Чешуйчатая улитка - Scaly-foot snail
| Чешуйчатая улитка / Морской панголин | |
|---|---|
 | |
| Chrysomallon squamiferum из Longqi. Шкала шкалы 1 см. | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Моллюска |
| Учебный класс: | Брюхоногие |
| Clade: | Неомфалина |
| Семья: | Peltospiridae |
| Род: | Хризомаллон Чен, Линсе, Копли и Роджерс, 2015 г. |
| Разновидность: | С. squamiferum |
| Биномиальное имя | |
| Chrysomallon squamiferum Чен, Линсе, Копли и Роджерс, 2015 г.[2] | |
| Синонимы[2] | |
Crysomallon squamiferum (орт. ошибка) | |

Chrysomallon squamiferum, широко известный как то чешуйчатый брюхоногий моллюск, чешуйчатая улитка, или же морской панголин[3], это разновидность глубоководных гидротермальный источник улитка, а морской брюхоногие моллюски моллюск в семье Peltospiridae.[2] Этот эндемичный брюхоногий моллюск известен только из глубоководных гидротермальных источников в Индийский океан, где он был найден на глубине около 2400–2900 м (1,5–1,8 мили). Chrysomallon squamiferum сильно отличается от других глубоководных брюхоногих моллюсков, даже от близкородственных неомфалин.[4] В 2019 году было объявлено находящихся под угрозой исчезновения в Красном списке МСОП[5], первый вид, который будет указан в качестве такового из-за рисков глубоководная добыча жерловой среды обитания, где также добываются высококачественные металлические руды[6].
Оболочка уникальной конструкции, трехслойная; внешний слой состоит из сульфиды железа средний слой эквивалентен органическому периостракум найдены у других брюхоногих моллюсков, а самый внутренний слой состоит из арагонит. Стопа тоже необычна, по бокам бронирована железо-минерализованными склеритами.
Улитка пищеводная железа дома симбиотический гаммапротеобактерии из которого, кажется, улитка получает питание. Этот вид считается одним из самых своеобразных брюхоногих моллюсков с глубоководными гидротермальными источниками, и это единственное известное сохранившееся животное, которое включает сульфид железа в свой скелет (как в его склериты, так и в свой панцирь в качестве экзоскелета).[2] Его сердце, соответственно, необычно велико для любого животного: оно составляет примерно 4% от объема его тела.[4]
Таксономия
Этот вид был впервые обнаружен в апреле 2001 года, а с 2001 года его называют чешуйчатым брюхоногим.[7] Это упоминалось как Chrysomallon squamiferum с 2003 г., но формально не описывался в смысле Международный кодекс зоологической номенклатуры пока Чен и другие. назвал его в 2015 году.[2][8] Типовые образцы хранятся в Музей естественной истории, Лондон.[2] В то время, когда название еще не было формализовано, неправильный вариант написания был "Crysomallon squamiferum".[2]
С. squamiferum это типовой вид и единственный вид в роде Хризомаллон.[2] Общее название Хризомаллон из Древнегреческий язык, и означает "златовласый", потому что пирит (соединение, находящееся в его оболочке) имеет золотистый цвет.[2] Конкретное имя squamiferum происходит от латинского языка и означает «чешуйчатый» из-за склеритов.[2] Сначала не было известно, к какому семейству принадлежит этот вид.[7] Warén и другие. классифицировал этот вид в семействе Peltospiridae, в пределах Неомфалина в 2003 г.[9] Молекулярный анализ на основе последовательностей цитохром-с оксидаза I (COI) гены подтвердили размещение этого вида в пределах Peltospiridae.[2][10] Морфотипы из двух местонахождений темные; морфотип из третьего местонахождения белый (объяснение местонахождений см. в следующем разделе).[2][11][12] Согласно результатам генетического анализа, эти разноцветные улитки кажутся просто «разновидностями» одного и того же вида.[2]
Распределение

Чешуйчатый брюхоногий моллюск - эндемичный брюхоногий моллюск, известный только из глубоководных гидротермальных источников Индийского океана, глубина которых составляет около 2780 метров (1,73 мили).[2] Вид был обнаружен в 2001 году, обитает на основаниях черные курильщики в Kairei гидротермальный источник поле, 25 ° 19,239' ю.ш. 70 ° 02,429'E / 25.320650 ° ю.ш. 70.040483 ° в., на Центрально-Индийском хребте, к северу от Родригес Triple Point.[7] Впоследствии этот вид был также обнаружен в поле Solitaire, 19 ° 33,413' ю.ш. 65 ° 50,888'E / 19,556883 ° ю.ш. 65,848133 ° в., Центрально-Индийский хребет, в пределах Исключительная экономическая зона из Маврикий[13][14] и Longqi (что означает «флаг дракона» на китайском языке)[15] поле, 37 ° 47,027' ю.ш. 49 ° 38.963'E / 37,783783 ° ю. Ш. 49,649383 ° в., Юго-Западный Индийский хребет.[16][17] Месторождение Лунци было обозначено как типовая; все типы материалов происходят из этого вентиляционного поля.[2] Расстояние между Кайреем и Пасьянсом составляет около 700 км (430 миль). Расстояние между Solitaire и Longqi составляет около 2500 км (1600 миль).[2] Эти три участка принадлежат Индийскому океану. биогеографическая провинция гидротермальных жерловых систем Sensu Роджерс и другие. (2012).[18] Расстояние между площадками велико, но общая площадь распространения очень мала, менее 0,02 квадратных километра (0,0077 квадратных миль).[19]
Известно, что улитки Peltospiridae обитают в основном на жерловых полях Восточной части Тихого океана. Накамура и другие. предположили, что появление чешуйчатых брюхоногих моллюсков в Индийском океане предполагает связь фаун гидротермальных источников между этими двумя областями.[13]В исследовательские экспедиции вошли:
- 2000 г. - экспедиция Японское агентство морских наук и технологий используя корабль RV Kairei и ROV Кайко открыл жерловое поле Кайрей, но чешуйчатые брюхоногие моллюски в то время не были обнаружены.[20] Это было первое жерловое месторождение, обнаруженное в Индийском океане.[20]
- 2001 г. - экспедиция исследовательского судна США. RV Knorr с ROV Джейсон обнаружил чешуйчатых брюхоногих моллюсков в жерловом поле Кайрей.[7]
- 2007 г. - экспедиция RV Да Ян И Хао открыли жерловое месторождение Лунци.[2]
- 2009 г. - экспедиция RV Йокосука с DSV Синкай 6500 открыл поле Solitaire и отобрал там чешуйчатых брюхоногих моллюсков.[13]
- 2009 г. - экспедиция на Р.В. Да Ян И Хао визуально наблюдаемые чешуйчатые брюхоногие моллюски на жерловом поле Лунци[2][16]
