Глубоководное сообщество - Deep sea community


А глубоководное сообщество есть ли сообщество из организмы связаны общим среда обитания в глубокое море. Глубоководные сообщества остаются в значительной степени неизведанными из-за технологических и логистических проблем и затрат, связанных с посещением этого удаленного биом. Из-за уникальных проблем (особенно высоких барометрическое давление, экстремальные температуры и отсутствие света ), долгое время считалось, что в этой враждебной среде мало жизни. Однако с XIX века исследования показали, что биоразнообразие существует в глубоком море.
Три основных источника энергии и питательных веществ для глубоководных сообществ: морской снег, Кит падает, и хемосинтез в гидротермальные источники и холодные просачивания.
История
До XIX века ученые полагали, что в глубоком океане мало жизни. В 1870-е гг. Сэр Чарльз Вивилл Томсон и коллеги на борту Челленджер экспедиция обнаружил много глубоководных существ самых разных типов.
Первое открытие глубоководных хемосинтетический сообщество, включающее высших животных, неожиданно образовалось в гидротермальных источниках на востоке Тихий океан во время геологоразведочных работ (Corliss et al., 1979).[1] Два ученых, Дж. Корлисс и Дж. Ван Андел, впервые стали свидетелями плотных хемосинтетических пластов моллюсков из подводного аппарата. DSV Элвин 17 февраля 1977 года, после их неожиданного обнаружения двумя днями ранее с помощью телекамеры.[1]
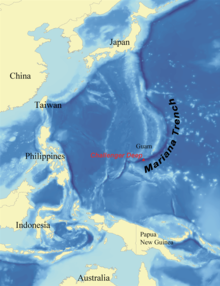
В Challenger Deep самая глубокая из исследованных точек всех океанов Земли; он расположен на южной оконечности Марианская впадина недалеко от Марианские острова группа. Депрессия названа в честь HMS Претендент, исследователи которого сделали первые записи глубины 23 марта 1875 г. вокзал 225. Сообщенная глубина составила 4475 сажень (8184 метра) на основе двух отдельных зондирований. В 1960 г. Дон Уолш и Жак Пикар спустился на дно Бездны Челленджера в Триест батискаф. На этой большой глубине была замечена небольшая рыба, похожая на камбалу, удаляющуюся от прожектора батискафа.
Японский дистанционно управляемый автомобиль (ROV) Кайко стал вторым судном, достигшим дна Challenger Deep в марте 1995 года. Нерей, гибридный дистанционно управляемый автомобиль (ГТС) Океанографическое учреждение Вудс-Хоул, является единственным транспортным средством, способным исследовать глубины океана за пределами 7000 метров. Нерей 31 мая 2009 г. достигла глубины 10902 метра в Глубине Челленджера.[2][3] 1 июня 2009 г. сонарное картирование Глубины Челленджера Симрад EM120 батиметрия многолучевого сонара система на борту R / V Кило Моана указал максимальную глубину 10 971 метр (6 817 миль). В сонарной системе используются фаза и амплитуда обнаружение дна с точностью лучше 0,2% от глубины воды (это ошибка около 22 метров на этой глубине).[3][4]
Среда
Тьма

Океан можно представить как разделенный на различные зоны в зависимости от глубины и наличия или отсутствия Солнечный свет. Почти все формы жизни в океане зависят от фотосинтетический деятельность фитопланктон и другие морские растения преобразовать углекислый газ в органический углерод, который является основным строительным блоком органическая материя. Фотосинтез, в свою очередь, требует энергии солнечного света для запуска химических реакций, которые производят органический углерод.[5]
Слой столб воды до которого проникает солнечный свет, называется световая зона. В световая зона можно разделить на две разные вертикальные области. Самая верхняя часть фотической зоны, где есть достаточно света для поддержки фотосинтеза фитопланктоном и растениями, называется эвфотическая зона (также называемый эпипелагическая зона, или же поверхностная зона).[6] Нижняя часть фотической зоны, где интенсивность света недостаточна для фотосинтеза, называется дисфотическая зона (дисфотический означает «плохо освещенный» по-гречески).[7] Дисфотическая зона также упоминается как мезопелагическая зона, или зона сумерек.[8] Самая нижняя его граница находится на термоклин 12 ° C (54 ° F), что в тропики обычно находится между 200 и 1000 метров.[9]
