Гаплогруппа L2 (мтДНК) - Haplogroup L2 (mtDNA)
| Гаплогруппа L2 | |
|---|---|
| Возможное время происхождения | 80 000–111 100 YBP[1] |
| Возможное место происхождения | Западная Африка[2] или же Центральная Африка |
| Предок | L2─6 |
| Потомки | L2a─d, L2e |
| Определение мутаций | 146, 150, 152, 2416, 8206, 9221, 10115, 13590, 16311!, 16390[3] |
Гаплогруппа L2 это митохондриальная ДНК человека (мтДНК) гаплогруппа с широко распространенным современным распространением, особенно в Приэкваториальной Африке. Его субклад L2a является довольно частым и широко распространенным кластером мтДНК на континенте, а также среди афроамериканцев.
Источник
L2 - распространенная линия в Африке. Считается, что он появился от 87 000 до 107 000 лет назад.[4] или ок. 90,000 YBP.[1] Его возраст, широкое распространение и разнообразие по всему континенту затрудняют определение точного места его происхождения в Африке.[5] Несколько гаплотипов L2 наблюдались у гвинейцев и других Западная Африка популяции имели общие генетические совпадения с Восточная Африка и Северная Африка.[6] Кажется вероятным происхождение L2b, L2c, L2d и L2e в Западной или Центральной Африке.[5] Раннее разнообразие L2 можно наблюдать на всем Африканском континенте, но, как мы видим в разделе Субклады ниже, наибольшее разнообразие наблюдается в Западная Африка. Большинство субкладов в основном приурочены к Западной и Западной Центральной Африке.[7]
Согласно исследованию 2015 года, «результаты показывают, что линии в южной части Африки объединяются с линиями Западной / Центральной Африки в недавнем временном масштабе, тогда как восточные линии кажутся значительно более древними. Три момента расширения из центральноафриканского источника связаны с L2: одна миграция 70–50 тыс. Лет назад в Восточную или Южную Африку, послеледниковые перемещения 15–10 тыс. Лет назад в Восточную Африку; и экспансия банту на юг за последние 5 тыс. Лет назад. Анализ комплементарной популяции и филогеографии L0a не указывает убедительных доказательств наличия гена мтДНК. переход между восточными и южными популяциями во время более позднего перемещения, что свидетельствует о низкой степени смешения восточноафриканского населения и мигрантов банту. Это означает, что, по крайней мере на ранних этапах, распространение банту было в основном демическим рассредоточением с незначительным включением местного населения ".[8]
Распределение
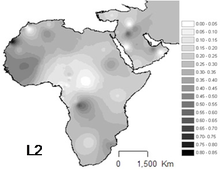
L2 - самая распространенная гаплогруппа в Африке, и она наблюдалась по всему континенту. Он встречается примерно у одной трети африканцев и их недавних потомков.
Самая высокая частота встречается среди Пигмеи мбути (64%).[9] Важное присутствие в Западная африка, особенно в Сенегал (43-54%).[6] Также важен для популяций не банту Восточная Африка (44%),[10] в Судан и Мозамбик.
Особенно много в Чад и Канембу (38% выборки), но также относительно часто встречается в Кочевой Арабов (33%) [Cerny et al. 2007][5] и Народ акан (~33%)[11]
Субклады
|
L2 имеет пять основных подгаплогрупп: L2a, L2b, L2c, L2d и L2e. Из этих линий наиболее распространенным субкладом является L2a, который встречается как в Африке, так и в Левант.