- 2011 г. - экспедиция британского королевского исследовательского корабля. RRS Джеймс Кук с ROV Киль 6000 опробовали жерловое месторождение Longqi.[2][21]
Описание
Склериты

У этого вида стороны лапы улитки чрезвычайно необычны, поскольку они покрыты сотнями минерализованных железом склериты; они состоят из сульфидов железа[9] Greigite и пирит.[22] Каждый склерит имеет мягкий эпителиальный тканевое ядро, а конхиолин Покрытие и самый верхний слой, содержащий пирит и грейгит.[2] До открытия чешуйчатого брюхоногого моллюска считалось, что единственные сохранившиеся моллюски, обладающие чешуевидными структурами, относились к классам Каудофовеата, Соленогастр и Полиплакофора.[17] Склериты не гомологичный брюхоногому моллюску крышка. Склериты брюхоногих моллюсков также не гомологичны склеритам, обнаруженным в хитоны (Полиплакофора).[17] Было высказано предположение, что склериты кембрия халваксииды Такие как Халкирия потенциально могут быть более похожими на склериты этой улитки, чем склериты хитонов или аплакофоров.[17] Еще в 2015 году подробный морфологический анализ для проверки этой гипотезы не проводился.[17]
Склериты Chrysomallon squamiferum в основном белковые (конхиолин - сложный белок); напротив, склериты хитонов в основном известковые.[17] На поперечных срезах склеритов отсутствуют видимые линии роста конхиолина.[17] Никакие другие современные или вымершие брюхоногие моллюски не обладают кожными склеритами.[17] и ни одно другое современное животное не использовало сульфиды железа таким образом, ни в своем скелет,[2] или же экзоскелет.
Размер каждого склерита у взрослых составляет около 1 × 5 мм.[2] У молодых особей чешуйки расположены в несколько рядов, а у взрослых - плотные и асимметричные.[23] Популяция улиток Solitaire имеет белые склериты вместо черных; это связано с недостатком железа в склеритах.[17] Склериты черепчатые (нахлестываются наподобие черепица ).[4] Предполагалось, что целью склеритов является защита или детоксикация.[24] Склериты могут помочь защитить брюхоногих моллюсков от вентиляционной жидкости, так что его бактерии могут жить рядом с источником доноров электронов для хемосинтеза.[4] Или, альтернативно, склериты могут быть результатом отложения токсичных сульфид отходы эндосимбионтов и поэтому представляют собой новое решение для детоксикации.[4] Но истинная функция склеритов пока неизвестна.[13] Склериты популяции Кайрей, имеющие слой сульфида железа, являются ферримагнитный.[2] Склерит морфотипа Solitaire, не минерализованный сульфидом железа, показал большую механическая сила всей конструкции при испытании на трехточечный изгиб (12,06 МПа), чем склерит морфотипа Кайрей (6,54 МПа).[13]
В жизни внешние поверхности склеритов содержат множество разнообразных эпибионты: Эпсилонпротеобактерии и Дельтапротеобактерии.[25] Вероятно, эти бактерии обеспечивают их минерализацию.[25] Гоффреди и другие. (2004) предположили, что улитка выделяет некоторые органические соединения, которые способствуют прикреплению бактерий.[25]
Ракушка

Kairei, Longqi, Solitaire (слева направо)
Панцирь этих видов имеет три завитки.[2] Форма раковины шаровидная, а шпиль сжат.[2] Скульптура раковины состоит из ребер и тонких линий роста.[2] Форма отверстие эллиптическая.[2] В вершина скорлупы хрупкие и у взрослых подвержены коррозии.[2]
Это очень крупные пельтоспириды по сравнению с большинством других видов, которые обычно ниже 15 миллиметров (3⁄5 дюйм) в длине оболочки.[2] Ширина корпуса составляет 9,80–40,02 мм (0,39–1,58 дюйма);[2] максимальная ширина корпуса достигает 45,5 миллиметра (1,79 дюйма).[2] Средняя ширина раковины взрослой улитки 32 мм.[2] Средняя ширина раковины у популяции Solitaire была немного меньше, чем у популяции Kairei.[14] Высота оболочки 7,65–30,87 мм (0,30–1,22 дюйма).[2] Ширина апертуры составляет 7,26–32,52 мм (0,29–1,28 дюйма).[2] Высота апертуры составляет 6,38–27,29 мм (0,25–1,07 дюйма).[2]
Необычна и раковина улитки. Структура оболочки состоит из трех слоев. Внешний слой черного цвета толщиной около 30 мкм состоит из сульфидов железа, содержащих Greigite Fe3S4.[26] Эта особенность делает этого брюхоногого моллюска единственным известным до сих пор животным, которое использует этот материал в своем скелете.[2] Средний слой (около 150 мкм) эквивалентен органическому периостракуму, который также встречается у других брюхоногих моллюсков.[26] Периостракум толстый, коричневый.[2] Самый внутренний слой состоит из арагонит (толщиной около 250 мкм), форма карбонат кальция это обычно встречается как в раковинах моллюсков, так и в различных кораллах.[26] Цвет слоя арагонита - молочно-белый.[2]
Кажется, что каждый слой панциря по-разному способствует эффективности защиты улитки. Средний органический слой, по-видимому, поглощает механическое напряжение и энергию, генерируемую атакой сжатия (например, клешнями краба), что делает панцирь намного более прочной. Органический слой также рассеивает тепло.[27] Особенности этого композитный материал находятся в центре внимания исследователей для возможного использования в гражданских и военных защитных приложениях.[26]
 Chrysomallon squamiferum из жерлового месторождения Кайрей. |  С. squamiferum из вентиляционного поля Solitaire. |
Operculum
У этого вида форма крышка в процессе роста изменяется от округлой формы у молодых до изогнутой формы у взрослых.[13] Относительный размер жаберной крышки уменьшается по мере роста особей.[4] Около половины взрослых улиток этого вида имеют жаберную крышечку среди склеритов в задней части животного.[13] Кажется вероятным, что склериты постепенно разрастаются и полностью покрывают всю стопу для защиты, а жаберная крышка теряет свою защитную функцию по мере роста животного.[13]
 Молодь с крышечкой, обозначенной красным указателем. Длина раковины около 2 мм. |  Жаберная крышка молодой улитки. Масштабная линейка 1 мм. |  Жаберная крышка взрослой улитки. Масштабная линейка 1 мм. |  Взрослые улитки с крышечкой, обозначенной красными стрелками. Масштабная линейка 5 мм. |
Внешняя анатомия
sn - морда,
тт - головное щупальце.
