Маркер размера молекулярной массы - Molecular-weight size marker

А маркер размера молекулярной массы, также называемый белок лестница, ДНК лестница, или же РНК лестница, представляет собой набор стандарты которые используются для идентификации приблизительный размер молекула бежать на гель в течение электрофорез, с использованием принцип который молекулярный вес обратно пропорциональна скорости миграции через гелевую матрицу. Поэтому при использовании в гель-электрофорез, маркеры эффективно обеспечивают логарифмическая шкала для оценки размера других фрагментов (при условии, что размеры фрагментов маркера известны).
Белковые, ДНК- и РНК-маркеры с заранее определенными размерами фрагментов и концентрациями коммерчески доступны. Их можно запустить в любом агароза или же полиакриламидные гели. Маркеры загружаются на дорожки, прилегающие к дорожкам для отбора проб, до начала цикла.
Маркеры ДНК

Разработка
Хотя концепция маркеров молекулярной массы сохранилась, методы разработки менялись на протяжении многих лет. Новые изобретения маркеров молекулярной массы распространяются в наборах, соответствующих типу маркера.
Первой проблемой при разработке маркеров было достижение высокого разрешения по всей длине маркера.[1] В зависимости от условий проведения гель-электрофореза фрагменты могли быть сжаты, что нарушало четкость изображения. Чтобы решить эту проблему, комплект для Саузерн Блот Анализ был разработан в 1990 году, предоставив первый маркер, объединяющий ДНК-мишень и ДНК зонда. В этом методе использовалось преимущество логарифмического интервала, и его можно было использовать для определения целевых диапазонов длиной более 20000 нуклеотиды.[2]
Дизайн
Существует два распространенных метода конструирования маркера размера молекулярной массы ДНК.[3] Один из таких методов использует технику частичного перевязка.[3] Лигирование ДНК - это процесс, при котором линейные части ДНК соединяются друг с другом через ковалентные связи; более конкретно, эти облигации фосфодиэфирные связи.[4] Здесь фрагмент дуплексной ДНК размером 100 п.н. частично лигирован. Следствием этого является то, что образуются димеры размером 200 пар оснований, тримеры размером 300 пар оснований, тетрамеры размером 400 пар оснований, пентамеры размером 500 пар оснований и т.д. Кроме того, останется часть дцДНК размером 100 п.н. В результате "лестница" ДНК, состоящая из кусочков ДНК известных молекулярная масса создается на геле.[3]
Второй метод предполагает использование рестрикционные ферменты и распознанная последовательность ДНК.[3] ДНК переварен с помощью определенного рестрикционного фермента, в результате чего образуются фрагменты ДНК различной молекулярной массы. Одним из преимуществ этого метода является то, что можно легко создать больше маркеров, просто переварив больше известной ДНК.[3] С другой стороны, размер кусочков ДНК зависит от участков, где рестрикционный фермент разрезает. Это затрудняет контроль размера фрагментов в маркере.[5]
В последнее время в лабораториях используется другой метод конструирования маркеров размера молекулярной массы ДНК. Эта стратегия предполагает использование Полимеразная цепная реакция (ПЦР).[5] Это достигается одним или двумя способами: 1) ДНК-мишень одновременно амплифицируется через грунтовка наборы, или 2) разные ДНК-мишени амплифицируются независимо через определенные праймеры.[5]
Влияние условий геля
Как и в случае с экспериментальными образцами, условия геля могут влиять на маркер размера молекулярной массы, который проходит рядом с ними. Такие факторы, как буфер, обвинять/Напряжение, и концентрация геля может повлиять на мобильность и / или внешний вид вашего маркера / лестницы / штандарта. Эти элементы необходимо учитывать при выборе маркера и при анализе окончательных результатов на геле.