Гаплогруппа L2 обнаружена среди особей на островном кладбище в г. Кулубнарти, Судан, которые датируются Ранний христианин период (550–800 гг. нашей эры).[12]
Гаплогруппа L2a
L2a широко распространен в Африка и наиболее распространенная и широко распространенная африканская гаплогруппа к югу от Сахары, а также довольно часто встречается на уровне 19% в Америка среди потомков африканцев (Salas et al., 2002). L2a имеет возможную дату происхождения ок. 48000 лир.[1]
Особенно много в Чад (38% выборки; 33% недифференцированных L2 среди Чадские арабы,[13]), а в популяциях небанту Восточная Африка (Кения, Уганда и Танзания ) на уровне 38%.[10] Около 33% в Мозамбик[14] и 32% в Гана.[11]
Этот субклад характеризуется мутациями в 2789, 7175, 7274, 7771, 11914, 13803, 14566 и 16294. Он составляет 52% от общего L2 и является единственным субкладом L2, широко распространенным по всей Африке.[15]
Широкое распространение L2a и разнообразие затрудняют определение географического происхождения. Основная загадка - это почти повсеместная гаплогруппа L2a, которая, возможно, распространилась на восток и запад вдоль Сахель Коридор в Северная Африка после Последний ледниковый максимум, или истоки этих расширений могут лежать раньше, в начале позднего каменного века ∼ 40 000 лет назад.[5][15]
В Восточная Африка L2a был обнаружен 15% в Долина Нила –Нубия, 5% от Египтяне, 14% Кушит спикеры, 15% Семитский Люди амхара, 10% от Gurage, 6% Тыграй-тигринский народ, 13% Эфиопы и 5% Йеменцы.[14]
Гаплогруппа L2a также появляется в Северная Африка, с максимальной частотой 20% Туареги, Фулани (14%). Найдено также среди некоторых Алжир Арабов, он встречается у 10% среди Марокканский Арабов, некоторые марокканские берберы и Тунисский Берберы. (Watson 1997) и др. (бдительный 1991) и др. 1991 г.
У пациентов, которым вводят препарат ставудин лечить ВИЧ, Гаплогруппа L2a связана с меньшей вероятностью периферическая невропатия как побочный эффект.[16]
Гаплогруппа L2a1
L2a может быть далее разделен на L2a1, укрывающий переход на 16309 (Salas et al. 2002).
Этот субклад с разной частотой наблюдается в Западной Африке среди Малинке, Волоф, и другие; среди Североафриканцы;в Сахель среди Хауса, Фульбе, и другие; в Центральная Африка среди Бамилеке, Фали и другие; в Южная Африка среди Хойсан семья, включая Кхве и банту компьютерные колонки; И в Восточная Африка среди Кикую из Кения.
Все клады L2 присутствуют в Эфиопия в основном происходят от двух субкладов, L2a1 и L2b. L2a1 определяется мутациями в 12693, 15784 и 16309. Большинство эфиопских последовательностей L2a1 имеют общие мутации в nps 16189 и 16309. Однако, в то время как большинство (26 из 33) афро-американцы Общие полные последовательности L2a гаплогруппы могут быть разделены на четыре субклада с помощью замен в nps L2a1e-3495, L2a1a-3918, L2a1f-5581 и L2a1i-15229. Ни одна из этих последовательностей не наблюдалась в образцах 16309 L2a1 Эфиопии. (Салас 2002) и др.
Гаплогруппа L2a1 также наблюдалась среди Махра (4.6%).[17]
Гаплогруппа L2a1 была обнаружена в древних окаменелостях, связанных с Докерамический неолит культура в Скажи Халуле, Сирия.[18] Образец, раскопанный на Саванна Пастораль эпохи неолита сайт Люксманда в Танзания также несли кладу L2a1. Кластерный анализ примесей далее показал, что индивид имел значительную родословную из древнего Леванта, подтверждая родовые связи между создателями пасторального неолита саванны и докерамического неолита.[19]
Гаплогруппа L2a1a
Субклад L2a1a определяется заменами в 3918, 5285, 15244 и 15629. Есть два кластера L2a, которые хорошо представлены у юго-восточных африканцев, L2a1a и L2a1b, оба определяются переходами в довольно стабильных положениях HVS-I. Оба они, по-видимому, происходят из Западной Африки или Северо-Западная Африка (на что указывает распределение совпадающих или соседних типов) и претерпели резкое расширение либо в Юго-Восточной Африке, либо в популяции, являющейся предком современных жителей Юго-Восточной Африки.
Самые недавние вспышки звездообразования в субкладах L2a1a и L2a2 предполагают подпись для расширений банту, как также было предложено Перейрой и др. (2001).