Эуфотическая зона несколько условно определяется как простирающаяся от поверхности до глубины, где интенсивность света составляет примерно 0,1–1% от солнечного света на поверхности. сияние, в зависимости от время года, широта и степень воды мутность.[6][7] В чистейшей океанской воде эвфотическая зона может достигать глубины около 150 метров.[6] реже - до 200 метров.[8] Растворенные вещества и твердые частицы поглощают и рассеивают свет, а в прибрежных районах высокая концентрация этих веществ приводит к быстрому ослаблению света с глубиной. В таких местах глубина эвфотической зоны может составлять всего несколько десятков метров или меньше.[6][8] Дисфотическая зона, где интенсивность света значительно меньше 1% поверхностной освещенности, простирается от основания эвфотической зоны примерно до 1000 метров.[9] Распространяется от нижней части фотической зоны до морское дно это афотическая зона, область вечной тьмы.[8][9]
Поскольку средняя глубина океана составляет около 4300 метров,[10] фотическая зона представляет собой лишь крошечную долю от общего объема океана. Однако из-за своей способности к фотосинтезу фотическая зона имеет наибольшее биоразнообразие и биомасса всех океанических зон. Почти все основное производство в океане встречается здесь. Любые формы жизни, присутствующие в афотической зоне, должны быть способны к движение вверх через толщу воды в фотическую зону для кормления, или должны полагаться на опускание материала сверху,[5] или должны найти другой источник энергии и питания, например, в хемосинтетический археи найдено рядом гидротермальные источники и холодные просачивания.
Гипербаричность
У этих животных есть развился выжить в экстремальном давлении суб-световые зоны. Давление увеличивается примерно на один атмосфера каждые десять метров. Чтобы справиться с напором, многие рыбы довольно мелкие, обычно не превышающие 25 см в длину. Кроме того, ученые обнаружили, что чем глубже живут эти существа, тем более студенистая их плоть и более минималистична их скелетная структура. Эти существа также устранили все лишние полости, которые могли разрушиться под давлением, например, плавательные пузыри.[11]
Давление является величайшим фактором окружающей среды, действующим на глубоководные организмы. В глубоком море, хотя большая часть глубоководного моря находится под давлением от 200 до 600 атм, диапазон давления составляет от 20 до 1000 атм. Давление играет большую роль в распространении глубоководных организмов. До недавнего времени людям не хватало подробной информации о прямых последствиях давления на большинство глубоководных организмов, потому что практически все организмы, выловленные тралом из морских глубин, попадали на поверхность мертвыми или умирающими. С появлением ловушек, включающих в себя специальную камеру для поддержания давления, неповрежденные большие многоклеточный животные были извлечены из морских глубин в хорошем состоянии. Некоторые из них были сохранены для экспериментальных целей, и мы получаем больше знаний о биологических эффектах давления.
Температура
Две области наибольшего и наиболее быстрого температура Изменения в океанах - это переходная зона между поверхностными водами и глубинными водами, термоклин и переход между глубоководным дном и потоками горячей воды в гидротермальных жерлах. Толщина термоклинов варьируется от нескольких сотен метров до почти тысячи метров. Ниже термоклина водная масса глубокого океана холодная и гораздо более однородная. Термоклины наиболее сильны в тропиках, где температура эпипелагическая зона обычно выше 20 ° C. От основания эпипелагии температура падает на несколько сотен метров до 5 или 6 ° C на высоте 1000 метров. Он продолжает снижаться к основанию, но скорость намного меньше. Ниже 3000-4000 м вода изотермический. На любой заданной глубине температура практически не меняется в течение длительного времени. Нет ни сезонных изменений температуры, ни годовых изменений. Никакая другая среда обитания на земле не имеет такой постоянной температуры.
