Гаплогруппа L3 (мтДНК) - Haplogroup L3 (mtDNA)
| Гаплогруппа L3 | |
|---|---|
| Возможное время происхождения | 80 000–60 000 YBP[1] |
| Возможное место происхождения | Восточная Африка[1][2] или же Азия[3] |
| Предок | L3'4 |
| Потомки | L3a, L3b'f, L3c'd, L3e'i'k'x, L3h, M, N |
| Определение мутаций | 769, 1018, 16311[4] |
Гаплогруппа L3 это митохондриальная ДНК человека (мтДНК) гаплогруппа. Клада сыграла ключевую роль в раннем расселении анатомически современные люди.
Это прочно связано с миграция из Африки современного человека около 70–50 000 лет назад. Он унаследован всеми современными неафриканскими популяциями, а также некоторыми популяциями в Африке.[5][6]
Источник

Гаплогруппа L3 возникла около 70000 лет назад, примерно во времена недавнее событие за пределами Африки. Это рассеяние произошло в Восточная Африка и распространился на Западную Азию, а затем в Южную и Юго-Восточную Азию в течение нескольких тысячелетий, и некоторые исследования показывают, что L3 участвовал в этой миграции из Африки. Оценка 2007 года для возраста L3 предполагает диапазон от 104 до 84 000 много лет назад.[7] Более поздние анализы, в том числе Soares et al. (2012) пришли к более поздней дате, примерно 70–60 000 лет назад. Соарес и др. также предполагают, что L3, скорее всего, распространился из Восточной Африки в Евразию примерно 65–55 000 лет назад в рамках недавнего события за пределами Африки, а также из Восточной Африки в Центральную Африку 60–35 000 лет назад.[1]В 2016 году Соарес и др. снова предположил, что гаплогруппа L3 возникла в Восточной Африке, что привело к миграции из Африки примерно 70-60 000 лет назад.[8]
Гаплогруппы L6 и L4 образуют сестринские клады L3, которые возникли в Восточной Африке примерно в то же время, но не участвовали в миграции за пределы Африки. Родовая клада L3'4'6 оценивается примерно в 110 тыс. лет назад, а L3'4 клад в 95 тыс. лет назад.[5]


а: Выход предшественника L3 в Евразию. б: Возвращение в Африку и распространение в Азию базальных линий L3 с последующей дифференциацией на обоих континентах.
Возможность происхождения L3 уже в Азии была также предложена Cabrera et al. (2018) на основе аналогичных дат слияния L3 и его распределенных по Евразии M и N производные клады (ок. 70 тыс. лет назад), удаленное местонахождение в Юго-Восточная Азия старейших субкладов M и N, и сопоставимый возраст отцовской гаплогруппы DE. Согласно этой гипотезе, после первоначальной миграции из Африки носителей pre-L3 (L3'4 *) около 125 тыс. Лет назад, должна была произойти обратная миграция самок, несущих L3, из Евразии в Восточную Африку где-то после 70 лет. кя. Предполагается, что эта обратная миграция связана с носителями отцовской гаплогруппы. E, который также предположительно возник в Евразии. Затем предполагается, что эти новые евразийские линии в значительной степени заменили старые автохтонные мужские и женские линии Северо-Восточной Африки.[3]
Согласно другим исследованиям, хотя и происходили более ранние миграции из Африки анатомически современных людей, нынешнее население Евразии происходит вместо более поздней миграции из Африки, датируемой примерно 65 000–50 000 лет назад (связанной с миграцией из L3).[9][2][10] Вай и др. (2019) предполагают, что из недавно обнаруженной старой и глубоко укоренившейся ветви материнской гаплогруппы N, обнаруженной в останках Северной Африки раннего неолита, гаплогруппа L3 возникла в Восточной Африке между 70 000-60 000 лет назад, и обе распространились в Африке и покинули Африку как часть миграции из Африки, с гаплогруппой N, расходящейся от нее вскоре (между 65 000-50 000 лет назад) либо в Аравии, либо, возможно, в Северной Африке, а гаплогруппа M возникла на Ближнем Востоке примерно в то же время, что и N.[2]
В исследовании Lipson et al (2019), анализирующем останки камерунского памятника Шум Лака нашел их более похожими на современные Пигмеи чем западноафриканцам, и предполагает, что несколько других групп (включая общих предков западноафриканцев, восточноафриканцев и предков неафриканцев), как правило, происходят от человеческой популяции, происходящей из Восточной Африки примерно 80 000-60 000 лет назад, которую они Предполагается, что около 70 000 лет назад источник и зона происхождения гаплогруппы L3.[11]
Распределение
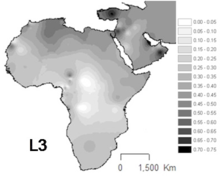
L3 часто встречается в Северо-Восточная Африка и некоторые другие части Восточной Африки,[12] в отличие от других частей Африки, где гаплогруппы L1 и L2 представляют около двух третей линий мтДНК.[13] Подлиния L3 также часто встречаются в Аравийский полуостров.