Шкала 2 мм.
У чешуйчатого брюхоногого моллюска толстая морда, сужающаяся дистально к тупому концу. В сжатом и закрытом состоянии рот представляет собой круговое кольцо мышц.[4] Два гладких головных щупальца толстые у основания и постепенно сужаются к тонкому кончику на дистальных концах.[4] У этой улитки нет глаз.[4] Специализированного копулятивного придатка нет.[4] Нога красная и большая, и улитка не может полностью погрузить ногу в раковину.[2] Здесь нет педальный сальник в передней части стопы.[4] Эпиподиальных щупалец также нет.[4]
Внутренняя анатомия
В Chrysomallon squamiferumмягкие части животного занимают примерно два оборота внутренней части панциря.[4] Панцирная мышца имеет форму подковы и большая, разделена на две части слева и справа и соединена более узким прикреплением.[4] Край мантии толстый, но простой, без каких-либо отличительных особенностей.[4] В полость мантии глубокая и достигает заднего края раковины.[4] С медиальной левой стороны полости преобладает очень большой двояковыпуклый ктенидий.[4] Вентрально по отношению к висцеральной массе полость тела занята огромной пищеводной железой, которая расширяется, заполняя вентральное дно мантийной полости.[4][25]
В пищеварительная система прост, и его размер составляет менее 10% от объема, типичного для брюхоногих моллюсков.[4][25] В радула «слабый», типа рипидоглоссана, с единственной парой радулярных хрящей.[4][25] В формула радулы составляет ∼50 + 4 + 1 + 4 + ∼50.[2] Лента радула имеет длину 4 мм, ширину 0,5 мм;[2] отношение ширины к длине составляет примерно 1:10.[4] Здесь нет челюсть, а слюнных желез нет.[4] Часть передней части пищевода быстро разрастается в огромную гипертрофированную пищеводную железу с слепым концом, которая занимает большую часть вентральной стороны мантийной полости (приблизительно 9,3% объема тела).[4] Когда улитка растет, увеличивается пищеводная железа изометрически с ростом.[23] Пищеводная железа имеет однородную структуру и сильно васкуляризована с мелкими кровеносными сосудами.[4] В желудок имеет по крайней мере три протока спереди справа, соединяющиеся с пищеварительная железа.[4] Консолидированные гранулы находятся как в желудке, так и в задней части кишечника.[4] Эти гранулы, вероятно, представляют собой гранулы сера вырабатывается эндосимбионтом как способ детоксикации сероводорода.[4] Кишечник редуцирован и имеет только одну петлю.[4] Обширная и рыхлая пищеварительная железа простирается к задней части, заполняя оболочку. вершина оболочки.[4] Прямая кишка не проникает в сердце, а проходит к нему вентрально.[4] В анус находится с правой стороны улитки, над половым отверстием.[4]
в выделительная система, то нефридиум находится в центре, стремится к правой стороне тела в виде тонкого темного слоя железистой ткани.[4] Нефридий находится спереди и снизу от пищеварительной железы и контактирует с дорсальной стороной передней кишки.[4]
В дыхательная система и сердечно-сосудистая система состоят из одного левого двояковыпуклого ктенидий (жабры), которые очень велики (15,5% от объема тела) и поддерживаются множеством больших и подвижных кровеносных пазух, заполненных гемоцель.[4][23] При вскрытии кровяные пазухи и комки материала гемоцель являются заметным элементом по всей полости тела.[4] Хотя кровеносная система в Хризомаллон в основном закрытый (это означает, что гемоцель в основном не выходит из кровеносных пазух), выступающие кровеносные пазухи кажутся временными и возникают в разных частях тела у разных людей.[23] По обе стороны от ктенидия тонкие жаберные нити.[4] Бипектинатный ктенидий заходит далеко за сердце в верхние обороты раковины; это намного больше, чем в Пельтоспира. Хотя этот вид имеет форму раковины и общую форму, аналогичную другим пельтоспиридам, размер ктенидия пропорционален размеру раковины. Hirtopelta, у которого самая большая жабра среди родов пельтоспирид, анатомически исследованных до сих пор.[4]
Ктенидиум обеспечивает улитку кислородом, но кровеносная система расширена за пределы возможностей других подобных брюхоногих моллюсков.[4] В жабрах и на жабрах эндосимбионтов нет. С. squamiferum.[4] Увеличение жабр, вероятно, должно способствовать извлечению кислорода в условиях низкого содержания кислорода, которые типичны для экосистем гидротермальных источников.[4]
На задний ктенидия - очень большое и хорошо развитое сердце.[4] Сердце необычно велико для любого животного пропорционально.[4] Исходя из объема одного предсердия и желудочка, комплекс сердца составляет примерно 4% от объема тела (например, сердце человека составляет 1,3% от объема тела).[4] Желудочек имеет размер 0,64 мм у молодых животных с длиной раковины 2,2 мм, а у взрослых животных размер желудочка достигает 8 мм.[23] Это пропорционально гигантское сердце в основном всасывает кровь через ктенидий и снабжает кровеносную систему пищеводной железы.[4] В С. squamiferum эндосимбионты размещаются в пищеводной железе, где они изолированы от вентральной жидкости.[4] Таким образом, хозяин, вероятно, будет играть важную роль в снабжении эндосимбионтов необходимыми химическими веществами, что приведет к увеличению респираторных потребностей.[4] Детальное исследование гемоцела С. squamiferum раскроет дополнительную информацию о его дыхательных пигментах.[4]

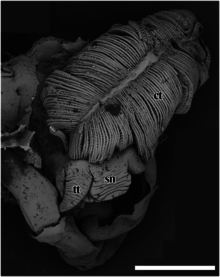
ct - ктенидий,
pm - педальная мышца,
sc - чешуя,
си - кровяной синус,
те - яички.
