Эхинодон - Echinodon
| Эхинодон | |
|---|---|
 | |
| Стоматология паратип образец NHMUK 48215b | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Заказ: | †Орнитишия |
| Семья: | †Heterodontosauridae |
| Род: | †Эхинодон Оуэн, 1861[1] |
| Разновидность: | †E. becklesii |
| Биномиальное имя | |
| †Эхинодон becklesii Оуэн, 1861 г.[1] | |
Эхинодон это род из гетеродонтозаврид динозавр которые жили в самые ранние Меловой южных Англия в Берриасский эпоха. Первыми образцами были кости челюсти, названные Эхинодон becklesii сэр Ричард Оуэн в 1861 году, и с момента их первоначального описания были обнаружены только дополнительные зубы. Специфическое имя награждает коллекционера Сэмюэл Беклс кто открыл материал Эхинодон и многие другие таксоны со всей Англии, в то время как название рода переводится как «колючий зуб» в отношении анатомии зубов таксона.
Первоначально Эхинодон считался типом травоядный ящерица, хотя ее быстро пересмотрели в орнитисхийцев среднего уровня. Относился к кладе Стегозаврия основанный на общей анатомии зубов и неправильно названной броне, которая позже была идентифицирована как черепаха с. Эхинодон затем был отнесен к раннему семейству орнитисхий Fabrosauridae, которая позже была идентифицирована как искусственная группа с Эхинодон переназначен на Heterodontosauridae. В то время как семья изначально считалась наиболее близкой к более производным Орнитоподы, в конечном итоге он был повторно идентифицирован как самая базальная группа орнитисхий, что Эхинодон таксон произошел от многих родов Раннеюрский период, с призрачной линией 50 миллионов лет несохраненной эволюции.
Все образцы Эхинодон были найдены в Purbeck Group из Дорсет, который по разным причинам относится к поздней юре или раннему меловому периоду. Текущие исследования принимают раннемеловой берриасский возраст, делая Эхинодон и самый молодой, и самый мелкий гетеродонтозаврид. Среди других динозавров, с которыми он жил, были орнитоподы. Овенодон и теропод Nuthetes, которые также фрагментарны. Рядом обитало множество мелких млекопитающих. Эхинодон, а осадки показывают, что пурбекская группа представляла собой лагунную среду с разной степенью разницы, первоначально похожую на современную Средиземноморье но со временем стал влажнее.
История открытия

Множественные образцы костей челюсти были обнаружены Сэмюэл Беклс высоко на скале в Durdlestone Bay на Остров Пурбек на юге Англия. Эти окаменелости, в том числе многие зубы а также верхняя челюсть и зубной кости верхней и нижней челюстей, были найдены рядом с раковинами и окаменелостями растений в Кровати Purbeck. Впервые они были описаны в монографии, опубликованной в 1861 написано сэром Ричард Оуэн, британский палеонтолог кто также описал окаменелости Игуанодон и Мегалозавр. Оуэн дал имя Эхинодон becklesii для окаменелостей, которые он считал частью клады ящериц Ласертилия.[1] В то время как конкретное имя удостоилось чести Беклза за его открытие окаменелостей и позволив Оуэну изучить его коллекцию окаменелостей Пурбека, родовое название произошло от Древнегреческий εχινος, «ёжик» и ὀδών, «зуб», который Оуэн объединил как «колючий зуб», чтобы описать анатомию зубцы по бокам зубов. Оуэн переписывался с британским палеонтологом. Хью Фалконер, который предложил название «Саураэчинодон», но в сокращенной форме Эхинодон не было озабоченный, Оуэн решил использовать сокращенную форму в качестве имени для своего нового животного.[1] В 1861 году Фальконер выпустил исправление, в котором указывалось, что он предложил название «Саураечмодон» вместо «Саураечинодон».[2]
Первоначально Оуэн описал его как лацертила, но он пересмотрел свою классификацию 1874 группировать Эхинодон с Скелидозавр и Игуанодон в кладе он назвал Прионодонтия, который был в большей кладе Динозаврия.[3] В 1888 Британский палеонтолог Ричард Лидеккер следовал предыдущей классификации Эхинодон как динозавр, основываясь на анатомии его зубов, описывая их как похожие на Скелидозавр хотя и не относя их к более специфической кладе, чем Dinosauria undeterminate. Серии экземпляров, обозначенных Оуэном как типы, были приобретены Британский музей естественной истории (BMNH теперь NHMUK) в 1876 и являются частью коллекции Beckles, имеют номера экземпляров NHMUK 48209–48215.[4] Лидеккер также уточнил, что Эхинодон был найден в слоях Среднего Пурбека,[4] неформальная единица современного Purbeck Limestone Group, официальное название исторических кроватей Purbeck.[5] Британский палеонтолог Питер Гальтон сузили место отложения Эхинодон дальше в 1978 в пресноводную «Грязь»,[6] также известная как «Яма млекопитающих», раскопанная Беклзом в 1857.[5] Однако нет никаких доказательств связи Эхинодон к любой конкретной кровати в Формация Лулворт группы Purbeck.[7] Гальтон также сослался на образец NHMUK 48229. Эхинодон, фрагментарный зубной камень с зубами,[6] и единственное дальнейшее отнесение к роду включает изолированные зубы также из пластов Purbeck.[8]