L2a1a определяется мутацией в 16286. Кандидат в основатели L2a1a датируется 2700 (1200 SE) лет назад. (Перейра и др., 2001). Однако L2a1a, как определено заменой в (np 16286) (Salas et al. 2002), теперь поддерживается маркером кодирующей области (np 3918) (рис. 2A) и обнаружен в четырех из шести Йеменцы L2a1 линии. L2a1a встречается с самой высокой частотой в Юго-Восточной Африке (Pereira et al. 2001; Salas et al. 2002). Как частый гаплотип-основатель, так и производные линии (с мутацией 16092), обнаруженные среди Йеменцы иметь точные совпадения в Мозамбик последовательности (Перейра и др., 2001; Салас и др., 2002). L2a1a также встречается с меньшей частотой в Северо-Западной Африке, среди Мор и Бамбара из Мали и Мавритания.[20] (Рандо и другие. 1998; Мака-Мейер и другие. 2003)
Гаплогруппа L2a1a1
L2a1a1 определяется маркерами 6152C, 15391T, 16368C
Гаплогруппа L2a1b
L2a1b определяется заменами в 16189 и 10143. 16192 также часто встречается в L2a1b и L2a1c; он появляется в Северной Африке в Египте, он также появляется в Юго-Восточной Африке и поэтому он также может быть маркером для Расширение банту.[5]
Гаплогруппа L2a1c
L2a1c часто разделяет мутацию 16189 с L2a1b, но имеет свои собственные маркеры на 3010 и 6663. 16192 также часто встречается в L2a1b и L2a1c; он появляется в Юго-Восточной Африке, а также в Восточной Африке.[21] Это предполагает некоторую диверсификацию этой клады in situ.
Позиции T16209C C16301T C16354T на вершине L2a1 определяют небольшую подкладу, названную L2a1c Kivisild et al. (2004, рисунок 3) (см. Также рисунок 6 в Salas et al. 2002), который в основном появляется в Восточная Африка (например. Судан, Нубия, Эфиопия ), среди Туркана и Западная Африка (например. Канури ).
в Бассейн Чада, были идентифицированы четыре различных типа L2a1c с одной или двумя мутационными стадиями из типов Восточной и Западной Африки. (Кивисилд и др.) 2004 г.[21] (цитата на странице 9 или 443)[22][23]
Гаплогруппа L2a1c1
L2a1c1 имеет североафриканское происхождение.[24] Он определяется маркерами 198, 930, 3308, 8604, 16086. Он наблюдается у сефардов, ашкеназов, евреев, евреев, марокканцев, египтян, нубийцев и йеменцев Туниса.
Гаплогруппа L2a1f
Хосиан, Замбия, Мадагаскар
Гаплогруппа L2a1k
L2a1k определяется маркерами G6722A и T12903C. Ранее он был описан как специфичный для Европы субклад L2a1a и обнаружен в Чехи и Словаки.[25]
Гаплогруппа L2a1l2a
L2a1l2a распознается как "Ашкенази -специфическая гаплогруппа, наблюдаемая среди евреев-ашкенази с родословной в Центральной и Восточной Европе. Она также была обнаружена в небольшом количестве в якобы нееврейских польских популяциях, где предположительно произошла от примеси ашкенази.[26] Однако этот гаплотип составляет лишь очень небольшую часть митохондриальных линий ашкенази; различные исследования (включая исследование Бехара) показали, что заболеваемость составляет 1,4–1,6%.
Гаплогруппа L2a2
L2a2 характерен для Пигмеи мбути.[9]
Гаплогруппа L2b'c
L2b'c, вероятно, возник около 62 000 лет назад.[1]
Гаплогруппа L2b
Этот субклад преимущественно встречается в Западная Африка, но он распространен по всей Африке.[27]
Гаплогруппа L2c
L2c наиболее часто встречается в Западной Африке и, возможно, возник там.[15] Специально присутствует в Сенегал на 39%, Кабо-Верде 16% и Гвинея-Бисау 16%.[6]
Гаплогруппа L2d
L2d чаще всего встречается в Западной Африке, где он мог возникнуть.[15] Он также встречается в Йемене, Мозамбике и Судане.[14]
Гаплогруппа L2e
L2e (бывшее L2d2) типично в Западная Африка.[5] Он также встречается в Тунис,[28] и среди Народ мандинка из Гвинея-Бисау и афроамериканцы.[27]
Дерево

Это филогенетическое дерево субкладов гаплогруппы L2 основано на статье Манниса ван Овена и Манфреда Кайзера. Обновленное комплексное филогенетическое дерево глобальных вариаций митохондриальной ДНК человека[3] и последующие опубликованные исследования.