Масштабная линейка 1 см.
Брюхоногие моллюски на ножках являются хемосимбиотиками. холобионт.[25] Здесь обитают тиоавтотрофные (окисляющие серу) гаммапротеобактериальные эндосимбионты в значительно увеличенном масштабе. пищеводная железа, и, похоже, полагается на этих симбионтов для питания.[25][28] Ближайший известный родственник этого эндосимбионта - родственник Альвиниконча улитки.[29] У этого вида размер пищеводной железы примерно на два порядка больше обычного.[25] Внутри пищеводной железы имеется значительное разветвление, где артериальное давление, вероятно, снижается почти до нуля.[4] Сложная сердечно-сосудистая система, скорее всего, эволюционировала для насыщения кислородом эндосимбионтов в среде с низким содержанием кислорода и / или для обеспечения сероводород эндосимбионтам.[4] Тиоавтотрофные гаммапротеобактерии имеют полный набор генов, необходимых для аэробного дыхания, и, вероятно, способны переключаться между более эффективным аэробным дыханием и менее эффективным анаэробным дыханием в зависимости от наличия кислорода.[4] В 2014 году эндосимбионт брюхоногих моллюсков стал первым эндосимбионтом среди всех брюхоногих моллюсков, для которых был известен полный геном.[28] С. squamiferum ранее считалось, что это единственный вид Peltospiridae, который имеет увеличенную пищеводную железу,[2] но позже было обнаружено, что оба вида Гигантопельта также имеют увеличенную пищеводную железу.[10] Хризомаллон и Гигантопельта единственные вентиляционные животные, кроме сибоглинид трубчатые черви, которые содержат эндосимбионтов в закрытой части тела, не находящейся в прямом контакте с вентиляционной жидкостью.[23]
В нервная система большой, а мозг представляет собой твердую нервную массу без ганглиев.[4] Нервная система уменьшена по сложности и увеличена в размерах по сравнению с другими таксонами неомфалина.[4] Как это типично для брюхоногих моллюсков, нервная система состоит из переднего пищеводного нервного кольца и двух пар продольных нервных тяжей, вентральная пара иннервирует стопу, а дорсальная пара образует изгиб через стрептонерия.[4] Фронтальная часть кольца пищеводного нерва большая, соединяет два боковых вздутия.[4] Огромная сросшаяся нервная масса непосредственно примыкает к пищеводно-фагальной железе, где находятся бактерии, и проходит через нее.[4] В головные щупальца входят большие щупальцевые нервы.[4] В органы чувств брюхоногих моллюсков включают статоцисты окружены пищеводной железой, каждая статоциста с одним статолит.[4] На концах жаберных нитей также имеются сенсорные ктенидиальные бурсикулы; они, как известно, присутствуют у большинства ветигастропод и присутствуют в некоторых неомфалинах.[4]
В репродуктивная система имеет некоторые необычные особенности. Гонады взрослых улиток не находятся внутри раковины; они находятся в области голова-стопа на правой стороне тела.[4] У молоди с длиной раковины 2,2 мм гонад отсутствует.[23] Взрослые обладают обоими яички и яичник на разных уровнях развития.[4] Яичко расположено вентрально; яичник расположен дорсально, а нефридиум лежит между ними.[4] Рядом с яичком находится «упаковывающий орган сперматофора».[4] Гоноды от семенников и яичников изначально разделены, но, по-видимому, сливаются в один проток и появляются как единое генитальное отверстие справа от мантийной полости.[4] Копулятивного органа у животного нет.[2][4]
 Вид сзади Chrysomallon squamiferum с изображением мантийной полости. Ткань панциря и мантии удалены. Шкала шкалы 1 см. |  3D реконструкция показывает большой ктенидий и большое сердце, вид сверху. Масштабная линейка 250 мкм.
|  Трехмерная реконструкция пищеварительной системы показывает увеличенную пищеводную железу, вид сверху. Масштабная линейка 250 мкм.
|
Предполагается, что производная стратегия размещения эндосимбиотических микробов в пищеводе стала катализатором анатомических инноваций, которые служат в первую очередь для улучшения приспособленности бактерий, помимо потребностей улитки.[4] Большое увеличение пищеводной железы, защитные кожные склериты улитки, ее сильно увеличенная дыхательная и кровеносная системы и ее высокая плодовитость - все это считается адаптацией, полезной для ее эндосимбионтных микробов.[4] Эти адаптации, по-видимому, являются результатом специализации для удовлетворения энергетических потребностей в крайнем случае. хемосинтетический среда.[4]
Экология
Среда обитания

Обитает в гидротермальных жерловых полях Индийского океана. Он живет рядом как с кислой, так и с восстанавливающей вентиляционной жидкостью, на стенках дымовых труб черного курильщика или непосредственно на участках диффузного потока.[4]
Глубина месторождения Кайрей варьируется от 2415 до 2460 м (от 7923 до 8071 футов),[7] и его размеры примерно 30 на 80 м (98 на 262 фута).[7] Наклон поля от 10 ° до 30 °.[7] В субстрат рок это троктолит и истощенный срединно-океанский хребет базальт.[30] Чешуйчатые брюхоногие моллюски поля Кайрей живут в низкотемпературных диффузных жидкостях одного дымохода.[13] Переходная зона, где были найдены эти брюхоногие моллюски, составляет около 1-2 м (3-7 футов) в ширину, с температурой 2-10 ° C.[31] Предпочтительная температура воды для этого вида составляет около 5 ° C.[32] Эти улитки живут в среде с высокой концентрацией сероводорода и низкой концентрацией кислорода.[32]
Численность чешуйчатых брюхоногих моллюсков на поле Кайрей была ниже, чем на поле Лунци.[2] Сообщество гидротермального источника Кайрей состоит из 35 таксонов,[33] включая морские анемоны Марианактис sp., ракообразные Остиногрея родригезенсис, Rimicaris kairei, Мирокарис индика, Мунидопсис sp., Neolepadidae род и вид, Эохионелазм sp., двустворчатые моллюски Батимодиолус марисиндикус, брюхоногие моллюски Лепетодрилус sp., Псевдоримула sp., Эулепетопсис sp., Шинкаилепас sp., и Alviniconcha marisindica,[34] Desbruyeresia marisindica,[35] Брюсейелла Варени,[35] Phymorhynchus sp., Сутилизона sp., щелевое блюдце sp. 1, щелевое блюдце зр. 2, Ифинопсис бушети,[35] соленогастр Геликорадомения ? sp., кольчатые червя Амфисамита sp., Арчином Джейсони, Capitellidae sp. 1, Офётроча sp., Гесионоиды sp. 1, Гесионоиды sp. 2, Бранчинотоглума sp., Бранчиполиное sp., Harmothoe ? sp., Levensteiniella ? sp., Прионоспио sp., неопознанный Немертя и неопознанный Платигельминты.[33] Чешуйчатые брюхоногие моллюски живут колониями с Alviniconcha marisindica улиток, есть колонии Rimicaris kairei над ними.[32]