В конце 1970-х - начале 1980-х годов раскопки Музей естественной истории округа Лос-Анджелес обнаружил много мелких окаменелостей орнитисхий в Фруита Палеонтологический район г. Колорадо. Эти останки, собранные с песчаники в основе Brushy Basin Member из Формация Моррисон, были приблизительно 150,2-150,3 миллиона лет и первоначально были описаны как промежуточные останки фаброзавра их коллекционером Джорджем Каллистоном в 1984. Три года спустя Каллистон пересмотрел свое описание и сослался на материал, включая кости челюсти, позвонки и большую часть задних конечностей, чтобы Эхинодон sp., задание, поддержанное Гальтоном в 2002, хотя в 2006 он пересмотрел материал и заметил отличия в анатомии зубов от Эхинодон правильный. В 2009, этим окаменелостям был придан собственный род, Fruitadens haagarorum, родственный, но отличный от Эхинодон.[9]
Описание
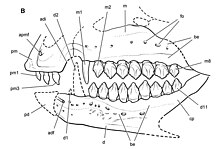
Известный материал Эхинодон ограничен костями черепа, но многочисленные образцы включают по крайней мере некоторые из предчелюстная кость, верхняя челюсть, слезный, скуловой, небный, эктоптеригоид и зубной вместе с большинством зубы верхней и нижней челюстей. На основании пропорций родственного рода Гетеродонтозавр, череп Эхинодон был бы 62 мм (2,4 дюйма) в длину, что сопоставимо с Тяньюлонг на 66 мм (2,6 дюйма), но меньше, чем у взрослых Fruitadens при длине 75 мм (3,0 дюйма), что делает Эхинодон самый маленький предположительно взрослый гетеродонтозавр и один из самые маленькие нептичьи динозавры.[7]
Основная часть обоих предчелюстных суставов сохраняется в лектотип образец Эхинодон, хотя и расколотый и раздавленный. Предчелюстной отверстие присутствует рядом с передний край отдельной кости и ямка он вложен в, больше похож по форме на Гетеродонтозавр чем более производный Гипсилофодон. Поверхность передней части кости вдоль зуба текстурированный и не имеет зубов, что отличает кость от кости Лесотозавр имеющий зубцы по всей длине. На предчелюстной кости было всего три зуба, без зубчиков (зубцов), присутствующих у большинства базальных орнитисхий, хотя все зубы примерно равны по размеру, причем третий был самым большим.[7]
Три максиллы сохранились среди материала Эхинодон, сохраняя почти всю кость, включая большинство областей контакта с другими костями черепа. Верхнечелюстные кости слегка уплощены, что сводит к минимуму силу щечной выемки, диагностического признака орнитисхиев, когда ряд верхнечелюстных зубов и кость непосредственно над ними вставляются от внешнего края кости. Наличие арочного диастема в Эхинодон это тема разногласий.[7] В то время как Гальтон и американский палеонтолог Пол Серено интерпретировать диастему как присутствующую Гетеродонтозавр,[6][10] Британские палеонтологи Дэвид Б. Норман и Пол М. Барретт в 2002 году пришел к выводу, что, основываясь на изображениях Оуэна до более позднего повреждения окаменелости NHMUK 48209, диастема отсутствовала.[8] Серено повторила в 2012 что диастема присутствовала и была изогнута на основе верхней челюсти NHMUK 48211.[7] На верхней челюсти имеется девять зубов. Эхинодон, первый из которых был увеличенным и тонким собачий аналогичны тем, которые наблюдаются в предчелюстной кости Lycorhinus и Гетеродонтозавр.[7] Оуэн также изобразил частичный зуб перед собачьей формой, но с тех пор он был утерян из-за повреждения материала. Хотя Норман и Барретт использовали это как доказательство наличия второй псовидной формы меньшего размера, чем та, что позади,[8] Гальтон в 1978 г. и Серено в 2012 г. идентифицировали только одну собаку.[6][7] Первый постклыковидный зуб Эхинодон является самым большим, хотя и ненамного выше, чем следующие, которые все того же размера. Вдоль середины коронок зубов имеется круглый выступ, но на коронке нет выступающих выступов. На каждой стороне коронки зуба по восемь-десять зубчиков.[7]
