейронные сети обычно не являются точными копиями своих биологических аналогов.
Раннее обучение
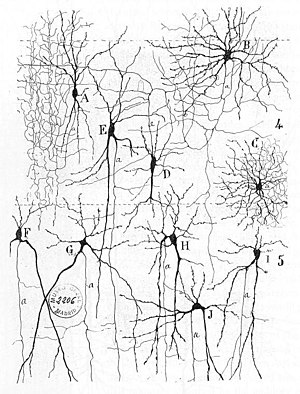
Раннее лечение невральной сети можно найти в Герберт Спенсер с Принципы психологии, 3-е издание (1872 г.), Теодор Мейнерт с Психиатрия (1884), Уильям Джеймс ' Принципы Психология (1890), и Зигмунд Фрейд "Проект научной психологии" (составлен в 1895 г.).[3] Первое правило обучения нейронов было описано Hebb в 1949 г. Хеббийская теория. Таким образом, хеббовское сочетание пресинаптической и постсинаптической активности может существенно изменить динамические характеристики синаптической связи и, следовательно, либо облегчить, либо подавить передача сигнала. В 1959 г. нейробиологи, Уоррен Стерджис Маккалох и Уолтер Питтс опубликовал первые работы по обработке нейронных сетей.[4] Теоретически они показали, что сети искусственных нейронов могут воплощать в жизнь логичный, арифметика, и символический функции. Упрощенный модели биологических нейронов были созданы, теперь обычно называются перцептроны или же искусственные нейроны. Эти простые модели составили нейронное суммирование (т.е. потенциалы на постсинаптической мембране будут суммироваться в Тело клетки ). Более поздние модели также обеспечивали возбуждающую и тормозящую синаптическую передачу.
Связи между нейронами
Связи между нейронами головного мозга намного сложнее, чем у нейронов. искусственные нейроны используется в коннекционист нейронные вычислительные модели искусственные нейронные сети. Основные виды связей между нейронами: синапсы: обе химический и электрические синапсы.
Создание синапсов позволяет соединять нейроны в миллионы перекрывающихся и взаимосвязанных нейронных цепей. Пресинаптические белки, называемые нейрексины занимают центральное место в этом процессе.[5]
Один принцип, по которому работают нейроны: нейронное суммирование – потенциалы на постсинаптическая мембрана суммируется в теле клетки. Если деполяризация нейрона на аксональный бугорок превышает пороговое значение, возникает потенциал действия, который движется вниз аксон к конечным окончаниям для передачи сигнала другим нейронам. Возбуждающая и тормозная синаптическая передача осуществляется главным образом посредством возбуждающие постсинаптические потенциалы (EPSP), и тормозящие постсинаптические потенциалы (IPSP).
На электрофизиологический уровне, существуют различные явления, которые изменяют характеристики отклика отдельных синапсов (называемых синаптическая пластичность ) и отдельные нейроны (собственная пластичность ). Их часто делят на кратковременную пластичность и долговременную пластичность. Долгосрочная синаптическая пластичность часто считается наиболее вероятной. объем памяти субстрат. Обычно термин "нейропластичность «относится к изменениям в мозгу, вызванным деятельностью или опытом.
Связи отображают временные и пространственные характеристики. Временные характеристики относятся к непрерывно изменяющейся зависящей от активности эффективности синаптической передачи, называемой пластичность, зависящая от времени всплеска. В нескольких исследованиях было замечено, что синаптическая эффективность этой передачи может кратковременно увеличиваться (называемая содействие ) или уменьшить (депрессия ) по активности пресинаптического нейрона. Индукция долгосрочных изменений синаптической эффективности за счет долгосрочное потенцирование (LTP) или депрессия (LTD), сильно зависит от относительного времени начала возбуждающий постсинаптический потенциал и постсинаптический потенциал действия. LTP индуцируется серией потенциалов действия, которые вызывают различные биохимические реакции. В конечном итоге реакции вызывают экспрессию новых рецепторов на клеточных мембранах постсинаптических нейронов или повышают эффективность существующих рецепторов за счет фосфорилирование.