- Самый недавний общий предок (MRCA)
- L1'2'3'4'5'6
- L2'3'4'6
- L2
- L2a'b'c'd
- L2a
- L2a1
- L2a1a
- L2a1a1
- L2a1a2
- L2a1a2a
- L2a1a2a1
- L2a1a2b
- L2a1a2a
- L2a1a3
- 16189 (16192)
- L2a1b
- L2a1b1
- L2a1f
- L2a1f1
- L2a1b
- 143
- L2a1c
- L2a1c1
- L2a1c2
- L2a1c3
- L2a1c4
- L2a1d
- L2a1e
- L2a1e1
- L2a1h
- 16189
- L2a1i
- L2a1j
- L2a1k
- 16192
- L2a1l
- L2a1l1
- L2a1l1a
- L2a1l2
- L2a1l1
- L2a1l
- L2a1c
- L2a1a
- L2a2
- L2a2a
- L2a2a1
- L2a2b
- L2a2b1
- L2a2a
- L2a1
- L2b'c
- L2b
- L2b1
- L2b1a
- L2b1a2
- L2b1a3
- L2b1a
- L2b1
- L2c
- L2c2
- L2c2a
- L2c3
- L2c2
- L2b
- L2d
- L2d1
- L2d1a
- L2d1
- L2a
- L2e
- L2a'b'c'd
- L2
- L2'3'4'6
- L1'2'3'4'5'6
Смотрите также
- Генеалогический ДНК-тест
- Генетическая генеалогия
- Митохондриальная генетика человека
- Популяционная генетика
- Гаплогруппы митохондриальной ДНК человека
Филогенетическое дерево гаплогруппы митохондриальной ДНК человека (мтДНК) | |||||||||||||||||||||||||||||||||||||||
| Митохондриальная Ева (L ) | |||||||||||||||||||||||||||||||||||||||
| L0 | L1–6 | ||||||||||||||||||||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | L6 | ||||||||||||||||||||||||||||||||||
| M | N | ||||||||||||||||||||||||||||||||||||||
| CZ | D | E | грамм | Q | О | А | S | р | я | W | Икс | Y | |||||||||||||||||||||||||||
| C | Z | B | F | R0 | pre-JT | п | U | ||||||||||||||||||||||||||||||||
| HV | JT | K | |||||||||||||||||||||||||||||||||||||
| ЧАС | V | J | Т | ||||||||||||||||||||||||||||||||||||
Рекомендации
- ^ а б c d Соарес, Педро; Лука Эрмини; Ноэль Томсон; Мару Мормина; Тереза Рито; Арне Рёль; Антонио Салас; Стивен Оппенгеймер; Винсент Маколей; Мартин Б. Ричардс (4 июня 2009 г.). «Поправка на очищающий отбор: улучшенные митохондриальные молекулярные часы человека». Американский журнал генетики человека. 84 (6): 82–93. Дои:10.1016 / j.ajhg.2009.05.001. ЧВК 2694979. PMID 19500773. Получено 2009-08-13.
- ^ Сильва, Марина (2015). «60 000 лет взаимодействий между Центральной и Восточной Африкой, документально подтвержденных основной африканской митохондриальной гаплогруппой L2». Научные отчеты. Природа. 5: 12526. Bibcode:2015НатСР ... 512526С. Дои:10.1038 / srep12526. ЧВК 4515592. PMID 26211407.
- ^ а б ван Овен, Маннис; Манфред Кайзер (13 октября 2008 г.). «Обновленное комплексное филогенетическое дерево глобальных вариаций митохондриальной ДНК человека». Человеческая мутация. 30 (2): E386 – E394. Дои:10.1002 / humu.20921. PMID 18853457. S2CID 27566749. Архивировано из оригинал 4 декабря 2012 г.. Получено 2009-05-20.