Гидротермальные источники - прямой контраст с постоянной температурой. В этих системах температура воды, выходящей из дымовых труб «черного курильщика», может достигать 400 ° C (она удерживается от кипения за счет высокого гидростатического давления), а в пределах нескольких метров может снова упасть до 2–4 ° C.[12]
Соленость

Соленость постоянно на всех глубинах морских глубин. Из этого правила есть два заметных исключения:
- в Средиземное море, потеря воды через испарение значительно превышает вклад от осадки и речной сток. Из-за этого соленость в Средиземном море выше, чем в Атлантический океан.[13] Испарение особенно велико в его восточной половине, что вызывает снижение уровня воды и повышение солености в этой области.[14] Возникающий в результате градиент давления выталкивает относительно прохладную воду с низкой соленостью из Атлантического океана через бассейн. Эта вода нагревается и становится более соленой по мере продвижения на восток, а затем опускается в районе озера. Левант и циркулирует на запад, чтобы разлиться обратно в Атлантику через Гибралтарский пролив.[15] Конечным результатом этого является то, что в Гибралтарском проливе имеется поверхностное течение на восток с холодной водой меньшей солености из Атлантики и одновременное течение на запад теплой соленой воды из Средиземного моря в более глубоких зонах. Вернувшись в Атлантику, этот химически отличный Средиземноморская промежуточная вода может сохраняться за тысячи километров от источника.[16]
- Солевые бассейны большие площади рассол на морское дно. Эти бассейны представляют собой водоемы, соленость которых в три-пять раз превышает соленость окружающего океана. Для глубоководных бассейнов с рассолом источником соли является растворение больших соль депозиты через соляная тектоника. Рассол часто содержит высокие концентрации метана, обеспечивая энергия к хемосинтетический экстремофилы которые живут в этом специализированном биом. Также известно, что бассейны с рассолом существуют на Антарктический континентальный шельф где источником рассола является соль, исключенная во время образования морской лед. Глубоководные бассейны с рассолом в Антарктике могут быть токсичными для морских животных. Бассейны с рассолом иногда называют озера на морском дне потому что плотный рассол создает галоклин которая плохо смешивается с вышележащей морской водой. Высокая соленость увеличивает плотность рассола, что создает четкую поверхность и береговую линию бассейна.[17]
В глубокое море, или глубокий слой, это самый нижний слой океана, существующий ниже термоклина на глубине 1000 саженей (1800 м) или более. Самая глубокая часть глубокого моря - это Марианская впадина расположен в западной части северной части Тихого океана. Это также самая глубокая точка земной коры. Он имеет максимальную глубину около 10,9 км, что превышает высоту гора Эверест. В 1960 г. Дон Уолш и Жак Пикар достигли дна Марианской впадины в Триест батискаф. Давление составляет около 11318 метрические тонны-сила за квадратный метр (110,99 МПа или 16100 psi ).
Зоны
Мезопелагический

Мезопелагическая зона - это верхняя часть средняя зона и простирается от 200 до 1000 метров (от 660 до 3280 футов) ниже уровень моря. Это в просторечии известно как «сумеречная зона», поскольку свет все еще может проникать в этот слой, но он слишком низкий, чтобы поддерживать фотосинтез. Однако ограниченное количество света все еще позволяет организмам видеть, а существа с чувствительным зрением могут обнаруживать добычу, общаться и ориентироваться с помощью своего зрения. У организмов в этом слое большие глаза, чтобы максимально использовать свет в окружающей среде.[18]