Слезные, скуловые и небные части почти не сохранились, хотя край орбита можно выявить в фрагменте слезной жидкости.[7] Эктоптеригоидная кость нёбо частично завершен и сохранился в сочленении с верхней челюстью NHMUK 48210. Отсутствие сохранности ограничивает анатомические детали, которые могут быть идентифицированы за пределами его сходства с костью, известной у других орнитизхов.[7]

В прецедентный кость не сохранилась в Эхинодон, но его наличие подтверждается морфологией переднего конца известных зубных костей. Как и у других гетеродонтозаврид, преддентарный при жизни лишь слабо артикулировался с дентарным, не имея боковой и вентральный процессы. У базального орнитиша зубная кость была глубокой, ее высота в середине составляла 30% от общей длины. Кость сужается кпереди по длине, хотя края под серединой зубного ряда субпараллельны. Ряд отверстие присутствуют по краю щечной выемки, как на верхней челюсти. В короноид В отличие от других базальных орнитисхов, отросток выражен, хотя контакт между зубными костями имеет V-образную форму, как у базальных орнитизхов.[7] В зубной части было 11 зубов, первые два из которых были специализированными по сравнению с большинством орнитизхов.[7] Хотя специальные передние зубы не обсуждались Гальтоном в 1978 году,[6] и считались отсутствующими Норманом и Барреттом в 2002 году,[8] Серено описал первые два альвеолы из Эхинодон как отличались по размеру от последующих зубов в 2012 году. Первая альвеола была чрезвычайно уменьшена, что указывает на небольшой штыревой первый зубной зуб, как на Lycorhinus, а второй был значительно увеличен, указывая на то, что клык больше, чем на верхней челюсти.[7] Обычные зубные зубы были немного выше, чем зубы верхней челюсти, и имели зубцы вдоль верхней половины коронки вместо верхних 25%. Как и в верхней челюсти, коронки имеют среднюю выпуклость, по восемь-десять зубцов по обе стороны от вершины и симметричны. эмаль.[7]
Классификация
Оуэн первоначально классифицировал Эхинодон как травоядная ящерица,[1] он изменил его размещение на один в Dinosauria.[3] Эхинодон считался промежуточным, потенциально орнитишическим динозавром, пока не был упомянут Франц Нопсато клады Scelidosaurinae в Стегозавры в 1928 на основе высокого венечного отростка и сужающихся зубов,[11] оба из которых считаются широко распространенными среди орнитисхийцев.[12] Изолированная кожная броня, найденная в пластах Пурбек, была отнесена к Эхинодон на основе классификации стегозавров Джастина Делэра в 1959,[13] хотя с тех пор они были переведены на солемидид черепахи.[12][14]

Классификации многих базальных орнитисхий были рассмотрены Ричард Талборн в 1971, где он разместил Эхинодон в Hypsilophodontidae: полученный из Триасовый формы без клыков (так называемые «фаброзавры»), близкие к юрским родам Лаозавр и Нанозавр, и примитивнее, чем меловые роды Гипсилофодон, Парксозавр и Тескелозавр.[15] В следующем году Гальтон опубликовал опровержение, не согласившись с тем, что Талборн использовал Hypsilophodontidae. Поскольку зубы в предчелюстной кости, одна из диагностических черт Талборна в семье, были примитивными, Гальтон утверждал, что они не должны использоваться для классификации орнитопод. Вместо, Фаброзавр и Эхинодон объединились в новую семью Fabrosauridae основан на боковом расположении зубов и удален из Hypsilophodontidae.[16] Гальтон продолжил эту классификацию, назвав род Лесотозавр в 1978 г., и вместе с его описанием рассмотрел анатомию нескольких базальных родов орнитисхий, включая Эхинодон, Нанозавр и Фаброзавр. Все но Эхинодон были окончательно отнесены им к Fabrosauridae, хотя размещение Эхинодон был сомнительным, поскольку он также имел сходство с кладой Heterodontosauridae, которые, как известно, также обладают увеличенными клыками. Однако Гальтон считал, что идентичность фаброзаврид более вероятна, поскольку на зубах этого таксона отсутствуют плоские фасетки износа.[6]
Серено была первой, кто реклассифицировал Эхинодон как член семейства Heterodontosauridae. В 1991, он пересмотрел черепную анатомию Лесотозаври сравнил его с большим количеством других базальных орнитисхий. Зубы Эхинодонна самом деле, были интерпретированы как наличие плоских фасеток износа на зубах, а также наличие зубцов, вставленных от края верхней челюсти, как у орнитисхов, более производных, чем Лесотозавр. Поскольку это было то, что было предложено для объединения рода с «фаброзаврами», Серено считал Эхинодон быть гетеродонтозавридом по наличию клыков и арочных промежутков в передней части зубного ряда.[17] Гетеродонтозавр, Абриктозавр и неописанная форма из Кайентская свита также были помещены в семью.[10] Норман и Барретт переописаны Эхинодон в 2002 г. и поддержал классификацию гетеродонтозаврид, но вместо этого отнес его к кладе на основании отсутствия отверстий на зубных костях и наличия зубцов, ограниченных верхней третью коронки.[12][8]