L3 подразделяется на несколько клад, две из которых породили макрогаплогруппы. M и N которые сегодня переносятся большинством людей за пределами Африки.[13] Существует по крайней мере одна относительно глубокая не-M, не-N клада L3 за пределами Африки, L3f1b6, которая встречается с частотой 1% в Астурия, Испания. Он отклонился от африканских линий L3 по крайней мере 10 000 лет назад.[14]
По данным Maca-Meyer et al. (2001), «L3 больше связан с Евразийский гаплогруппы, чем самые расходящиеся Африканский кластеры L1 и L2 ".[15] L3 - это гаплогруппа, от которой произошли все современные люди за пределами Африки.[16] Однако есть большая разнообразие из основных ветвей L3 в Африке, чем за ее пределами, двумя основными неафриканскими ветвями являются ответвления L3 M и N.
Распределение субкладов

L3 имеет семь равноудаленных потомков: L3a, L3b'f, L3c'd, L3e'i'k'x, L3h, M, N. Пять из них африканцы, а два связаны с Из Африки мероприятие.
- N – Евразия и части Африки из-за обратной миграции.[6][17]
- M – Азия, то Средиземноморский бассейн, и части Африки из-за обратной миграции.[6][17]
- L3a - Восточная Африка.[5][6] Частоты от умеренных до высоких, найденные среди Санье, Самбуру, Ирак, Яаку, Эль-Моло и другие малочисленные коренные народы из Восточноафриканская рифтовая долина. В Судане и в зоне Сахеля он встречается нечасто или отсутствует.[18]
- L3a1 - Встречается в Восточной Африке. Ориентировочный возраст 35,8–39,3 тыс. Лет назад.[6]
- L3a2 - Встречается в Восточной Африке. Ориентировочный возраст 48,3–57,7 тыс. Лет назад.[19][Примечание 1]
- L3b'f
- L3b - Распространение из Восточной Африки в верхний палеолит в Западно-Центральную Африку. Некоторые субклады распространились из Центральной Африки в Восточную Африку с миграцией банту.[6]
- L3f - Северо-Восточная Африка, Сахель, Аравийский полуостров, Иберия. Gaalien,[21] Бежа[21]
- L3f1
- L3f1a - Переносится мигрантами из Восточной Африки в Сахель и Центральную Африку.[6]
- L3f1b - Переносится мигрантами из Восточной Африки в Сахель и Центральную Африку.[6]
- L3f1b1 - Переносится из Центральной Африки в Южную и Восточную Африку с миграцией банту.[6]
- L3f1b1a - Поселился от Восточно-Центральной Африки до Центрально-Западной Африки, а также в Северной Африке и берберских регионах.[6]
- L3f1b4 - Переносится из Центральной Африки в Южную и Восточную Африку с миграцией банту.[6]
- L3f1b1 - Переносится из Центральной Африки в Южную и Восточную Африку с миграцией банту.[6]
- L3f1b6 - Редко, встречается в Иберии.[14]
- L3f2 - В основном распространен в Восточной Африке.[6] Также встречается в Северной и Центральной Африке.[20]
- L3f3 - Распространение от Восточной Африки до Чада и Сахеля около 8–9 тыс. Лет назад.[6] Найдено в Бассейн Чада.[20][22]
- L3f1
- L3c'd
- L3c - Чрезвычайно редкое происхождение, всего два образца, найденные до сих пор в Восточной Африке и Ближний Восток.[6]
- L3d - Распространение из Восточной Африки в верхний палеолит в Центральную Африку. Некоторые субклады распространились в Восточную Африку с миграцией банту.[6] Найдено среди Фулани,[5] Чадцы,[5] Эфиопы,[23] Народ акан,[24] Мозамбик,[23] Йеменцы,[23] Египтяне, Берберы[25]