Большинство мезопелагических рыб производят ежедневно вертикальные миграции, перемещаясь ночью в эпипелагиали, часто следуя аналогичным миграциям зоопланктона, а днем возвращаясь в глубину в целях безопасности.[19][20]:585 Эти вертикальные миграции часто происходят на большие вертикальные расстояния и выполняются с помощью плавательный пузырь. Плавательный пузырь надувается, когда рыба хочет подняться, и, учитывая высокое давление в мезопелегической зоне, это требует значительных затрат энергии. По мере того, как рыба поднимается, давление в плавательном пузыре должно регулироваться, чтобы он не лопнул. Когда рыба хочет вернуться на глубину, плавательный пузырь сдувается.[21] Некоторые мезопелагические рыбы совершают ежедневные миграции через термоклин, где температура колеблется от 10 до 20 ° C (от 18 до 36 ° F), таким образом демонстрируя значительные допуски на изменение температуры.[20]:590
Мезопелагические рыбы обычно лишены защитных шипов и используют окраску и биолюминесценция к камуфляж их от других рыб. Засадные хищники темные, черные или красные. Поскольку более длинные красные волны света не достигают глубины моря, красный эффективно действует так же, как черный. Использование миграционных форм затушеванный серебристые цвета. На животе они часто показывают фотофоры производит низкокачественный свет. Для хищника снизу, смотрящего вверх, это биолюминесценция маскирует силуэт рыбы. Однако у некоторых из этих хищников есть желтые линзы, которые фильтруют (недостаточный красный) окружающий свет, оставляя биолюминесценцию видимой.[22]
Bathyal

Батильная зона - это нижняя часть средней водной зоны, охватывающая глубины от 1000 до 4000 метров (от 3300 до 13 100 футов). Свет не достигает этой зоны, что дало ей прозвище «полуночная зона»; из-за недостатка света он менее густонаселен, чем эпипелагиаль, хотя и намного больше.[23] Рыбам трудно жить в этой зоне, так как там давление давки, низкие температуры 4 ° C (39 ° F), низкий уровень растворенный кислород, а также недостаток питательных веществ.[20]:585 То немногое энергии, которое доступно в батипелагической зоне, фильтруется сверху в виде детрита, фекалий и случайных беспозвоночных или мезопелагических рыб.[20]:594 Около 20% пищи, происходящей из эпипелагиали, попадает в мезопелагиали, но только около 5% фильтруется в батипелагиали.[24] У рыб, которые там живут, возможно, уменьшились или полностью утрачены жабры, почки, сердце и плавательный пузырь, кожа стала слизистой, а не чешуйчатой, а скелет и мускулатура были слабыми.[20]:587 Большинство животных, обитающих в батильной зоне, - беспозвоночные, такие как морские губки, головоногие моллюски, и иглокожие. За исключением очень глубоких участков океана, батильная зона обычно достигает бентической зоны на морское дно.[23]
Абиссал и Хадаль

Абиссальная зона остается в постоянной темноте на глубине от 4000 до 6000 метров (от 13000 до 20 000 футов). Единственные организмы, населяющие эту зону: хемотрофы и хищников, которые могут противостоять огромному давлению, иногда достигающему 76 мегапаскалей (750 атм; 11 000 фунтов на квадратный дюйм). Хадальная зона (названа в честь Аид, то греческий бог подземного мира) - зона, предназначенная для самых глубоких траншеи в мире, достигая глубины менее 6000 метров (20 000 футов). Самая глубокая точка хадальной зоны - это Марианская впадина, который спускается на 10 911 метров (35 797 футов) и имеет давление 110 мегапаскалей (1100 атм; 16 000 фунтов на квадратный дюйм).[25][26][27]
Источники энергии

Морской снег
Верхняя фотическая зона океана заполнена частицами органического вещества (ПОМ) и весьма продуктивна, особенно в прибрежных районах и районах апвеллинга. Однако большая часть ПОМ имеет небольшие размеры и легкость. Этим частицам могут потребоваться сотни или даже тысячи лет, чтобы осесть через толщу воды в глубоком океане. Этой задержки достаточно для того, чтобы частицы реминерализовались и поглощались организмами в пищевых сетях.
Ученые из Океанографического института Вудс-Хоул провели эксперимент три десятилетия назад в глубоком Саргассовом море, изучая скорость опускания.[28] Они нашли то, что стало известно как морской снег в котором ПОМ переупаковывается в гораздо более крупные частицы, которые тонут с гораздо большей скоростью, «падая, как снег».