Хотя размещение в качестве гетеродонтозаврида для Эхинодон был подтвержден в дальнейших анализах, включая некоторые из первых филогенетических анализов орнитисхийцев, размещение самого семейства со временем изменилось.[18] Семья, либо исключая Эхинодон как в исследовании Тулборна 1971 года, или включая род, как в работе Гальтона и Серено, изначально считался группой базальных орнитоподы более производный, чем анкилозавры и стегозавры.[15][16][6][17][10] Дальнейшие исследования, включая обширный филогенетический анализ британского палеонтолога Ричард Дж. Батлер и коллеги в 2008 поддержали гетеродонтозаврид как наиболее базальных орнитисхийцев.[18] Пересмотренный анализ Батлера и другие. в 2011 также решено базальное размещение гетеродонтозаврид и решены внутренние взаимоотношения семейства с Эхинодон являясь базальным родом в семье вместе с Абриктозавр также.[19] После дополнительного анализа, проведенного южноамериканским палеонтологом Диего Полом и его коллегами в 2011 году, Эхинодон был решен как базальный орнитисхий, но не входящий в семейство Heterodontosauridae. Pol и другие. посчитал, что размещение за пределами Heterodontosauridae было связано не с тем, что он не принадлежал к семье, а по причине неполного характера его останков.[20]
В кладограмма Ниже следует анализ, проведенный Серено в 2012 году, включая правильно названные гетеродонтозавриды, описанные в то время. Кладу, включающую позднюю юру и ранний меловой период. Эхинодон, Fruitadens и Тяньюлонг плохо поддерживалась из-за небольшого количества зубов.[7]

В соответствии с резолюцией анализа 2012 года, проведенного Sereno, Эхинодон и другие лауразийские гетеродонтозавриды были исключены из клады гондванских гетеродонтозаврид. Эхинодон, Fruitadens, Тяньюлонг и у неописанных гетеродонтозаврид Кайента все зубы с низкой коронкой, в отличие от гондванских форм, а также дольчатые зубы. цингулум и заметная передняя бороздка, ведущая к отверстию на боковой поверхности зубной кости. Все, кроме гетеродонтозаврид Кайента, также значительно моложе остальных гетеродонтозаврид, и присутствие трех, объединенных в кладу, за исключением других таксонов, подразумевает неизвестную линию гетеродонтозаврид, которая существует 50 миллионов лет в юрском периоде. Отсутствуют данные для этого призрачное происхождение может быть из-за исключительно небольшого размера группы.[7]
Следуя более ранним результатам китайского палеонтолога Сюй Син и коллеги в 2006, где клады Гетеродонтозаврообразные был создан для объединения гетеродонтозавров, цератопсы и пахицефалозавры, Французский палеонтолог Поль-Эмиль Дьедонне и его коллеги предложили Эхинодон и другие гетеродонтозавры были ранними формами пахицефалозавров. Их результаты поместили Эхинодон как сестринский таксон таксонов, обычно классифицируемых в рамках Pachycephalosauria, за которым следует Тяньюлонг, в то время как все остальные изученные гетеродонтозавры сформировали группу у истоков клады. Эта гипотеза уменьшит призрачное происхождение пахицефалозавров и отодвинуть происхождение орнитопод до ранней юры.[21]
Палеобиология
Анатомия Эхинодон и другие гетеродонтозавриды малоизвестны; большинство палеобиологических аспектов основаны на почти полном роде Гетеродонтозавр. Носите фаски на коронках Гетеродонтозавр указывают на окклюзию в верхней части зуба, в отличие от предполагаемого движения челюсти в других формах, таких как Эхинодон где фаски износа есть только по бокам коронок. Несмотря на уникальное движение челюсти Гетеродонтозавр по сравнению с более примитивными гетеродонтозавридами функция увеличенных клыков, вероятно, была одинаковой для всей клады.[7] Основываясь на наличии износа на концах предчелюстной коронки и смещении зубного клыка относительно других зубов при закрытии челюсти, согласно Sereno в 2012 году, первичная функция зубного ряда предчелюстной кости будет заключаться в уборке растительности.[7] Это контрастирует с более ранними гипотезами Батлера. и другие. в 2008 г., что увеличенные клыки клады были всеядными.[18] Гипотеза о всеядности также была поддержана Норманом и его коллегами в 2011 году при интерпретации того, что клыки и предчелюстные зубы не изнашиваются от сельскохозяйственных культур.[22] В беззубый области предчелюстной кости и предчелюстной кости, вероятно, были основой ороговевших клювов, как и у других орнитисхий.[7]