- ^ Тишкофф и др., Анализ последовательности генома цельной мтДНК древних африканских линий, Молекулярная биология и эволюция, т. 24, вып. 3 (2007), стр. 757–768.
- ^ а б c d е ж Салас, Антонио и др., Создание африканского ландшафта мтДНК, Американский журнал генетики человека, т. 71, нет. 5 (2002), стр. 1082–1111.
- ^ а б c Роза, Александра; Брем, А; Кивисилд, Т; Мецпалу, Э; Виллемс, Р. и другие. (2004). «Профиль мтДНК жителей Западной Африки в Гвинее: на пути к лучшему пониманию региона Сенегамбия». Анналы генетики человека. 68 (Pt 4): 340–352. Дои:10.1046 / j.1529-8817.2004.00100.x. PMID 15225159. S2CID 15391342.
- ^ Атлас путешествия человека: гаплогруппа L2 В архиве 2011-10-06 на Wayback Machine Генографический проект, National Geographic.
- ^ Сильва, Марина; Альшамали, Фарида; Сильва, Паула; Каррильо, Карла; Мандлат, Флавио; Хесус Тровоада, Мария; Черный Виктор; Перейра, Луиза; Соарес, Педро (2015). «60 000 лет взаимодействий между Центральной и Восточной Африкой, документально подтвержденных основной африканской митохондриальной гаплогруппой L2». Научные отчеты. 5: 12526. Bibcode:2015НатСР ... 512526С. Дои:10.1038 / srep12526. ЧВК 4515592. PMID 26211407.
- ^ а б Quintana-Murci et al. 2008 г. Материнские следы глубокого общего предка и асимметричный поток генов между охотниками-собирателями пигмеев и фермерами, говорящими на банту «Труды Национальной академии наук Соединенных Штатов Америки». 105 (5): 1599
- ^ а б Сэди Андерсон-Манн 2006, Филогенетический и филогеографический анализ изменчивости африканской митохондриальной ДНК. В архиве 2011-09-10 на Wayback Machine
- ^ а б Вирама, Кришна Р и др. 2010 г., Небольшая генетическая дифференциация, оцененная по однопородным маркерам, при наличии значительных языковых вариаций у народов региона Кросс-Ривер в Нигерии.
- ^ Сирак, Кендра; Френандес, Даниэль; Новак, Марио; Ван Гервен, Деннис; Пинхаси, Рон (2016). «Тезисы докладов Межконгресса IUAES 2016 - Сообщество разделено? Выявление генома (ов) сообщества средневековой Кулубнарти с использованием секвенирования следующего поколения». Тезисы докладов Межконгресса Iuaes 2016. IUAES: 115.
- ^ Сересо, Мария; и другие. (2011). «Новое понимание структуры населения бассейна озера Чад, выявленное с помощью высокопроизводительного генотипирования митохондриальной ДНК, кодирующей SNP». PLOS ONE. 6 (4): e18682. Bibcode:2011PLoSO ... 618682C. CiteSeerX 10.1.1.291.8871. Дои:10.1371 / journal.pone.0018682. ЧВК 3080428. PMID 21533064.
- ^ а б c Тоомас Кивисилд и др., Эфиопское наследие митохондрий ДНК: отслеживание потока генов через и вокруг ворот слез, Американский журнал генетики человека, т. 75, нет. 5 (ноябрь 2004 г.), стр. 752–770.
- ^ а б c d Антонио Торрони и др., Развиваются ли четыре клады гаплогруппы L2 мтДНК с разной скоростью ?, Американский журнал генетики человека, т. 69 (2001), стр. 348–1356.
- ^ Кампира, Э; Kumwenda, J; ван Остерхаут, JJ; Дандара, К. (август 2013 г.). «Субгаплогруппы L0a2 и L2a митохондриальной ДНК изменяют предрасположенность к периферической невропатии у взрослых малавийских жителей, получающих ставудин, содержащий высокоактивную антиретровирусную терапию». Синдр иммунодефицита J Acquir. 63 (5): 647–652. Дои:10.1097 / QAI.0b013e3182968ea5. ЧВК 3815091. PMID 23614993.