Из-за скудности пищи организмы, живущие на дне и на дне, обычно являются условно-патогенными. У них есть особые приспособления к этой экстремальной среде: быстрый рост, влияние механизма расселения личинок и способность использовать «временные» пищевые ресурсы. Типичный пример - растачивание по дереву. двустворчатые моллюски, которые проникают в древесину и другие остатки растений и питаются органическими веществами из остатков.
Кит падает
Для глубоководной экосистемы смерть КИТ это самое важное событие. Мертвый кит может унести на дно сотни тонн органического вещества. Кит падает Сообщество проходит три этапа:[29]
- Этап передвижного мусорщика: большие и подвижные глубоководные животные прибывают на место почти сразу после падения китов на дно. Амфиподы, крабы, спящие акулы и миксина все мусорщики.
- Оппортунистическая стадия: прибывают организмы, которые колонизируют кости и окружающие отложения, загрязненные органическими веществами туши и любой другой тканью, оставленной мусорщиками. Один род Оседакс,[30] трубчатый червь. Личинка рождается без пола. Окружающая среда определяет пол личинки. Когда личинка оседает на китовой кости, она превращается в самку; когда личинка поселяется на самке или в ней, она превращается в карликового самца. Одна самка Оседакс может нести более 200 таких особей мужского пола в своем яйцеводе.
- Сульфофильный Стадия: На этой стадии происходит дальнейшее разложение костей и восстановление сульфата морской воды. Бактерии создают богатую сульфидами среду, аналогичную гидротермальным источникам. Полиноиды, двустворчатые моллюски, брюхоногие моллюски и другие любящие серу существа переселяются.
Хемосинтез
Гидротермальные источники

Гидротермальные источники были обнаружены в 1977 году учеными из Института океанографии Скриппса. Пока все обнаруженные гидротермальные жерла расположены на границах плит: Восточная часть Тихого океана, Калифорния, Срединно-Атлантический хребет, Китай и Япония.
Новый материал океанического бассейна создается в таких регионах, как Срединно-Атлантический хребет, когда тектонические плиты отдаляются друг от друга. Скорость растекания пластин 1–5 см / год. Холодная морская вода течет вниз через трещины между двумя пластинами и нагревается, проходя через горячую породу. Минералы и сульфиды растворяются в воде при взаимодействии с горной породой. В конце концов, горячие растворы исходят из активного разлома на морском дне, создавая гидротермальный выход.
Хемосинтез бактерий обеспечивает энергией и органическими веществами всю пищевую сеть в вентиляционных экосистемах. Гигантские трубчатые черви может вырасти до 2,4 м (7 футов 10 дюймов) в высоту из-за обилия питательных веществ. В гидротермальных источниках обнаружено более 300 новых видов.[31]
Гидротермальные источники представляют собой целые экосистемы, независимые от солнечного света, и могут быть первым доказательством того, что Земля может поддерживать жизнь без солнца.
Холодные просачивания
А холодная утечка (иногда называемый холодным вентилем) - это область дно океана куда сероводород, метан и другие углеводород - происходит просачивание богатой жидкости, часто в виде бассейн с рассолом.
Экология
Глубокое море пищевые полотна сложны, а аспекты системы плохо изучены. Обычно взаимодействия хищника и жертвы в глубинах собираются путем прямого наблюдения (вероятно, из дистанционно управляемые подводные аппараты ), анализ содержимого желудка и биохимический анализ. Наиболее распространенный метод - анализ содержимого желудка, но для некоторых видов он ненадежен.[32]
В глубоководных пелагических экосистемах у побережья Калифорнии в трофической сети преобладают глубоководные рыбы, головоногие моллюски, студенистый зоопланктон, и ракообразные. В период с 1991 по 2016 год 242 уникальных кормовых взаимоотношения между 166 видами хищников и жертв продемонстрировали, что студенистый зоопланктон оказывает экологическое воздействие, подобное влиянию крупных рыб и кальмаров. Наркомедузы, сифонофоры (семьи Physonectae ), гребневики, а головоногие моллюски потребляли наибольшее разнообразие добычи в порядке убывания.[32] Каннибализм был зарегистрирован у кальмаров рода Гонат.[33]
Смотрите также
Рекомендации
- ^ а б Служба управления полезными ископаемыми Регион OCS Мексиканского залива (ноябрь 2006 г.). «Продажи OCS Oil and Gas в Мексиканском заливе: 2007–2012 годы. Продажи в западной зоне планирования 204, 207, 210, 215 и 218. Продажи в зоне центрального планирования 205, 206, 208, 213, 216 и 222. Проект воздействия на окружающую среду Заявление. Том I: главы 1–8 и приложения ». Министерство внутренних дел США, Служба управления полезными ископаемыми, Мексиканский залив, регион OCS, Новый Орлеан. стр. 3-27. PDF
- ^ «Подводная лодка-робот достигает глубочайшего океана». Новости BBC. 3 июня 2009 г.. Получено 2009-06-03.