Поле Solitaire находится на глубине 2606 м (8550 футов), а его размеры составляют примерно 50 на 50 м (160 на 160 футов).[13] Скальный субстрат - обогащенный базальтом срединно-океанического хребта.[13][30] Чешуйчатые брюхоногие моллюски живут рядом с высокотемпературными диффузными жидкостями дымовых труб в вентиляционном поле.[13] Численность чешуйчатых брюхоногих моллюсков была ниже, чем на месторождении Лунци.[2] Сообщество гидротермальных источников Solitaire включает 22 таксона, в том числе морские анемоны. Марианактис sp., ракообразные Остиногрея родригезенсис, Rimicaris kairei, Мирокарис индика, Мунидопсис sp., Neolepadidae gen et sp., Эохионелазм sp., двустворчатые моллюски Батимодиолус марисиндикус, брюхоногие моллюски Лепетодрилус sp., Эулепетопсис sp., Шинкаилепас sp., Альвиниконча sp. тип 3, Desbruyeresia sp., Phymorhynchus sp., кольчатые червя Альвинеллиды род и вид, Арчином Джейсони, Бранчинотоглума sp., иглокожие голотурии Аподацея gen et sp., рыба Макрурусовые род и вид, не идентифицированы Немертя, и неопознанные Платигельминты.[33]

Жидкое месторождение Longqi находится на глубине 2780 м (9120 футов),[2] и его размеры примерно 100 на 150 м (330 на 490 футов).[19] Chrysomallon squamiferum была густо заселена в районах, непосредственно окружающих выход диффузного потока.[4] Сообщество гидротермальных источников Лунци включает 23[Примечание 1] Таксоны макро- и мегафауны: морские анемоны Актиностолиды sp., кольчатые червя Полиноиды п. ген. п. sp. «655», Бранчиполиное п. sp. "Дракон", Пейналеополыное п. sp. "Дракон", Гезиолира ср. Берги, Hesionidae sp. indet., Офриотроча п. sp. «Ф-038 / 1б», Прионоспио ср. однослойный, Амфаретиды sp. индет., мидии Батимодиолус марисиндикус, брюхоногие моллюски Gigantopelta aegis,[10] Dracogyra subfuscus, Лирапекс политус,[15] Phymorhynchus п. sp. «SWIR», Лепетодрилус п. sp. «SWIR», ракообразные Неолепас sp. 1, Rimicaris kairei, Мирокарис индика, Chorocaris sp., Кива п. sp. «SWIR» 17, г. Мунидопсис sp. и иглокожие голотурии Чиридота sp.[16][36] Плотность Лепетодрилус п. sp.«SWIR» и чешуйчатые брюхоногие моллюски составляют более 100 улиток на м² в непосредственной близости от источников жидкости на жерловом поле Longqi.[36]
Кормление
Брюхоногие моллюски с чешуйчатой стопой симбиотроф на протяжении всей постпоселковой жизни.[23] Питание чешуйчатого брюхоногого моллюска на протяжении всей его постличиночной жизни зависит от химиоавтотрофия эндосимбиотических бактерий, которые обеспечивают все его питание.[25][23] Чешуйчатый брюхоногий моллюск не является фильтром[4][23] и не использует другие механизмы для кормления.[4] Радула составляет только 0,4% от объема тела молодых особей, а хрящи радулы составляют 0,8% от объема тела молодых особей,[23] потому что они больше не используются для кормления.
Для идентификации трофические взаимодействия в местообитаниях, где прямое наблюдение за питанием затруднено, были измерены составы стабильных изотопов углерода и азота.[31] Есть истощенные значения δ13C в пищеводе (по отношению к органическому углероду, полученному в результате фотосинтеза).[25] Источником такого углерода считались хемоавтотрофные симбионты.[25] Хемоавтотрофное происхождение стабильного изотопа углерода 13C подтверждено экспериментально.[28]
| ткань | δ13C | δ15N |
|---|---|---|
| пищеводная железа | −20.7 ± 0.9 ‰ | 3.3 ± 1.8 ‰ |
| жабры | −18,3 ± 0,6, от −17,4 до −18,8 ‰ | 3,9 ± 0,6, с 3,1 до 4,2 ‰ |
| мантия | от −17,5 до −18,6 ‰ | от 3,5 до 4,7 ‰ |
| оплачивать | −18.2 ± 0.6 ‰ | 3.8 ± 0.5 ‰ |
| напольные весы | −16.7 ± 0.6 ‰ | 3.8 ± 0.9 ‰ |
Жизненный цикл
Этот брюхоногий моллюск - одновременный гермафродит.[4] Это единственный вид семейства Peltospiridae, который, как известно, является одновременным гермафродитом.[4] Имеет высокий плодовитость.[4] Откладывает яйца, вероятно, лецитотрофного типа.[21] Яйца чешуйчатых брюхоногих моллюсков обладают отрицательной плавучестью при атмосферном давлении.[14] Ни личинки, ни протоконх известен по состоянию на 2016 год, но считается, что этот вид находится на стадии планктонного распространения.[21] Наименьший Chrysomallon squamiferum когда-либо собранные молодые особи имели длину раковины 2.2 мм.[23] Результаты статистического анализа не выявили генетической дифференциации между двумя популяциями в месторождениях Кайрей и Солитер, что предполагает потенциальную связь между двумя жерловыми полями.[14] Популяция Кайрей представляет собой потенциальную исходную популяцию для двух популяций Центрально-Индийского хребта.[14] Этих улиток сложно сохранить в искусственной среде, однако они выжили в аквариумах при атмосферном давлении более трех недель.[32]
Меры по сохранению и угрозы
Чешуйчатый брюхоногий моллюск не защищен.[1][19] Его общая площадь распространения составляет не более 0,27 км.2, ограничивается тремя зарегистрированными участками, между которыми происходит лишь незначительная миграция.[37] Гидротермальные жерла на юго-западе Индийского хребта распространяются медленно, и его сообщества считаются более чувствительными к нарушениям и с медленными темпами восстановления.[19] Этот вид подвержен риску нанесения вреда окружающей среде. глубоководная добыча.[19] Коммерческий лицензия на разведку полезных ископаемых в район Кайрей был пожалован Международный орган по морскому дну в Германию с 2015 по 2030 год.[19] Лицензия на коммерческую разведку полезных ископаемых на участке Лунци была предоставлена Китаю с 2011 по 2026 год.[19] Он был занесен в список исчезающих видов в Красный список видов, находящихся под угрозой исчезновения МСОП 4 июля 2019 г.[1]
Примечания
- ^ По состоянию на 2016 год из Лунци был известен 21 вид, а в 2017 были описаны два новых брюхоногих моллюска.