- L3e'i'k'x
- L3e - Распространение из Восточной Африки в верхний палеолит в Западно-Центральную Африку. Это наиболее распространенная подклада L3 в банту говорящее население.[26] Предполагается, что L3e связан с центральноафриканским происхождением, а также является наиболее распространенным субкладом L3 среди афроамериканцев, Афро-бразильцы и Карибские острова[27]
- L3e1 - Распространение из Западной и Центральной Африки в Юго-Западную Африку с миграцией банту. Нашел в Ангола (6.8%).[28] Мозамбик, Суданский и Кикую из Кения а также в Йемене и среди Народ акан[24]
- L3e5 - Возникла в бассейне Чада. Нашел в Алжир,[29] а также Буркина-Фасо, Нигерия, Юг Тунис, Юг Марокко и Египет[30]
- L3i Практически исключительно встречается в Восточной Африке.[6]
- L3k - Редкая гаплогруппа в основном встречается в Северной Африке и Сахеле.[6][20]
- L3x - Практически исключительно встречается в Восточной Африке.[6] Найдено среди эфиопских Оромос,[23] Египтяне[Заметка 2][31]
- L3e - Распространение из Восточной Африки в верхний палеолит в Западно-Центральную Африку. Это наиболее распространенная подклада L3 в банту говорящее население.[26] Предполагается, что L3e связан с центральноафриканским происхождением, а также является наиболее распространенным субкладом L3 среди афроамериканцев, Афро-бразильцы и Карибские острова[27]
- L3h - Практически исключительно встречается в Восточной Африке.[6]
- L3h1 - В основном встречается в Восточной Африке, а ветви L3h1b1 спорадически встречаются в Сахеле и Северной Африке.[19][20]
- L3h2 - Встречается в Северо-Восточной Африке и на Сокотре. Откололся от других ветвей L3h еще 65–69 тыс. средний палеолит.[19][20]
Древние и исторические образцы
Гаплогруппа L3 наблюдалась в древней окаменелости, принадлежащей Докерамический неолит B культура.[32] L3x2a наблюдался в 4500-летнем охотничьем скоплении, раскопанном в Мота, Эфиопия, где древние окаменелости наиболее тесно связаны с современными Юго-запад Эфиопии населения.[33][34] Гаплогруппа L3 также была обнаружена среди древнеегипетский мумии (1/90; 1%), раскопанные на Абусир эль-Мелек археологические раскопки в Среднем Египте, а остальные происходят из евразийских субкладов, которые датируются пре-Птолемеев /поздно Новое Королевство и периоды Птолемея. Древние египетские мумии несли ближневосточный геномный компонент, наиболее близкий к современным ближневосточным.[35] Кроме того, гаплогруппа L3 наблюдалась в древних Гуанчи окаменелости, раскопанные в Гран-Канария и Тенерифе на Канарские острова, которые были радиоуглеродный между 7 и 11 веками нашей эры. Все кладоносные особи были ингумированы на территории Гран-Канарии, при этом было обнаружено, что большинство этих экземпляров принадлежат к субкладу L3b1a (3/4; 75%), а остальные с обоих островов (8/11; 72%) происходят от из евразийских субкладов. Скелеты гуанчей также несли автохтонный геномный компонент Магриби, который наиболее популярен среди современных Берберы, что предполагает, что они произошли от исконных берберских популяций, населявших северо-западный регион Аффаунден на высоком уровне.[36]
Разнообразие L3 было обнаружено в древних останках, связанных с пасторальным неолитом и пастырским железным веком Восточной Африки.[37]