Потенциалы обратного распространения не могут возникать, потому что после того, как потенциал действия проходит по заданному сегменту аксона, м ворота на напряжение-управляемые натриевые каналы закрыть, таким образом блокируя любое временное открытие ворота h от изменения внутриклеточного иона натрия (Na+) концентрации и предотвращение генерации потенциала действия обратно к телу клетки. Однако в некоторых камерах нейронное обратное распространение действительно происходит через дендритное ветвление и может иметь важные последствия для синаптической пластичности и вычислений.
Нейрон в головном мозге требует единственного сигнала для нервномышечное соединение стимулировать сокращение постсинаптических мышечных клеток. Однако в спинном мозге не менее 75 афферентный нейроны должны производить возбуждение. Эта картина еще больше усложняется из-за изменения постоянной времени между нейронами, поскольку некоторые клетки могут испытывать свои ВПСП в течение более длительного периода времени, чем другие.
Пока в синапсах в развивающийся мозг синаптическая депрессия особенно широко наблюдалась; предполагалось, что она переходит в облегчение во взрослом мозге.
Схема

Примером нейронной схемы является трисинаптический контур в гиппокамп. Другой - это Трасса Папеза связь гипоталамус к лимбическая доля. В системе есть несколько нейронных цепей. кортико-базальные ганглии-таламо-кортикальная петля. Эти цепи несут информацию между корой головного мозга, базальный ганглий, таламус и обратно в кору. Самая крупная структура базальных ганглиев - полосатое тело, рассматривается как имеющая собственную внутреннюю микросхему.[6]
Нейронные цепи в спинной мозг называется генераторы центральных паттернов отвечают за управление моторными инструкциями, участвующими в ритмическом поведении. Ритмичное поведение включает ходьбу, мочеиспускание, и эякуляция. Генераторы центральных паттернов состоят из различных групп спинномозговые интернейроны.[7]
Существует четыре основных типа нейронных цепей, которые отвечают за широкий спектр нейронных функций. Эти схемы представляют собой расходящаяся цепь, а сходящаяся схема, а реверберационный контур, а параллельная цепь постразряда.[8]
В расходящейся цепи один нейрон соединяется с несколькими постсинаптическими клетками. Каждый из них может быть синапсом со многими другими, что позволяет одному нейрону стимулировать до тысяч клеток. Это проиллюстрировано тем, как тысячи мышечных волокон могут быть стимулированы первоначальным входом от одного двигательный нейрон.[8]
В конвергентной схеме входы из многих источников объединяются в один выход, влияя только на один нейрон или пул нейронов. Этот тип схемы проиллюстрирован на дыхательный центр из мозговой ствол, который реагирует на ряд входных сигналов из разных источников, выдавая соответствующий паттерн дыхания.[8]
Схема реверберации дает повторяющийся выходной сигнал. В процедуре передачи сигналов от одного нейрона к другому в линейной последовательности один из нейронов может послать сигнал обратно инициирующему нейрону. Каждый раз, когда срабатывает первый нейрон, другой нейрон, расположенный ниже по последовательности, снова срабатывает, отправляя его обратно источнику. . Это рестимулирует первый нейрон, а также позволяет продолжить путь передачи до его выхода. Результирующий повторяющийся паттерн - это результат, который прекращается только в том случае, если один или несколько синапсов выходят из строя, или если тормозящий поток из другого источника вызывает его остановку. Этот тип реверберирующего контура находится в дыхательном центре, который посылает сигналы в дыхательные мышцы, вызывая вдыхание. Когда цепь прерывается тормозным сигналом, мышцы расслабляются, вызывая выдох. Этот тип схемы может играть роль в Эпилептические припадки.[8]