Рекомендации
Эта статья включает текст Creative Commons (CC-BY-4.0) из ссылок[4][14][23] и текст CC-BY-2.5 из справочника[13]
- ^ а б c «Красный список видов, находящихся под угрозой исчезновения МСОП». https://www.iucnredlist.org/species/103636217/103636261. 2019. Получено 2019-07-18. Внешняя ссылка в
| сайт =(помощь) - ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф аг ах ай эй ак аль являюсь ан ао ap водный ар в качестве в Чен, Чонг; Линсе, Катрин; Копли, Джонатан Т .; Роджерс, Алекс Д. (2015). «Чешуйчатые брюхоногие моллюски: новый род и вид гидротермальных брюхоногих моллюсков (Neomphalina: Peltospiridae) из Индийского океана». Журнал исследований моллюсков. 81 (3): 322–334. Дои:10.1093 / mollus / eyv013.
- ^ Зигварт, Джулия Д .; Чен, Чонг; Thomas, Elin A .; Оллкок, А. Луиза; Бём, Моника; Седдон, Мэри (22.07.2019). «Красный список может защитить глубоководное биоразнообразие». Природа Экология и эволюция. 3 (8): 1134. Дои:10.1038 / s41559-019-0930-2. ISSN 2397-334X. PMID 31332328.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф аг ах ай эй ак аль являюсь ан ао ap водный ар в качестве в au средний ау топор ай az ба bb до н.э bd быть парень bg бх би Ъ bk бл бм млрд бо бп бк br bs bt бу bv чб Чен, Чонг; Копли, Джонатан Т .; Линсе, Катрин; Роджерс, Алекс Д .; Зигварт, Джулия Д. (2015). «Сердце дракона: трехмерная анатомическая реконструкция« чешуйчатого брюхоногого моллюска »(Mollusca: Gastropoda: Neomphalina) показывает его необычную систему кровообращения». Границы зоологии. 12: 13. Дои:10.1186 / s12983-015-0105-1. ЧВК 4470333. PMID 26085836.
- ^ «Красный список видов, находящихся под угрозой исчезновения МСОП». Красный список видов, находящихся под угрозой исчезновения МСОП. Получено 2019-07-28.
- ^ Ламберт, Джонатан (22.07.2019). «Океанская улитка - первое животное, которое официально находится под угрозой исчезновения из-за глубоководной добычи полезных ископаемых». Природа. 571 (7766): 455–456. Дои:10.1038 / d41586-019-02231-1. PMID 31337912.
- ^ а б c d е ж грамм Довер, Синди Л. Ван; Humphris, S.E .; Fornari, D .; Cavanaugh, C.M .; Collier, R .; Гоффреди, Шана К .; Hashimoto, J .; Lilley, M.D .; Reysenbach, A. L .; Shank, T. M .; Фон Дамм, К. Л .; Banta, A .; Gallant, R.M .; Gotz, D .; Green, D .; Hall, J .; Harmer, T. L .; Уртадо, Л. А .; Johnson, P .; McKiness, Z. P .; Meredith, C .; Olson, E .; Pan, I. L .; Репа, М .; Выиграл, Y .; Янг, К. Р. 3-й; Вриенхук, Р. К. (2001). «Биогеография и экологическая обстановка гидротермальных источников Индийского океана». Наука. 294 (5543): 818–23. Bibcode:2001Наука ... 294..818В. Дои:10.1126 / science.1064574. PMID 11557843.
- ^ Буше, П. (2014). "Chrysomallon squamiferum". Всемирный регистр морских видов. Получено 2015-04-22.
- ^ а б Варен, Андерс; Бенгтсон, Стефан; Гоффреди, Шана К .; Довер, Синди Л. Ван (2003). «Горячий брюхоногий моллюск с кожными склеритами из сульфида железа». Наука. 302 (5647): 1007. Дои:10.1126 / science.1087696. PMID 14605361.
- ^ а б c Чен, Чонг; Линсе, Катрин; Ротерман, Кристофер Н .; Копли, Джонатан Т .; Роджерс, Алекс Д. (2015). «Новый род крупных гидротермальных эндемичных брюхоногих моллюсков (Neomphalina: Peltospiridae)» (PDF). Зоологический журнал Линнеевского общества (Представлена рукопись). 175 (2): 319–335. Дои:10.1111 / zoj.12279.
- ^ (на японском языке) (2010) "硫化 鉄 を 纏 わ な い 白 ス ケ フ ッ ト を 世界 で て 発 見 ~ イ 洋 に け る 新 規 熱水 探査 の 成果 ~". Японское агентство морских наук и технологий, Токийский университет, Kōchi University. (пресс-релиз). Проверено 16 июля 2016.
- ^ «Новый чешуйчатый брюхоногий моллюск найден в Индийском океане; открытие белого чешуйчатого брюхоногого моллюска». Южная жареная наука. 1 июля 2011 г.. Получено 2016-07-16.