| Культура | Генетический кластер или родство | Страна | Сайт | Дата | Материнская гаплогруппа | Отцовская гаплогруппа | Источник |
| Ранняя пастырская | PN | Кения | Овраг Преттджона (GsJi11) | 4060–3860 | L3f1b | – | Прендергаст 2019 |
| Пасторальный неолит | PN | Кения | Захоронение Коула (GrJj5a) | 3350–3180 | L3i2 | E-V32 | Прендергаст 2019 |
| Пасторальный неолит или эльментейтан | PN | Кения | Пещера Риго (GrJh3) | 2710–2380 | L3f | E-M293 | Прендергаст 2019 |
| Пасторальный неолит | PN | Кения | Naishi Rockshelter | 2750–2500 | L3x1a | E-V1515 (проб. E-M293) | Прендергаст 2019 |
| Пасторальный неолит | PN | Танзания | Пещера Гисимангеда | 2490–2350 | L3x1 | – | Прендергаст 2019 |
| Пасторальный неолит | PN | Кения | Могильник Найваша | 2350–2210 | L3h1a1 | E-M293 | Прендергаст 2019 |
| Пасторальный неолит | PN | Кения | Могильник Найваша | 2320–2150 | L3x1a | E-M293 | Прендергаст 2019 |
| Пасторальный неолит | PN | Танзания | Пещера Гисимангеда | 2150–2020 | L3i2 | E-M293 | Прендергаст 2019 |
| Пасторальный неолит или эльментейтан | PN | Кения | Пещера реки Нджоро II | 2110–1930 | L3h1a2a1 | – | Прендергаст 2019 |
| Пасторальный неолит | Нет данных | Танзания | Пещера Гисимангеда | 2000–1900 | L3h1a2a1 | – | Прендергаст 2019 |
| Пасторальный неолит | PN | Кения | Оль Калу | 1810–1620 | L3d1d | E-M293 | Прендергаст 2019 |
| Пасторальный железный век | PIA | Кения | Кисима Ферма, C4 | 1060–940 | L3h1a1 | E-M75 (кроме M98) | Прендергаст 2019 |
| Пасторальный железный век | PIA | Кения | Эмуруа Оле Полос (GvJh122) | 420–160 | L3h1a1 | E-M293 | Прендергаст 2019 |
| Пасторальный железный век | Выброс PN | Кения | Кокурматакоре | Нет данных | L3a2a | E-M35 (не E-M293) | Прендергаст 2019 |
Дерево
Этот филогенетическое дерево субклада гаплогруппы L3 основана на работе Манниса ван Овена и Манфреда Кайзера Обновленное комплексное филогенетическое дерево глобальных вариаций митохондриальной ДНК человека[4] и последующие опубликованные исследования.[38]
Самый недавний общий предок (MRCA)
- L1-6
- L2-6
- L2'3'4'6
- L3'4'6
- L3'4
- L3
- L3a
- L3a1
- L3a1a
- L3a1b
- L3a2
- L3a2a
- L3a1
- L3b'f
- L3b
- L3b1
- L3b1a
- L3b1a1
- L3b1a2
- L3b1a3
- L3b1a4
- L3b1a5
- L3b1a5a
- L3b1a6
- L3b1a7
- L3b1a7
- L3b1a8
- L3b1a9
- L3b1a9a
- L3b1a10
- L3b1a11
- L3b1b
- L3b1b1
- L3b1a
- L3b2
- L3b2a
- L3b2a
- L3b3
- L3b1
- L3f
- L3f1
- L3f1a
- L3f1a1
- L3f1b
- L3f1b1
- L3f1b2
- L3f1b2a
- L3f1b3
- L3f1b4
- L3f1b4a
- L3f1b4a1
- L3f1b4b
- L3f1b4c
- L3f1b4a
- L3f1b5
- L3f1a
- L3f2
- L3f2a
- L3f2b
- L3f3
- L3f3a
- L3f3b
- L3f1
- L3b
- L3c'd
- L3c
- L3d
- L3d1-5
- L3d1
- L3d1a
- L3d1a1
- L3d1a1a
- L3d1a1
- L3d1b
- L3d1b1
- L3d1c
- L3d1d
- L3d1a
- 199
- L3d2
- L3d5
- L3d3
- L3d3a
- L3d4
- L3d5
- L3d1
- L3d1-5
- L3e'i'k'x
- L3e
- L3e1
- L3e1a
- L3e1a1
- L3e1a1a
- 152
- L3e1a2
- L3e1a3
- L3e1a1
- L3e1b
- L3e1c
- L3e1d
- L3e1e
- L3e1a
- L3e2
- L3e2a
- L3e2a1
- L3e2a1a
- L3e2a1b
- L3e2a1b1
- L3e2a1
- L3e2b
- L3e2b1
- L3e2b1a
- L3e2b2
- L3e2b3
- L3e2b1
- L3e2a
- L3e3'4'5
- L3e3'4
- L3e3
- L3e3a
- L3e3b
- L3e3b1
- L3e4
- L3e3
- L3e5
- L3e3'4
- L3e1
- L3i
- L3i1