Экология

Группа Purbeck представляет собой отличительную последовательность эвапориты, тонкий песчаники и шелли известняки перемежается с мергель и сланцы. Седиментология показывает, что они откладывались в течение пресная вода, солоноватый, гиперсоленый и квазиморской среды. Флора и фауна указывают на переменную земной, озерный, физиологический раствор и лагунный ассоциации. Климат ранней группы Пурбек, вероятно, был похож на современный Средиземноморье и стал более влажным к концу берриаса.[23] Хотя группа Purbeck изначально называлась неформальными слоями Purbeck, теперь ее можно разделить на верхние слои. Формация Дурлстон и нижний Формация Лулворт. «Верхние слои Пурбека» и большинство «срединных слоев Пербека» находятся в формации Дерлстон, самым старым отложением которой являются шлаковые слои в пачке ступенчатых отверстий.[5] Шлаковые пласты иногда считались границей юрского и мелового периодов, что привело бы к тому, что вся формация Лулворт была поздней юрской, Титонский, в возрасте.[6] Однако, несмотря на неопределенность в отношении возраста пластов из-за отсутствия корреляции по фауне или датировке, общепринято считать, что группа Пурбек полностью относится к самому раннему меловому периоду, а формация Лулворт является ранней. Берриасский.[6][23][24] В основе группы Purbeck явно лежит позднеюрский период. Portland Group в заливе Дурлстон и имеет переходную, но местами скрытую границу с вышележащими Wealden Group в Певерил Пойнт.[23]

Существует большая неуверенность относительно местонахождения образцов, собранных в формации Лулворт; единственный окончательный способ проверки - это анализ матрицы каждого образца для определения его солености.[5] Группа Purbeck имеет самую разнообразную орнитисхийскую фауну среди всех месторождений в Дорсет, и является одним из немногих берриасских отложений в мире, но почти полностью ограничивается черепным или дентальным материалом и следами. Owenodon hoggii - единственный другой орнитисхий, названный из пластов, и известен только по зубному камню с зубами, первоначально описанными как разновидность Игуанодон. Бедренная и дорсальная части промежуточного гадрозаврический также известен, наряду с промежуточными орнитопод и анкилозавры известен как по окаменелостям, так и по следам.[12] Помимо орнитишианов, формация Лулворт также содержит теропод. Nuthetes, амфибии, черепахи, ящерицы, змеи, млекопитающие и крокодилы, и разновидности беспозвоночных.[5][25][26] Амфибии из формации Лулворт включают саламандры Абрикосирен и промежуточный батрахозавроидид, то альбанерпетонтид Celtedens и лягушка Саннибатрахус.[25] Известно четыре таксона черепах: криптографы Дорсеточелис, Helochelydra, Hylaeochelys и Pleurosternon.[27][28] Пурбек - одно из самых разнообразных месторождений раннего мела во всем мире. лепидозавры.[29] Роды Беклезиус, Дорсетизавр, Дуротригия, Paramacellodus, Псевдозавриллы, Паразауриллы, Purbicella, Зауриллы, Парвираптор и три безымянных морфологии зубов представляют собой известные чешуйчатые,[30] и окаменелости, относящиеся к ринхоцефалы Гомеозавр и Опистии также были найдены.[31]
Разнообразный комплекс млекопитающих включает мелких евтерийцы Дурлстодон и Дурлстотерий;[23] неевтерианский Peramurans Перамус, Peramuroides, Магнимус и Куриогенис;[32][33] неевтерианский симродонты Спалакотерий, Тинодон и Тамодон;[34][35] неевтерианский дриолестоиды Ахиродон, Амблотерий, Дорсетодон, Чуннелодон и Фазолесты;[36][37][38] Затем на-евтерский многотуберкулезные Альбионбаатар, Болодон, Герхардодон, Плагиаулакс и Санниодон;[39][40][41] неевтерианский эвтриконодонты Триоракодон и Триконодон, Затем на-млекопитающее морганукодонтан Purbeckodon;[42] и не млекопитающие докодонт Перайоцинодон.[43] Крокодилы из залежей Лулворта включают Goniopholis gracilidens, Theriosuchus pusillus, Pholidosaurus purbeckensis, сомнительные останки, ранее известные как Goniopholis tenuidens,[5] и сомнительный таксон Macellodus brodiei.[1][30] Определенные участки внутри формации также сохраняют примитивные бекас летит Симулидиум и Псевдосимулиум,[26] и нематоцераны мухи Эоптихоптеры, Бродилка и Евкоретрина.[24]
Рекомендации
- ^ а б c d е ж Оуэн, Р. (1861). Монография по ископаемым рептилиям формаций Wealden и Purbeck. Часть V. Отряд Lacertilia. Палеонтографическое общество. С. 31–39.