- ^ а б c d е ж грамм час я j k л м Накамура, Кентаро; Ватанабэ, Хироми; Миядзаки, Дзюнъити; Такай, Кен; Кавагуччи, Шинсуке; Ногути, Такуро; Немото, Сугуру; Вацудзи, Томо-о; Мацудзаки, Такуя; Сибуя, Такадзо; Окамура, Кей; Мотидзуки, Масаси; Орихаши, Юджи; Ура, Тамаки; Асада, Акира; Мари, Даниэль; Кунджул, Мира; Сингх, Манвендра; Бедесси, Гириш; Бхикаджи, Митрасен; Тамаки, Кенсаку; Шнур, Джоэл М. (2012). «Открытие новой гидротермальной активности и хемосинтетической фауны на Центрально-Индийском хребте на 18–20 ° ю.ш.». PLOS ONE. 7 (3): e32965. Bibcode:2012PLoSO ... 732965N. Дои:10.1371 / journal.pone.0032965. ЧВК 3303786. PMID 22431990.
- ^ а б c d е ж Бедесси, Гириш; Ватанабэ, Хироми; Огура, Томоми; Немото, Сугуру; Яхаги, Такуя; Накагава, Сатоши; Накамура, Кентаро; Такай, Кен; Кунджул, Мира; Мари, Дэниел Э. П. (2013). «Высокая взаимосвязанность популяций животных в глубоководных гидротермальных жерловых полях в Центрально-Индийском хребте, имеющая отношение к его геологическим условиям». PLOS ONE. 8 (12): e81570. Bibcode:2013PLoSO ... 881570B. Дои:10.1371 / journal.pone.0081570. ЧВК 3864839. PMID 24358117.
- ^ а б Чен, Чонг; Чжоу, Ядун; Ван, Чуньшэн; Копли, Джонатан Т. (2017). «Две новые пельтоспиридные улитки с горячим выходом (Gastropoda: Neomphalina) из гидротермального поля Лунци, юго-западный Индийский хребет». Границы морских наук. 4. Дои:10.3389 / fmars.2017.00392. ISSN 2296-7745.
- ^ а б c Тао, Чуньхуэй; Линь, Цзянь; Го, Шицинь; Чен, Юншун Джон; Ву, Гуанхай; Хан, Xiqiu; Герман, Кристофер Р .; Yoerger, Dana R .; Чжоу, Нин; Ли, Хуайминь; Су, Синь; Чжу, Цзянь (2012). DY115-19 (этапы 1–2) и DY115-20 (этапы 4–7) Научные вечеринки. «Первые активные гидротермальные источники в центре сверхмедленного спрединга: Юго-западный Индийский хребет». Геология. 40 (1): 47–50. Bibcode:2012Гео .... 40 ... 47 т. Дои:10.1130 / G32389.1.
- ^ а б c d е ж грамм час я Чен, Чонг; Копли, Джонатан Т .; Линсе, Катрин; Роджерс, Алекс Д .; Зигварт, Джулия (2015). «Как у моллюска появились свои чешуйки: конвергентная эволюция склеритома моллюска». Биологический журнал Линнеевского общества. 114 (4): 949–954. Дои:10.1111 / bij.12462.
- ^ Роджерс, Алекс Д .; Тайлер, Пол А .; Коннелли, Дуглас П .; Копли, Джон Т .; Джеймс, Рэйчел; Лартер, Роберт Д.; Линсе, Катрин; Миллс, Рэйчел А .; Гарабато, Альфредо Навейра; Панкост, Ричард Д .; Пирс, Дэвид А .; Полунин, Николай В. С .; Герман, Кристофер Р .; Шанк, Тимоти; Boersch-Supan, Philipp H .; Алкер, Белинда Дж .; Акилина, Альфред; Беннет, Сара А .; Кларк, Эндрю; Динли, Роберт Дж. Дж .; Graham, Alastair G.C .; Грин, Дэррил Р. Х .; Хоукс, Джеффри А .; Хепберн, Лаура; Иларио, Ана; Huvenne, Veerle A. I .; Марш, Ли; Рамирес-Льодра, Ева; Рид, Уильям Д. К .; Ротерман, Кристофер Н .; Свитинг, Кристофер Дж .; Татже, Свен; Цвирглмайер, Катрин (2012). «Открытие новых сообществ глубоководных гидротермальных жерл в Южном океане и их значение для биогеографии». PLOS Биология. 10 (1): –1001234. Дои:10.1371 / journal.pbio.1001234. ISSN 1545-7885. ЧВК 3250512. PMID 22235194.
- ^ а б c d е ж грамм Зигварт, Джулия Д .; Чен, Чонг; Марш, Ли (2017). «Вредна ли разработка морского дна для моллюсков?». Наутилус. 131 (1): 43–49.
- ^ а б Хашимото, Джун; Охта, Сугуру; Гамо, Тошитака; Тиба, Хитоши; Ямагути, Тосиюки; Цучида, Синдзи; Окудаира, Такамото; Ватабе, Хадзиме; Яманака, Тоширо; Китадзава, Мицуко (2001). «Обнаружены первые сообщества гидротермальных источников в Индийском океане». Зоологическая наука. 18 (5): 717–721. Дои:10.2108 / zsj.18.717.
- ^ а б c Чен, Чонг; Копли, Джонатан Т .; Линсе, Катрин; Роджерс, Алекс Д. (2015). «Низкая взаимосвязь между популяциями« чешуйчатых брюхоногих моллюсков »(Mollusca: Peltospiridae) в гидротермальных жерлах на юго-западе Индийского хребта и Центрально-Индийском хребте». Разнообразие и эволюция организмов. 15 (4): 663–670. Дои:10.1007 / s13127-015-0224-8.
- ^ Пикрелл, Джон (07.11.2003). "Бронированная улитка, обнаруженная в глубоком море". Новости National Geographic. Получено 2016-07-16.
- ^ а б c d е ж грамм час я j k л м Чен, Чонг; Уэмацу, Кацуюки; Линсе, Катрин; Зигварт, Джулия Д. (2017). "По-разному: быстрое схождение в гидротермальных жерлах, показанное на трехмерной анатомической реконструкции Гигантопельта (Mollusca: Neomphalina) ". BMC Эволюционная биология. 17 (1): 62. Дои:10.1186 / s12862-017-0917-z. ISSN 1471-2148. ЧВК 5333402. PMID 28249568.