- L3i1a
- L3i1b
- L3i2
- L3i1
- L3k
- L3k1
- L3x
- L3x1
- L3x1a
- L3x1a1
- L3x1a2
- L3x1b
- L3x1a
- L3x2
- L3x2a
- L3x2a1
- L3x2a1a
- L3x2a1
- L3x2b
- L3x2a
- L3x1
- L3e
- L3h
- L3h1
- L3h1a
- L3h1a1
- L3h1a2
- L3h1a2a
- L3h1a2b
- L3h1b
- L3h1b1
- L3h1b1a
- L3h1b1a1
- L3h1b1a
- L3h1b2
- L3h1b1
- L3h1a
- L3h2
- L3h1
- M
- N
- L3a
- L3
- L3'4
- L3'4'6
- L2'3'4'6
- L2-6
Смотрите также
Филогенетическое дерево гаплогруппы митохондриальной ДНК человека (мтДНК) | |||||||||||||||||||||||||||||||||||||||
| Митохондриальная Ева (L ) | |||||||||||||||||||||||||||||||||||||||
| L0 | L1–6 | ||||||||||||||||||||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | L6 | ||||||||||||||||||||||||||||||||||
| M | N | ||||||||||||||||||||||||||||||||||||||
| CZ | D | E | грамм | Q | О | А | S | р | я | W | Икс | Y | |||||||||||||||||||||||||||
| C | Z | B | F | R0 | pre-JT | п | U | ||||||||||||||||||||||||||||||||
| HV | JT | K | |||||||||||||||||||||||||||||||||||||
| ЧАС | V | J | Т | ||||||||||||||||||||||||||||||||||||
Рекомендации
- ^ а б c Соарес, P; Alshamali, F; Pereira, J. B; Фернандес, V; Сильва, Н. М; Афонсу, К; Коста, M.D; Мусилова, Э; Маколей, V; Ричардс, М. Б.; Cerny, V; Перейра, Л. (2011). «Экспансия мтДНК гаплогруппы L3 в Африке и за ее пределами». Молекулярная биология и эволюция. 29 (3): 915–927. Дои:10.1093 / молбев / msr245. PMID 22096215.Точечная оценка 71,6 тыс. Лет назад, проведенная Соаресом и др. (2009), из 70,2 автора Fernandes et al. (2015).
- ^ а б c d Вай С., Сарно С., Лари М., Луизелли Д., Манци Дж., Галлинаро М., Матайч С., Хюбнер А., Моди А., Пилли Е., Тафури М. А., Карамелли Д., ди Лерния С. (март 2019 г.). «Митохондриальная линия предков N из неолитической« зеленой »Сахары». Научный представитель. 9 (1): 3530. Bibcode:2019НатСР ... 9,35 30В. Дои:10.1038 / s41598-019-39802-1. ЧВК 6401177. PMID 30837540.
- ^ а б c Кабрера В.М., Марреро П., Абу-Амеро К.К., Ларруга Дж. М. (июнь 2018 г.). «Носители базальных линий макрогаплогруппы L3 митохондриальной ДНК мигрировали обратно в Африку из Азии около 70 000 лет назад». BMC Эволюционная биология. 18 (1): 98. Дои:10.1186 / s12862-018-1211-4. ЧВК 6009813. PMID 29921229.
- ^ а б Ван Овен, Маннис; Кайзер, Манфред (2009). «Обновленное комплексное филогенетическое дерево глобальных вариаций митохондриальной ДНК человека». Человеческая мутация. 30 (2): E386–94. Дои:10.1002 / humu.20921. PMID 18853457. S2CID 27566749.
- ^ а б c d е Бехар, Дорон М .; Виллемс, Ричард; Судьял, Химла; Блю-Смит, Джейсон; Перейра, Луиза; Мецпалу, Эне; Скоццари, Розария; Маккан, Хиран; и другие. (2008). "Рассвет человеческого матрилинейного разнообразия" (PDF). Американский журнал генетики человека. 82 (5): 1130–40. Дои:10.1016 / j.ajhg.2008.04.002. ЧВК 2427203. PMID 18439549.
- ^ а б c d е ж грамм час я j k л м п о п q р s т Soares, P .; Alshamali, F .; Pereira, J. B .; Fernandes, V .; Silva, N. M .; Afonso, C .; Costa, M.D .; Мусилова, Е .; Маколей, В. (2011-11-16). «Экспансия мтДНК гаплогруппы L3 в Африке и за ее пределами». Молекулярная биология и эволюция. 29 (3): 915–927. CiteSeerX 10.1.1.923.345. Дои:10.1093 / молбев / msr245. ISSN 0737-4038. PMID 22096215.