- Буферы
- Буферы действуют для 1) установления pH и 2) обеспечения ионы для поддержки проводимости. В Электрофорез ДНК, то ТАЕ (трис-ацетат-ЭДТА) и TBE (трис-борат-EDTA) являются обычными предпочтительными буферами.[6] Буфер TBE предпочтителен для небольших фрагментов ДНК, тогда как TAE лучше подходит для фрагментов, превышающих 1500 пар оснований. Что касается емкости буферизации, TAE ниже по сравнению с TBE; это обычно приводит к более медленной подвижности ДНК. TBE также имеет лучшее разрешение.[7]
- Следует отметить, что воды не может служить заменой одного из этих буферов, так как ДНК не будет перемещаться по гелю.[6] Кроме того, использование воды вместо буфера приведет к образованию геля. таяние.[8]
- Заряд / Напряжение
- Что касается напряжения, рекомендуемый диапазон составляет от 4 до 10 В / см (т. Е. Вольт / см).[8] Гели агарозы обычно работают при напряжении 5 В / см.[3][6] Единица расстояния, см, относится к расстоянию между электроды (т.е. анод и катод ), а не длину самого геля.[3][6]
- Напряжения, слишком большие или меньшие, чем этот диапазон, будут влиять на подвижность и разрешение полос. Низкое напряжение снизит подвижность и приведет к расширению полос. С другой стороны, высокое напряжение уменьшит разрешение полос. Во многом это связано с тем, что слишком высокое напряжение может вызвать перегрев геля и даже его плавление.[8]
- Концентрация
- При выборе маркера необходимо учитывать концентрацию агарозы. Гель процент влияет на миграцию ДНК.[3][6] Как правило, чем выше концентрация геля, тем медленнее скорость движения ДНК через гель. Это в дополнение к той роли, которую молекулярный вес играет в миграции ДНК-маркера или образца, то есть чем выше молекулярный вес, тем медленнее будет мигрировать ДНК.[3][6]
- Концентрация геля также влияет на возможность визуализации полос, выходящих на гель. Меньшие полосы лучше разрешаются на геле с более высоким процентным содержанием, тогда как полосы с увеличенной молекулярной массой легче визуализировать на геле с более низким процентным содержанием.[6]
Белковые маркеры
Разработка
Раньше белковые маркеры разрабатывались с использованием различных цельных белков. Разработка набора, включающего маркер размера молекулярной массы на основе фрагментов белка, началась в 1993 году. Этот маркер белка, состоящий из 49 различных аминокислота последовательности, включая мультидоменные белки, и позволил анализировать белки, расщепленные в разных местах.[9]
Современные технические усовершенствования белковых маркеров включают использование саморазвития. Первый саморазвивающийся белковый маркер с регулярным весом был изобретен в 2012 году.[10]
Дизайн
Подобно ДНК-маркерам, эти маркеры обычно состоят из очищенных белков, молекулярные массы которых уже известны.[3] В приведенном ниже списке указаны некоторые белки, а также молекулярная масса, которые обычно используются при создании маркера белка.

| Протеин | Молекулярная масса (кДа ) |
| Бета-галактозидаза | 120[11] |
| Фосфорилаза B | 94[3][12] |
| Альбумин бычьей сыворотки (BSA) | 67[3][12] |
| Овальбумин | 43[3] |
| индюк Альбумин | 40[12] |
| Карбоангидраза | 30[3][12] |
| Соя Трипсин Ингибитор | 20.1[3][12] |
| а-лактальбумин | 14.4[3][12] |
| Лизоцим | 14[13] |
Выбор правильного белкового маркера
Маркеры размера молекулярной массы можно разделить на две категории: маркеры молекулярной массы и маркеры молекулярной лестницы.[14] Маркеры бывают окрашенными или неокрашенными, и в зависимости от обстоятельств один может быть более подходящим, чем другой. Маркеры размера молекулярной массы также могут быть изменены биохимически.[15] Спряжение с биотин самый распространенный. Маркеры размера молекулярной массы чаще всего используются в Электрофорез в SDS-полиакриламидном геле и вестерн-блоттинг.С учетом всех различных типов и применений маркеров размера молекулярной массы важно выбрать соответствующий стандарт белка. Помимо наиболее распространенного использования, как способа расчета молекулярной массы образцов, другие применения включают предоставление визуальных доказательств миграции и эффективности переноса белка, а иногда даже используются для положительного контроля.[16]
- Маркер MW против протеиновых лестниц
- Маркер молекулярной массы - это один из типов стандарта белка. Они могут быть предварительно окрашенными или неокрашенными перед загрузкой; в зависимости от типа эксперимента один может быть более выгодным. В любом случае они обычно проходят по внешней дорожке геля, а образец загружается по средним дорожкам.[14] Молекулярные маркеры отличаются от белковых лестниц тем, что они состоят из смеси родные белки, спецификации которых хорошо классифицированы, но не соответствуют целым числам.[14] Обычно они намного дешевле, но анализ позволяет получить только приблизительное значение белков, разделенных электрофорезом.[14]