В параллельной цепи после разряда нейрон вводит сигналы в несколько цепочек нейронов. Каждая цепочка состоит из разного количества нейронов, но их сигналы сходятся на одном выходном нейроне. Каждый синапс в схеме задерживает сигнал примерно на 0,5 мс, так что чем больше синапсов будет, тем дольше будет задержка выходного нейрона. После того, как вход остановлен, выход продолжит работу в течение некоторого времени. Этот тип схемы не имеет петли обратной связи, как реверберирующая схема. Продолжение возбуждения после прекращения действия стимула называется после выписки. Этот тип схемы находится в рефлекторные дуги определенных рефлексы.[8]
Методы исследования
Разные нейровизуализация были разработаны методы исследования активности нейронных цепей и сетей. Использование «сканеров мозга» или функциональной нейровизуализации для исследования структуры или функций мозга является обычным явлением либо просто как способ более точной оценки травмы мозга с помощью изображений с высоким разрешением, либо путем изучения относительной активации различных областей мозга. Такие технологии могут включать функциональная магнитно-резонансная томография (фМРТ), позитронно-эмиссионная томография мозга (ПЭТ головного мозга) и компьютерная аксиальная томография (CAT) сканирование. Функциональная нейровизуализация использует определенные технологии визуализации мозга для получения снимков мозга, обычно когда человек выполняет конкретную задачу, в попытке понять, как активация определенных областей мозга связана с задачей. При функциональной нейровизуализации, особенно фМРТ, которая измеряет гемодинамическая активность (с помощью BOLD-контрастное изображение ), который тесно связан с нейронной активностью, ПЭТ и электроэнцефалография (ЭЭГ).
Коннекционист модели служат платформой для тестирования различных гипотез представления, обработки информации и передачи сигналов. Исследования поражения на таких моделях, например искусственные нейронные сети, где части узлов преднамеренно разрушаются, чтобы увидеть, как работает сеть, также может дать важную информацию о работе нескольких сборок ячеек. Аналогичным образом моделирование дисфункциональных нейротрансмиттеров в неврологических условиях (например, дофамин в базальных ганглиях Болезнь Паркинсона пациенты) могут дать представление о механизмах, лежащих в основе моделей когнитивных нарушений, наблюдаемых в конкретной группе пациентов. Прогнозы на основе этих моделей могут быть проверены на пациентах или с помощью фармакологических манипуляций, а эти исследования, в свою очередь, могут использоваться для информирования моделей, что делает процесс повторяющимся.
Современный баланс между коннекционистским подходом и одноклеточным подходом в нейробиология была достигнута в результате длительного обсуждения. В 1972 году Барлоу объявил революция одиночного нейрона: «наши восприятия вызваны активностью довольно небольшого числа нейронов, выбранных из очень большой популяции преимущественно молчащих клеток».[9] Этот подход был стимулирован идеей клетка бабушки выдвинут двумя годами ранее. Барлоу сформулировал «пять догм» нейронной доктрины. Недавние исследования 'клетка бабушки 'и явления разреженного кодирования развивают и видоизменяют эти идеи.[10] В экспериментах с одной клеткой использовались внутричерепные электроды в медиальной височной доле (гиппокамп и окружающая кора). Современное развитие концентрация меры теории (теоремы стохастического разделения) с приложениями к искусственные нейронные сети дают математическое обоснование неожиданной эффективности малых нейронных ансамблей в многомерном мозге.[11]
Клиническое значение
Иногда нейронные цепи могут становиться патологическими и вызывать такие проблемы, как болезнь Паркинсона когда базальный ганглий вовлечены.[12] Проблемы в Трасса Папеза может также вызвать ряд нейродегенеративные расстройства включая болезнь Паркинсона.
Смотрите также
- Обратная связь
- Список регионов человеческого мозга
- Сетевая наука
- Нейронное кодирование
- Нейронная инженерия
- Нейронные колебания
- Сети с импульсной связью
- Системная неврология
Рекомендации
- ^ Purves, Дейл (2011). Неврология (5-е изд.). Сандерленд, Массачусетс: Sinauer. п. 507. ISBN 9780878936953.