- ^ Gonder, M. K .; Mortensen, H.M .; Рид, Ф. А .; De Sousa, A .; Тишкофф, С.А. (2006). «Анализ последовательности генома цельной мтДНК древних африканских линий». Молекулярная биология и эволюция. 24 (3): 757–68. Дои:10.1093 / molbev / msl209. PMID 17194802.
- ^ Соарес П., Рито Т., Перейра Л., Ричардс М. (март 2016 г.). «Генетический взгляд на предысторию Африки» (PDF). Африка из MIS 6-2. Палеобиология и палеоантропология позвоночных (1): 383–405. Дои:10.1007/978-94-017-7520-5_18. ISBN 978-94-017-7519-9.
- ^ Posth C, Renaud G, Mittnik M, Drucker DG, Rougier H, Cupillard C, Valentin F, Thevenet C, Furtwängler A, Wißing C, Francken M, Malina M, Bolus M, Lari M, Gigli E, Capecchi G, Crevecoeur I , Beauval C, Flas D, Germonpré M, van der Plicht J, Cottiaux R, Gély B, Ronchitelli A, Wehrberger K, Grigorescu D, Svoboda J, Semal P, Caramelli D, Bocherens H, Harvati K, Conard NJ, Haak W. , Пауэлл А., Краузе Дж. (2016). «Митохондриальные геномы плейстоцена предполагают единственное крупное расселение неафриканцев и позднеледниковую смену населения в Европе». Текущая биология. 26 (6): 827–833. Дои:10.1016 / j.cub.2016.01.037. HDL:2440/114930. PMID 26853362. S2CID 140098861.
- ^ Хабер М., Джонс А.Л., Коннел Б.А., Асан, Арчиеро Э., Хуанмин Й., Томас М.Г., Сюэ Ю., Тайлер-Смит К. (июнь 2019 г.). «Редкая африканская гаплогруппа Y-хромосомы D0 с глубокими корнями и ее значение для распространения современного человека за пределы Африки». Генетика. 212 (4): 1421–1428. Дои:10.1534 / генетика.119.302368. ЧВК 6707464. PMID 31196864.
- ^ ДНК древнего человека из Шум Лака (Камерун) в контексте истории населения Африки, Липсон Марк и др., 2019
- ^ Мартина Куянова; Луиза Перейра; Вероника Фернандес; Жоана Б. Перейра; Виктор Черный (2009). "Ближневосточный неолитический генетический вклад в небольшой оазис египетской Западной пустыни". Американский журнал физической антропологии. 140 (2): 336–46. Дои:10.1002 / ajpa.21078. PMID 19425100.
- ^ а б Уоллес, Д.; Браун, Мэриленд; Лотт, MT (1999). «Вариации митохондриальной ДНК в эволюции и болезни человека». Ген. 238 (1): 211–30. Дои:10.1016 / S0378-1119 (99) 00295-4. PMID 10570998.
- ^ а б Pardiñas, AF; Мартинес, JL; Рока, А; Гарсиа-Васкес, Э; Лопес, Б. (2014). «По пескам и далеко: интерпретация иберийской митохондриальной линии с древними западноафриканскими корнями». Являюсь. J. Hum. Биол. 26 (6): 777–83. Дои:10.1002 / ajhb.22601. PMID 25130626. S2CID 22184219.
- ^ Мака-Мейер, Николь; Гонсалес, Ана М; Ларруга, Хосе М.; Флорес, Карлос; Кабрера, Висенте М (2001). «Основные геномные митохондриальные линии определяют раннюю экспансию человека». BMC Genetics. 2: 13. Дои:10.1186/1471-2156-2-13. ЧВК 55343. PMID 11553319.