- ^ а б Морской центр Гавайского университета (4 июня 2009 г.). «Ежедневные отчеты для НИС« КИЛО МОАНА »за июнь и июль 2009 г.». Гонолулу, Гавайи: Гавайский университет. Получено 24 июн 2010.
- ^ Морской центр Гавайского университета (4 июня 2009 г.). «Инвентаризация научного оборудования на НИС« КИЛО МОАНА »». Гонолулу, Гавайи: Гавайский университет. Получено 18 июн 2010.
- ^ а б К.Л. Смит-младший; Х.А. Руль; Б.Дж. Бетт; D.S.M. Биллетт; Р.С. Лэмпит; Р.С. Кауфманн (17 ноября 2009 г.). «Климат, круговорот углерода и глубоководные экосистемы». PNAS. 106 (46): 19211–19218. Bibcode:2009PNAS..10619211S. Дои:10.1073 / pnas.0908322106. ЧВК 2780780. PMID 19901326.
- ^ а б c d Хорхе Чирке (1997). «II. Пределы морской продуктивности» (PDF). В Эдварде А. Законе (ред.). Эль-Ниньо и перуанский промысел анчоусов (серия: Программа инструкций по глобальным изменениям). Обзоры в биологии рыб и рыболовстве. 9. Саусалито: Университетские научные книги. С. 118–121. Дои:10.1023 / А: 1008801515441. ISBN 978-0-935702-80-4. Получено 18 июн 2010.
- ^ а б Британская энциклопедия (2010). «Фотическая зона». Энциклопедия Britannica Online. Получено 18 июн 2010. Внешняя ссылка в
| publisher =(помощь) - ^ а б c d Жананда Кол (2004). «Зона Сумеречного океана (Дисфотическая)». EnchantedLearning.com. Получено 18 июн 2010.
- ^ а б c Кен О. Бюсселер; Карл Х. Ламборг; Филип В. Бойд; Фиби Дж. Лам; и другие. (27 апреля 2007 г.). «Возвращаясь к потоку углерода через сумеречную зону океана». Наука. 316 (5824): 567–570. Bibcode:2007Sci ... 316..567B. CiteSeerX 10.1.1.501.2668. Дои:10.1126 / science.1137959. PMID 17463282.
- ^ Национальное управление океанических и атмосферных исследований (2 декабря 2008 г.). "Насколько глубок океан?". Вашингтон, округ Колумбия: Национальное управление океанических и атмосферных исследований. Получено 19 июн 2010.
- ^ The Deep Sea на MarineBio.org - биология океана, морская жизнь, морские существа, охрана морской среды
- ^ Нюбаккен, Джеймс В. Морская биология: экологический подход. Пятое издание. Бенджамин Каммингс, 2001. стр. 136–141.
- ^ Пол Р. Пинет (1996). Приглашение к океанографии (3-е изд.). Сент-Пол, Миннесота: West Publishing Co., стр. 202. ISBN 978-0-314-06339-7.
- ^ Пинет 1996, стр. 206.
- ^ Пинет, 1996, стр. 206–207.
- ^ Пинет 1996, стр. 207.
- ^ NOAA исследование бассейна солевого раствора
- ^ «Средняя зона». Водная жизнь мира. 6. Тарритаун, Нью-Йорк: Marshall Cavendish Corporation. 2001. С. 340–341. ISBN 978-0-7614-7176-9.