- ^ Фальконер, Х. (1861 г.). "Примечание о синонимии ископаемого рода Эхинодон профессора Оуэна ". Летопись и журнал естественной истории. 8 (46): 341. Дои:10.1080/00222936108697425.
- ^ а б Оуэн, Р. (1874). Монография по ископаемым рептилиям формаций Wealden и Purbeck. Приложение № V. Динозаврия (Игуанодон). Палеонтографическое общество. С. 1–18.
- ^ а б Лидеккер, Р. (1888). Каталог ископаемых рептилий и амфибий в Британском музее (естественная история). Часть I. Содержит отряды Ornithosauria, Crocodilia, Dinosauria, Squamata, Rhynchocephalia и Pterosauria.. Тейлор и Фрэнсис. С. 247–248.
- ^ а б c d е ж Солсбери, С. (2002). «Крокодилы из нижнемеловой (берриасской) группы известняков Пурбек в Дорсете, южная Англия». In Milner, A.R .; Баттен, Д.Дж. (ред.). Жизнь и окружающая среда в Purbeck Times. Специальные статьи по палеонтологии. 68. С. 121–144.
- ^ а б c d е ж грамм час я Гальтон, П. (1978). «Fabrosauridae, базальное семейство орнитисхийских динозавров (Reptilia: Ornithopoda)». Paläontologische Zeitschrift. 52 (1–2): 138–159. Дои:10.1007 / BF03006735. S2CID 84613826.
- ^ а б c d е ж грамм час я j k л м п о п q р s т Серено, Пол С. (2012). «Таксономия, морфология, жевательная функция и филогения гетеродонтозавридных динозавров». ZooKeys (226): 1–225. Дои:10.3897 / zookeys.226.2840. ЧВК 3491919. PMID 23166462.
- ^ а б c d е Norman, D.B .; Барретт, П. (2002). «Орнитисхийские динозавры из нижнего мела (берриаса) Англии». In Milner, A.R .; Баттен, Д.Дж. (ред.). Жизнь и окружающая среда в Purbeck Times. Специальные статьи по палеонтологии. 68. С. 161–189.
- ^ Батлер, Р.Дж .; Гальтон, П.М.; Porro, L.B .; Chiappe, L.M .; Хендерсон, Д.М.; Эриксон, Г. (2009). «Нижние пределы размеров тела динозавров-орнитисхов, полученные из нового верхнеюрского гетеродонтозаврида из Северной Америки». Труды Королевского общества B. 277 (1680): 375–381. Дои:10.1098 / rspb.2009.1494. ЧВК 2842649. PMID 19846460.
- ^ а б c Серено, П. (1997). «Происхождение и эволюция динозавров». Ежегодный обзор наук о Земле и планетах. 25: 435–489. Дои:10.1146 / annurev.earth.25.1.435.
- ^ Нопса, Ф. (1928). «Роды рептилий». Palaeobiologica. 1: 163–188.
- ^ а б c d Barrett, P.M .; Maidment, S.C.R. (2011). «Динозавры Дорсета: Часть III, орнитисхийские динозавры (Dinosauria, Ornithischia) с дополнительными комментариями о зауроподах». Труды Дорсетского общества естествознания и археологии. 132: 145–163.