- ^ Сузуки, Йохей; Копп, Роберт Э .; Коруге, Тошихиро; Шуга, Акинобу; Такай, Кен; Цучида, Синдзи; Одзаки, Нориаки; Эндо, Кадзуёси; Хашимото, Джун; Като, Ясухиро; Мизота, Читоши; Хирата, Такафуми; Тиба, Хитоши; Nealson, Kenneth H .; Хорикоши, Коки; Киршвинк, Джозеф Л. (2006). «Образование склерита в гидротермальном источнике брюхоногих моллюсков« чешуйчатая ножка »- возможный контроль животным биоминерализации сульфида железа» (PDF). Письма по науке о Земле и планетах. 242 (1–2): 39–50. Bibcode:2006E и PSL.242 ... 39S. Дои:10.1016 / j.epsl.2005.11.029.
- ^ а б c d е ж грамм час я j k л м Гоффреди, Шана К .; Варен, Андерс; Сирота, Виктория Дж .; Довер, Синди Л. Ван; Вриенхук, Роберт К. (5 мая 2004 г.). «Новые формы структурной интеграции микробов и брюхоногих моллюсков с гидротермальными источниками из Индийского океана». Прикладная и экологическая микробиология. 70 (5): 3082–3090. Дои:10.1128 / AEM.70.5.3082-3090.2004. ЧВК 404406. PMID 15128570.
- ^ а б c d Яо, Хайминь; Дао, Мин; Имхольт, Тимоти; Хуанг, Джейми; Уиллер, Кевин; Бонилла, Алехандро; Суреш, Субра; Ортис, Кристина (2010). «Механизмы защиты железной брони брюхоногих моллюсков из глубоководных гидротермальных источников». PNAS. 107 (3): 987–992. Bibcode:2010ПНАС..107..987Г. Дои:10.1073 / pnas.0912988107. ЧВК 2808221. PMID 20133823.
- ^ "Железная броня Улитки глазами военных". CBC News. 2010-01-19. Получено 2016-07-16.
- ^ а б c Накагава, Сатоши; Шимамура, Сигеру; Такаки, Ёсихиро; Сузуки, Йохей; Мураками, Сюн-ичи; Ватанабэ, Тамаки; Фудзиёси, Итак; Мино, Саяка; Савабе, Томоо; Маэда, Такахиро; Макита, Хироко; Немото, Сугуру; Нисимура, Син-Ичиро; Ватанабэ, Хироми; Вацудзи, Томо-о; Такай, Кен (2014). «Союз с панцирными улитками: полный геном гаммапротеобактериального эндосимбионта». Журнал ISME. 8 (1): 40–51. Дои:10.1038 / ismej.2013.131. ЧВК 3869010. PMID 23924784.
- ^ Distel, Daniel L .; Altamia, Marvin A .; Линь, Чжэньцзянь; Шипвей, Дж. Рубен; Хан, Эндрю; Фортеза, Имельда; Антемано, Ровена; Лимбако, Ма Гвен Дж. Пеньяфлор; Тебо, Элисон Дж .; Дечавес, Ранде; Альбано, Джули; Розенберг, Гэри; Concepcion, Gisela P .; Шмидт, Эрик У .; Хейгуд, Марго Г. (2017-04-17). «Открытие хемоавтотрофного симбиоза у гигантского корабельного червя Kuphus polythalamia (Bivalvia: Teredinidae) расширяет теорию деревянных ступенек». Труды Национальной академии наук. 114 (18): E3652 – E3658. Дои:10.1073 / pnas.1620470114. ISSN 1091-6490. ЧВК 5422788. PMID 28416684. Получено 2017-04-18.
- ^ а б Накамура, Кентаро; Такай, Кен (2015). «Гидротермальные системы Индийского океана: гидротермальная деятельность на морском дне, физические и химические характеристики гидротермальных флюидов и биологические сообщества, связанные с вентиляционными отверстиями». In Ishibashi J.-i .; и другие. (ред.). Подводная биосфера, связанная с гидротермальными системами. Спрингер, Токио. С. 147–161. Дои:10.1007/978-4-431-54865-2_12. ISBN 9784431548645.
- ^ а б c Довер, Синди Ван (2002). «Трофические взаимоотношения между беспозвоночными на гидротермальном поле Кайрей (Центральный Индийский хребет)». Морская биология. 141 (4): 761–772. Дои:10.1007 / s00227-002-0865-у.
- ^ а б c d «Обнаружена обширная популяция« редкого »чешуйчатого брюхоногого моллюска». Японское агентство морских наук и технологий, Университет Хоккайдо, Аквариум Эношима. 30 ноября 2009 г.. Получено 2016-07-16.
- ^ а б c Ватанабэ, Хироми; Бидесси, Гириш (2015). «Вентиляционная фауна Центрального Индийского хребта». In Ishibashi J.-i .; и другие. (ред.). Подводная биосфера, связанная с гидротермальными системами. Спрингер, Токио. С. 205–212. Дои:10.1007/978-4-431-54865-2_16. ISBN 9784431548645.
- ^ Джонсон, Шеннон Б .; Варен, Андерс; Танниклифф, Верена; Довер, Синди Ван; Пшеница, К. Джеффри; Шульц, Томас Ф .; Вриенхук, Роберт К. (04.05.2015). "Молекулярная таксономия и названия пяти загадочных видов Альвиниконча улитки (Gastropoda: Abyssochrysoidea) из гидротермальных источников ". Систематика и биоразнообразие. 13 (3): 278–295. Дои:10.1080/14772000.2014.970673. ISSN 1477-2000.
- ^ а б c Окутани, Такаши; Хашимото, Джун; Сасаки, Такенори (2004). «Новые таксоны брюхоногих моллюсков из гидротермального источника (месторождение Кайрей) в центральной части Индийского океана» (PDF). Венера. 63 (1–2): 1–10. Архивировано из оригинал (PDF) на 2013-10-04.
- ^ а б Копли, Дж. Т .; Marsh, L .; Glover, A. G .; Hühnerbach, V .; Най, В. Э .; Reid, W. D. K .; Свитинг, С. Дж .; Wigham, B.D .; Виклунд, Х. (2016). «Экология и биогеография мегафауны и макрофауны в первых известных глубоководных гидротермальных жерлах на сверхмедленном спрединге Юго-Западного Индийского хребта». Научные отчеты. 6: 39158. Bibcode:2016НатСР ... 639158C. Дои:10.1038 / srep39158. ISSN 2045-2322. ЧВК 5155287. PMID 27966649.
- ^ Зигварт, Джулия Д. (зима 2017 г.). "Охрана морских глубин и чешуйчатые брюхоногие моллюски"'" (PDF). Щупальце. 25: 39–40.