- ^ «Архивная копия». Архивировано из оригинал на 2011-07-08. Получено 2009-03-09.CS1 maint: заархивированная копия как заголовок (связь)
- ^ а б Висенте М. Кабрера (2017). «Носители основных линий макрогаплогруппы L3 митохондриальной ДНК мигрировали обратно в Африку из Азии около 70 000 лет назад». bioRxiv 10.1101/233502.CS1 maint: использует параметр авторов (связь)
- ^ Бору, Хирбо, Джибриль (2011). "Комплексная генетическая история популяций человека в Восточной Африке" (PDF): 118. Цитировать журнал требует
| журнал =(помощь) - ^ а б c d е Soares, P .; Alshamali, F .; Pereira, J. B .; Fernandes, V .; Silva, N. M .; Afonso, C .; Costa, M.D .; Мусилова, Е .; Macaulay, V .; Richards, M. B .; Cerny, V .; Перейра, Л. (16 ноября 2011 г.). «Экспансия мтДНК гаплогруппы L3 в Африке и за ее пределами». Молекулярная биология и эволюция. 29 (3): 915–927. Дои:10.1093 / молбев / msr245. PMID 22096215.CS1 maint: ref = harv (связь) Дополнительные данные на [1]
- ^ а б c d е ж грамм час я Эрнандес, Кандела Л; Соарес, Педро; Dugoujon, Jean M; Новеллетто, Андреа; Родригес, Хуан Н.; Рито, Тереза; Оливейра, Мариса; Мелхауи, Мохаммед; Баали, Абделлатиф; Перейра, Луиза; Кальдерон, Росарио (2015). «Ранние голоценовые и исторические африканские сигнатуры мтДНК на Пиренейском полуострове: Андалузский регион как парадигма». PLOS ONE. 10 (10): e0139784. Bibcode:2015PLoSO..1039784H. Дои:10.1371 / journal.pone.0139784. ЧВК 4624789. PMID 26509580. Дополнительные данные Дои:10.1371 / journal.pone.0139784.s006.
- ^ а б Мохамед, Хишам Юсиф Хасан. «Генетические закономерности вариации Y-хромосомы и митохондриальной ДНК с последствиями для населения Судана» (PDF). Хартумский университет. Получено 14 июн 2016.
- ^ Черный Виктор; Фернандес, Вероника; Коста, Марта Д.; Гайек, Мартин; Маллиган, Конни Дж; Перейра, Луиса (2009). «Миграция скотоводов, говорящих на чадском языке, в Африке на основе структуры населения бассейна Чада и филогеографии митохондриальной гаплогруппы L3f». BMC Эволюционная биология. 9: 63. Дои:10.1186/1471-2148-9-63. ЧВК 2680838. PMID 19309521.
- ^ а б c d Кивисилд, Т; Рейдла, М; Мецпалу, Э; Роза, А; Брем, А; Pennarun, E; Парик, Дж; Геберхивот, Т; и другие. (2004). «Эфиопское наследие митохондриальной ДНК: отслеживание потока генов через ворота слез и вокруг них». Американский журнал генетики человека. 75 (5): 752–70. Дои:10.1086/425161. ЧВК 1182106. PMID 15457403.
- ^ а б Fendt, Liane; Рёк, Александр; Циммерманн, Беттина; Боднер, Мартин; Тай, Торстен; Ченчер, Франк; Овусу-Дабо, Эллис; Гебель, Таня М.К .; Шнайдер, Питер М .; Парсон, Вальтер (2012). «Разнообразие мтДНК Ганы: криминалистический и филогеографический взгляд». Forensic Science International: генетика. 6 (2): 244–49. Дои:10.1016 / j.fsigen.2011.05.011. ЧВК 3314991. PMID 21723214.
- ^ Sheet1 - Патогены PLOS
- ^ Андерсон, С. 2006, Филогенетический и филогеографический анализ изменчивости африканской митохондриальной ДНК. В архиве 2011-09-10 на Wayback Machine
- ^ Bandelt, HJ; Алвес-Силва, Дж; Гимарайнш, ЧП; Сантос, MS; Брем, А; Перейра, L; Коппа, А; Ларруга, JM; и другие. (2001). "Филогеография митохондриальной гаплогруппы человека L3e: снимок африканской доисторической и атлантической работорговли". Анналы генетики человека. 65 (Pt 6): 549–63. Дои:10.1046 / j.1469-1809.2001.6560549.x. PMID 11851985. S2CID 221411246.
- ^ Plaza, Стефани; Салас, Антонио; Калафель, Франсеск; Корте-Реаль, Франсиско; Бертранпетит, Жауме; Карраседо, Анхель; Комас, Дэвид (2004). «Понимание западного распространения банту: анализ линии мтДНК в Анголе». Генетика человека. 115 (5): 439–47. Дои:10.1007 / s00439-004-1164-0. PMID 15340834. S2CID 13213447.
- ^ Асмахан Бекада; Лара Р. Арауна; Тахрия Деба; Франческ Калафель; Сорайя Бенхамамуш; Дэвид Комас (24 сентября 2015 г.). «Генетическая неоднородность алжирских популяций человека». PLOS ONE. 10 (9): e0138453. Bibcode:2015PLoSO..1038453B. Дои:10.1371 / journal.pone.0138453. ЧВК 4581715. PMID 26402429.; Стол S5
- ^ Fadhlaoui-Zid, K .; Plaza, S .; Calafell, F .; Бен Амор, М .; Comas, D .; Bennamar, A .; Gaaied, Эль (2004). «Гетерогенность митохондриальной ДНК у тунисских берберов». Анналы генетики человека. 68 (Pt 3): 222–33. Дои:10.1046 / j.1529-8817.2004.00096.x. PMID 15180702. S2CID 6407058.