- ^ Нет, Эми. «АНАЛИЗ ГЕНЕТИЧЕСКИХ ДАННЫХ В МЕЖДИСЦИПЛИНАРНОЙ ОСНОВЕ ДЛЯ ИССЛЕДОВАНИЯ ПОСЛЕДНИЙ ЭВОЛЮЦИОННОЙ ИСТОРИИ ЧЕЛОВЕКА И КОМПЛЕКСНЫХ ЗАБОЛЕВАНИЙ» (PDF). Университет Флориды. Получено 2 ноября 2016.
- ^ Манко, Жан (2013). Путешествие к предкам: заселение Европы от первых авантюристов до викингов. Темза и Гудзон. п. 88. ISBN 978-0500771822. Получено 29 сентября 2017.
- ^ Скоглунд; и другие. (21 сентября 2017 г.). «Реконструкция доисторической структуры населения Африки». Клетка. 171 (1): 59–71. Дои:10.1016 / j.cell.2017.08.049. ЧВК 5679310. PMID 28938123. Получено 15 октября 2017.
- ^ Гонсалес, А. М. и др., 2006 г., Вариации митохондриальной ДНК в Мавритании и Мали и их генетическая связь с другими популяциями Западной Африки
- ^ а б "Интернет-библиотека Wiley | Научно-исследовательские статьи, журналы, книги и справочные материалы". Архивировано из оригинал на 2010-08-05. Получено 2009-05-19.
- ^ Черни, В. и др., 2006 г., Двунаправленный коридор в поясе Сахель-Судан и отличительные черты популяций бассейна Чада: история, выявленная геномом митохондриальной ДНК.
- ^ Ласкаро, Даниэла; кастельхана, Стефано; Гаспар, Джузеппе; Ромео, Джованни; Сакконе, Сесилия; Аттимонелли, Марселла (2008). «Сборник RHNumtS: особенности и биоинформатические подходы к поиску и количественной оценке Human NumtS». BMC Genomics. 9: 267. Дои:10.1186/1471-2164-9-267. ЧВК 2447851. PMID 18522722. Получено 12 апреля 2020.
- ^ Ласкаро, Даниэла; Кастеллана, Стефано; Гаспар, Джузеппе; Ромео, Джованни; Сакконе, Сесилия; Аттимонелли, Марселла (2008). «Сборник RHNumtS: особенности и биоинформатические подходы к поиску и количественной оценке Human NumtS». BMC Genomics. 9: 267. Дои:10.1186/1471-2164-9-267. ЧВК 2447851. PMID 18522722.
- ^ Борис Малярчук, Мирослава Деренко, Мария Перкова, Томаш Гржибовски, Томаш Ванчек и Ян Лазур, Реконструкция филогении линий митохондриальной ДНК африканских народов у славян, Европейский журнал генетики человека, т. 16 (2008), стр. 1091–1096.
- ^ Марта Мельник-Сикорска, Патрица Дака, Борис Малярчук, Мирослава Деренко, Катаржина Сконечна, Мария Перкова, Тадеуш Добош, Томаш Гжибовски, История славян на основании полных последовательностей митохондриального генома PLOSOne 14 января 2013 г .; 10.1371 / journal.pone.0054360
- ^ а б Бехар и др. 2008b, Рассвет человеческого матрилинейного разнообразия Am J Hum Genet. 2008 9 мая; 82 (5): 1130–1140
- ^ Коста MD и др. 2009 г., Данные полного секвенирования мтДНК тунисских долгожителей: тестирование ассоциации гаплогрупп и «золотая середина» долголетия. ([1] )
внешняя ссылка
- Яна Логана Гаплогруппа L2. Сайт митохондриальной ДНК
- Яна Логана L2bcd. Сайт митохондриальной ДНК
- Манниса ван Овена PhyloTree.org - поддерево мтДНК L
- Распространение гаплогруппы L2, из Национальная география