- ^ Делэр, Дж. Б. (1959). «Мезозойские рептилии Дорсета. Часть 2». Труды Дорсетского общества естествознания и археологии. 80: 52–90.
- ^ Barrett, P.M .; Clarke, J.B .; Brinkman, D.B .; Chapman, S.D .; Ensom, P.C. (2002). «Морфология, гистология и идентификация« граниконов »из известняковой формации Пурбек (нижний мел: берриас) в Дорсете, южная Англия». Меловые исследования. 23 (2): 279–295. Дои:10.1006 / cres.2002.1002.
- ^ а б Талборн, Р.А. (1971). «Происхождение и эволюция орнитисхийских динозавров». Природа. 234 (5324): 75–78. Дои:10.1038 / 234075a0. S2CID 4193262.
- ^ а б Гальтон, П. (1972). «Классификация и эволюция динозавров орнитопод». Природа. 239 (5373): 464–466. Дои:10.1038 / 239464a0. S2CID 4196759.
- ^ а б Серено, П. (1991). "Лесотозавр, «Фаброзавриды» и ранняя эволюция орнитиший ». Журнал палеонтологии позвоночных. 11 (2): 168–197. Дои:10.1080/02724634.1991.10011386.
- ^ а б c Батлер, Р.Дж .; Upchurch, P .; Норман, Д. (2008). «Филогения орнитисхийских динозавров». Журнал систематической палеонтологии. 6 (1): 1–40. Дои:10.1017 / S1477201907002271. S2CID 86728076.
- ^ Батлер, Р.Дж .; Liyong, J .; Jun, C .; Годфройт, П. (2011). "Посткраниальная остеология и филогенетическое положение небольшого орнитисхиального динозавра. Чангчунзавр парвус из формации Квантоу (мел: апт – сеноман) провинции Цзилинь, северо-восток Китая ». Палеонтология. 54 (3): 667–683. Дои:10.1111 / j.1475-4983.2011.01046.x.
- ^ Pol, D .; Rauhut, O.W.M .; Бесерра, М. (2011). «Среднеюрский динозавр гетеродонтозаврид из Патагонии и эволюция гетеродонтозаврид». Naturwissenschaften. 98 (5): 369–379. Дои:10.1007 / s00114-011-0780-5. PMID 21452054. S2CID 22636871.
- ^ Dieudonné, P. -E .; Cruzado-Caballero, P .; Godefroit, P .; Тортоса, Т. (2020). «Новая филогения цераподовых динозавров». Историческая биология: 1–21. Дои:10.1080/08912963.2020.1793979.
- ^ Norman, D.B .; Crompton, A.W .; Батлер, Р.Дж .; Porro, L.B .; Чариг, А.Дж. (2011). "Нижнеюрский орнитисхиальный динозавр Гетеродонтозавр tucki Кромптон и Чариг 1962: анатомия черепа, функциональная морфология, таксономия и взаимосвязи ». Зоологический журнал Линнеевского общества. 162: 182–279. Дои:10.1111 / j.1096-3642.2011.00697.x.
- ^ а б c d Sweetman, S.C .; Smith, G .; Мартил, Д. (2017). «Высоко производные эвтерианские млекопитающие из раннего мелового периода на юге Великобритании». Acta Palaeontologica Polonica. 62 (4): 657–665. Дои:10.4202 / app.00408.2017.
- ^ а б Лукашевич, Э.Д .; Coram, R.A .; Яржембовский, Э.А. (2001). «Новые настоящие мухи (Insecta: Diptera) из нижнего мела южной Англии». Меловые исследования. 22 (4): 451–460. Дои:10.1006 / cres.2001.0265.
- ^ а б Evans, S.E .; Макгоуэн, Г.Дж. (2002). «Остатки лиссамфибий из группы известняков Пурбек, южная Англия». In Milner, A.R .; Баттен, Д.Дж. (ред.). Жизнь и окружающая среда в Purbeck Times. Специальные статьи по палеонтологии. 68. С. 103–119.
- ^ а б Мостовский, М.Б .; Ross, A.J .; Szadziewski, R .; Кжеминский, В. (2003). "Повторное описание Simulidium priscum Вествуд и Псевдосимулиум хумидум (Brodie) (Insecta: Diptera: Rhagionidae) из группы известняков Пурбек (нижний мел) Англии ". Журнал систематической палеонтологии. 1 (1): 59–64. Дои:10.1017 / S1477201903001020. S2CID 55161519.
- ^ Милнер, А. (2004). "Черепахи Пурбекской известняковой группы Дорсета, южная Англия" (PDF). Палеонтология. 47 (6): 1441–1467. Дои:10.1111 / j.0031-0239.2004.00418.x.