- ^ "Нейронные цепи | Центр передового опыта в области интегративной функции мозга". Центр передового опыта в области интегративной функции мозга. 13 июня 2016 г.. Получено 4 июн 2018.
- ^ Майкл С. К. Томас; Джеймс Л. Макклелланд. «Коннекционистские модели познания» (PDF). Стэндфордский Университет. Архивировано из оригинал (PDF) на 2015-09-06. Получено 2015-08-31.
- ^ J. Y. Lettvin; Х. Р. Матурана; У. С. Маккалох; В. Х. Питтс (1959), «Что глаз лягушки сообщает мозгу лягушки», Proc. Inst. Radio Engr. (47), стр. 1940–1951.
- ^ Зюдхоф, ТЦ (2 ноября 2017 г.). «Синаптические комплексы нейрексинов: молекулярный код логики нейронных цепей». Клетка. 171 (4): 745–769. Дои:10.1016 / j.cell.2017.10.024. ЧВК 5694349. PMID 29100073.
- ^ Стокко, Андреа; Лебьер, Кристиан; Андерсон, Джон Р. (2010). «Условная маршрутизация информации в кору: модель роли базальных ганглиев в когнитивной координации». Психологический обзор. 117 (2): 541–74. Дои:10.1037 / a0019077. ЧВК 3064519. PMID 20438237.
- ^ Гертин, PA (2012). «Центральный генератор паттернов для передвижения: анатомические, физиологические и патофизиологические соображения». Границы неврологии. 3: 183. Дои:10.3389 / fneur.2012.00183. ЧВК 3567435. PMID 23403923.
- ^ а б c d е Саладин, К. Анатомия человека (3-е изд.). Макгроу-Хилл. п. 364. ISBN 9780071222075.
- ^ Барлоу, HB (1 декабря 1972 г.). «Единичные единицы и ощущение: нейронная доктрина для психологии восприятия?». Восприятие. 1 (4): 371–394. Дои:10.1068 / p010371. PMID 4377168.
- ^ Quian Quiroga, R; Редди, L; Крейман, G; Koch, C; Fried, I (23 июня 2005 г.). «Инвариантное визуальное представление отдельными нейронами в человеческом мозгу». Природа. 435 (7045): 1102–1107. Дои:10.1038 / природа03687. PMID 15973409.
- ^ Горбань, Александр Н .; Макаров, Валерий А .; Тюкин, Иван Юрьевич (июль 2019). «Неоправданная эффективность малых нейронных ансамблей в многомерном мозге». Обзоры физики жизни. 29: 55–88. arXiv:1809.07656. Дои:10.1016 / j.plrev.2018.09.005. PMID 30366739.
- ^ Французский, IT; Мутусами, KA (2018). "Обзор ядра педункулопонтина при болезни Паркинсона". Границы старения нейронауки. 10: 99. Дои:10.3389 / fnagi.2018.00099. ЧВК 5933166. PMID 29755338.
дальнейшее чтение
- Собственная пластичность Роберт Х. Кадмор, Нирадж С. Десаи Scholarpedia 3(2):1363. DOI: 10.4249 / scholarpedia.1363
внешняя ссылка
- Сравнение нейронных сетей в мозге и искусственных нейронных сетей
- Конспект лекций в MIT OpenCourseWare
- Вычисления в мозгу
- Набор инструментов биологической нейронной сети - Бесплатный набор инструментов Matlab для моделирования сетей из нескольких разных типов нейронов
- WormWeb.org: Интерактивная визуализация нейронной сети C. elegans C. elegans, нематода с 302 нейронами, является единственным организмом, для которого была раскрыта вся нейронная сеть. Используйте этот сайт для просмотра сети и поиска путей между любыми двумя нейронами.
- Введение в нейроны и нейронные сети, Неврология онлайн (Электронный учебник неврологии)
- Сети с задержкой импульсов (сети с волновыми помехами)
| Авторитетный контроль |
|---|