- ^ Стеванович, А .; Gilles, A .; Bouzaid, E .; Kefi, R .; Paris, F .; Gayraud, R.P .; Spadoni, J. L .; Эль-Ченави, Ф .; Беро-Коломб, Э. (2004). «Разнообразие последовательностей митохондриальной ДНК у оседлого населения из Египта». Анналы генетики человека. 68 (Пт 1): 23–39. Дои:10.1046 / j.1529-8817.2003.00057.x. PMID 14748828. S2CID 44901197.
- ^ Фернандес, Ева; и другие. (2014). «Анализ древней ДНК 8000 г. до н.э. фермеров, проживающих на Ближнем Востоке, поддерживает пионерскую морскую колонизацию материковой Европы через Кипр и Эгейские острова в эпоху неолита». PLOS Genetics. 10 (6): e1004401. Дои:10.1371 / journal.pgen.1004401. ЧВК 4046922. PMID 24901650.
- ^ Видеть дополнительные материалы из Льоренте, М. Гальего; Jones, E. R .; Eriksson, A .; Сиска, В .; Артур, К. З .; Arthur, J. W .; Curtis, M.C .; Stock, J. T .; Coltorti, M .; Pieruccini, P .; Stretton, S .; Brock, F .; Higham, T .; Park, Y .; Hofreiter, M .; Bradley, D.G .; Bhak, J .; Pinhasi, R .; Маника, А. (13 ноября 2015 г.). «Древний эфиопский геном обнаруживает обширную евразийскую примесь в Восточной Африке». Наука. 350 (6262): 820–822. Bibcode:2015Научный ... 350..820л. Дои:10.1126 / science.aad2879. PMID 26449472.
- ^ Льоренте, М. Гальего; Jones, E. R .; Eriksson, A .; Сиска, В .; Артур, К. З .; Arthur, J. W .; Curtis, M.C .; Stock, J. T .; Колторти, М. (13 ноября 2015 г.). «Древний эфиопский геном обнаруживает обширную евразийскую примесь в Восточной Африке». Наука. 350 (6262): 820–822. Bibcode:2015Научный ... 350..820л. Дои:10.1126 / science.aad2879. PMID 26449472.
- ^ Schuenemann, Verena J .; и другие. (2017). «Геномы древних египетских мумий предполагают увеличение числа африканских предков к югу от Сахары в постримские периоды». Nature Communications. 8: 15694. Bibcode:2017НатКо ... 815694S. Дои:10.1038 / ncomms15694. ЧВК 5459999. PMID 28556824.
- ^ Родригес-Варела; и другие. (2017). «Геномный анализ человеческих останков с Канарских островов до европейского завоевания выявил близкое родство с современными североафриканцами». Текущая биология. 27 (1–7): 3396–3402.e5. Дои:10.1016 / j.cub.2017.09.059. PMID 29107554. Получено 27 октября 2017.
- ^ Прендергаст, Мэри Э .; Липсон, Марк; Савчук, Елизавета А .; Олальде, Иньиго; Огола, Кристина А .; Роланд, Надин; Сирак, Кендра А .; Адамски, Николь; Бернардос, Ребекка (30 мая 2019 г.). «Древняя ДНК свидетельствует о многоступенчатом распространении первых пастухов в Африку к югу от Сахары». Наука. 365 (6448): eaaw6275. Bibcode:2019Научный ... 365.6275P. Дои:10.1126 / science.aaw6275. ISSN 0036-8075. ЧВК 6827346. PMID 31147405.
- ^ "PhyloTree.org | дерево | L3". phylotree.org. Получено 2018-06-25.
Примечания
- ^ См. Supplemental_TreeUpdatedOctober.xls в разделе Дополнительные данные Соарес и др. 2011 г.
- ^ GUR46 в таблице 1. представляет собой гаплогруппу L3x2a мтДНК.
внешняя ссылка
- Общий
- Яна Логана Сайт митохондриальной ДНК
- Гаплогруппа L3
- Манниса ван Овена PhyloTree.org - поддерево мтДНК L3
- Распространение гаплогруппы L3, из Национальная география