- ^ Перес-Гарсия, А. (2014). "Доработка малоизвестных Типокард дорсетохелиса, относительно многочисленная плевростернидная черепаха (Paracryptodira) в раннем меловом периоде Европы ». Меловые исследования. 49: 152–162. Дои:10.1016 / j.cretres.2014.02.015.
- ^ Sweetman, S.C .; Эванс, С. (2011). «Лепидозавры (Ящерицы)». В Баттен, Д. (ред.). Английские окаменелости Wealden. Палеонтологическая ассоциация. С. 264–284. ISBN 978-1-444-36711-9.
- ^ а б Evans, S.E .; Jones, M.E.H .; Мацумото, Р. (2012). «Новый череп ящерицы из группы известняков Пурбек (нижний мел) Англии». Bulletin de la Société Géologique de France. 183 (6): 517–524. Дои:10.2113 / gssgfbull.183.6.517.
- ^ Эванс, С. (1998). «Фауны лепидозавров из раннего мела: клады в переходный период». В Lucas, S.G .; Kirkland, J.I .; Эстеп, Дж. (ред.). Наземные экосистемы нижнего и среднего мела. Музей естественной истории Нью-Мексико Бюллетень 14. С. 195–200.
- ^ Дэвис, Б. (2012). «Микрокомпьютерная томография выявляет разнообразие перамуранских млекопитающих из группы Пурбек (берриас) в Англии». Палеонтология. 55 (4): 789–817. Дои:10.1111 / j.1475-4983.2012.01161.x.
- ^ Сигоньо-Рассел, Д. (1999). "Переоценка Перамуры (Млекопитающие, Кладотерия) на основе новых характеристик Crétacé inférieur d'Angleterre et du Maroc". Геодиверситас. 21 (1): 93–127.
- ^ Ensom, P.C .; Сигоньо-Рассел, Д. (2000). «Новые симродонты (Mammalia, Theria) из группы известняков Пурбек, нижний мел, юг Англии». Меловые исследования. 21 (6): 767–779. Дои:10.1006 / cres.2000.0227.
- ^ Sigogneau-Russell, D .; Ensom, P.C. (1998). "Тамодон (Theria, Symmetrodonta) из нижнего мела Северной Африки и Европы и краткий обзор симмродонтов ». Меловые исследования. 19 (3–4): 445–470. Дои:10.1006 / cres.1998.0115.
- ^ Ensom, P.C .; Сигоньо-Рассел, Д. (1998). «Новые дриолестоидные млекопитающие из группы известняков Пурбек базального мела на юге Англии». Палеонтология. 41 (1): 35–55.
- ^ Оуэн, Р. (1871). "Монография ископаемых млекопитающих мезозойских формаций". Палеонтографическое общество: 1–115.
- ^ Аверьянов, А.О .; Martin, T .; Лопатин, А. (2013). "Новая филогения базальных Trechnotheria и Cladotheria и родство южноамериканских эндемичных позднемеловых млекопитающих". Naturwissenschaften. 100 (4): 311–326. Дои:10.1007 / s00114-013-1028-3. PMID 23494201. S2CID 18504005.
- ^ Kielan-Jaworowska, Z .; Ensom, P.C. (1994). «Крошечные плагиаулакоидные многотуберкулезные млекопитающие из известняковой формации Пурбек, Дорсет, Англия». Палеонтология. 37 (1): 17–31.
- ^ Kielan-Jaworowska, Z .; Ensom, P.C. (1994). «Многотуберкулезные млекопитающие из верхнеюрской формации известняков Пурбек на юге Англии». Палеонтология. 35 (1): 95–126.
- ^ Фальконер, Х. (1857 г.). "Описание двух видов ископаемых млекопитающих. Плагиаулакс от Purbeck ». Ежеквартальный журнал геологического общества. 13 (1–2): 261–282. Дои:10.1144 / GSL.JGS.1857.013.01-02.39. S2CID 128645686.
- ^ Батлер, П.М.; Sigogneau-Russell, D .; Ensom, P.C. (2012). «Возможное сохранение morganucodontans в группе известняков Purbeck нижнего мела (Дорсет, Англия)». Меловые исследования. 33 (1): 135–145. Дои:10.1016 / j.cretres.2011.09.007.
- ^ Сигоньо-Рассел, Д. (2003). «Докодонты из британского мезозоя». Acta Palaeontologica Polonica. 48 (3): 357–374.
| ||||||||||||||||||||||||||
| ||||||||||||||||||||||||||