- ^ Кость, Квентин; Мур, Ричард (2008). Биология рыб. Наука о гирляндах. п. 38. ISBN 978-0-203-88522-2.
- ^ а б c d е Moyle, P. B .; Чех, Дж. Дж. (2004). Рыбы, Введение в ихтиологию (5-е изд.). Бенджамин Каммингс. ISBN 978-0-13-100847-2.
- ^ Дуглас, Э .; Friedl, W .; Пиквелл, Г. (1976). «Рыбы в зонах минимума кислорода: характеристики оксигенации крови». Наука. 191 (4230): 957–9. Bibcode:1976Научный ... 191..957D. Дои:10.1126 / science.1251208. PMID 1251208.
- ^ Мунц, В. Р. А. (2009). «О желтых линзах у мезопелагических животных». Журнал Морской биологической ассоциации Соединенного Королевства. 56 (4): 963–976. Дои:10.1017 / S0025315400021019.
- ^ а б Эниг, К. С. (1997). Исследования морского бентоса. Испанский институт океанографии (на испанском языке). Мадрид: Министерство сельского хозяйства. С. 23–33. ISBN 978-84-491-0299-8.
- ^ Райан, Пэдди (21 сентября 2007 г.). «Глубоководные существа: Батипелагическая зона». Те Ара - Энциклопедия Новой Зеландии. Получено 4 сентября 2016.
- ^ "NOAA Ocean Explorer: История: цитаты: зондирование, морское дно и геофизика". NOAA, Управление океанических исследований и исследований. Получено 4 сентября 2016.
- ^ Смит, Крейг Р .; de Leo, Fabio C .; Бернардино, Анджело Ф .; Sweetman, Эндрю К .; Арбизу, Педро Мартинес (2008). «Глубинное ограничение пищи, структура экосистемы и изменение климата» (PDF). Тенденции в экологии и эволюции. 23 (9): 518–528. Дои:10.1016 / j.tree.2008.05.002. PMID 18584909.
- ^ Виноградова, Н. Г. (1997). «Зоогеография Абиссальной и Хадальской зон». Биогеография океанов. Успехи в морской биологии. 32. С. 325–387. Дои:10.1016 / S0065-2881 (08) 60019-X. ISBN 978-0-12-026132-1.
- ^ «Морской снег и фекальные гранулы».
- ^ Шана Гоффреди, Необычная донная фауна, связанная с падением кита в Монтерей-Каньон, Калифорния, Deep-Sea Research, 1295–1304, 2004
- ^ Ной К. Уайтмен, Между китовой костью и глубоким синим морем: происхождение карликовых самцов в трубчатых червях, питающихся китами, Молекулярная экология, 4395–4397, 2008
- ^ Ботос, Соня. «Жизнь на гидротермальном источнике».
- ^ а б Чой, К. Анела; Хэддок, Стивен Х.Д .; Робисон, Брюс Х. (2017-12-06). "Структура глубокой пелагической пищевой сети, выявленная на месте наблюдения за кормлением ". Proc. R. Soc. B. 284 (1868): 20172116. Дои:10.1098 / rspb.2017.2116. ЧВК 5740285. PMID 29212727.
- ^ Кляйн, Джоанна (19 декабря 2017 г.). "What Eats What: Путеводитель сухопутных жителей по глубоководной трапезе". Нью-Йорк Таймс. ISSN 0362-4331. В архиве с оригинала 20 декабря 2017 г.. Получено 2017-12-20.
дальнейшее чтение
- Куприянова Е.К., Винн О., Тейлор П.Д., Шопф Дж. У., Кудрявцев А. и Бейли-Брок, Дж. (2014). «Серпулиды, живущие глубоко: известковые трубчатые черви за бездной». Глубоководные исследования, часть I. 90: 91–104. Bibcode:2014DSRI ... 90 ... 91К. Дои:10.1016 / j.dsr.2014.04.006. Получено 2014-01-09.CS1 maint: несколько имен: список авторов (связь)