- Белковая лестница - это еще один тип стандарта белка. Они почти всегда в пятнах.[14] Белковые лестницы отличаются от молекулярных маркеров тем, что они состоят из смеси высокоочищенных белков, характеристики которых известны и соответствуют целым числам.[14] Обычно белковые лестницы состоят из 10-12 белков.[14] В конце эксперимента, после того как произойдет миграция размера, одна полоса будет представлять размер каждого белка, содержащегося в лестнице.[17] Маркеры расположены равномерно, и анализ размера с использованием этих маркеров позволяет точно определить интересующий белок. В некоторых случаях в качестве метода молекулярного подтверждения MW-маркеры используются с белковыми лестницами для проверки.[14]
- Предварительно окрашенные и неокрашенные маркеры
- Белковые маркеры могут быть неокрашенными или предварительно окрашенными, но у обоих есть свои преимущества и недостатки.[18] Простая визуализация разделения и переноса белков стала возможной благодаря использованию предварительно окрашенных маркеров.[18] Они обычно используются как в электрофорезе в SDS-полиакриламидном геле, так и в вестерн-блоттинге. В SDS-PAGE он позволяет отслеживать миграцию белка, поскольку полосы белка будут разделяться и их можно будет увидеть во время электрофоретического анализа. При вестерн-блоттинге окрашенные стандарты белка позволяют визуализировать перенос белка на мембрану.[17] Однако определение размера не так точный с этими маркерами (дополнительные пояснения см. в разделе «Рекомбинантные и природные маркеры»).[18]
- Хотя неокрашенные маркеры позволяют более точно определить размер, их нельзя увидеть во время растекания геля. Таким образом, гель необходимо окрасить, чтобы увидеть полосы.[19]
- Рекомбинантные и природные маркеры
- Помимо окрашенных и неокрашенных маркеров, белковые маркеры можно рассматривать как рекомбинантные и природные.[18] Рекомбинантные маркеры состоят из рекомбинантных белков, которые были сильно очищены. Эти маркеры разработаны таким образом, чтобы выделять определенные характеристики.[18] Примеры этих характеристик включают: Теги сходства и молекулярные массы, которые равномерно расположены относительно друг друга.[18]
- Природные маркеры, как следует из названия, представляют собой смесь белков, которые встречаются в природе.[18] Предварительно окрашенные натуральные маркеры хорошо подходят для визуализации отделения геля. Однако эти маркеры имеют тенденцию связываться с пятном в ковалентный способом в разных количествах и на разных позициях.[18] Следовательно, результирующие полосы могут быть шире. Это особенно верно при сравнении с предварительно окрашенными рекомбинантными маркерами. Из-за этого эффекта определения молекулярной массы, вероятно, будут менее точными с предварительно окрашенными натуральными маркерами.[18]
- Биохимически измененный
- Стандарты белков также можно изменять химически. Обычное изменение связано с использованием биотин. Биотин имеет очень высокое сродство к стрептавидин, и поэтому связывание образует очень прочный комплекс. Для визуализации к стрептавидину прикрепляют цветовую метку.[15]
Влияние условий геля
Как и в случае с электрофорезом ДНК, при выборе белкового маркера следует учитывать такие условия, как буферность, заряд / напряжение и концентрация.
- Буферы
- Буферы могут влиять на подвижность как маркера, так и образцов. PH буфера варьируется в зависимости от используемой системы, и, следовательно, каждая буферная система будет по-разному влиять на заряд белка или белков.[20] Кроме того, в случае SDS-PAGE связывающая аффинность на SDS может влиять система буферизации.[20] Даже при использовании того же процент и тип геля, одни и те же белки будут мигрировать с разной скоростью в зависимости от используемого буфера.[20]
- Заряд / Напряжение
- Напряжение играет роль в подвижности белков на геле. Белки будут мигрировать быстрее при более высоких напряжениях. Следовательно, время растекания геля будет короче. И наоборот, более высокие напряжения могут привести к большей диффузии полосы.[20] Кроме того, если напряжение слишком высокое, температура в камере электрофореза может стать таким, что гель начнет таять.[20]
- Напряжение, при котором должен работать гель, зависит от типа геля. Для некоторых гелей напряжение остается постоянным на протяжении всего цикла, тогда как для других гелей начальное напряжение может оставаться постоянным в течение определенного времени, прежде чем оно будет увеличено.[20] Это второе напряжение затем используется в течение определенного периода времени, после чего оно также может быть увеличено.[20]
- Концентрация
- В процентном отношении гели, используемые для электрофореза белков, можно разделить на однопроцентные гели и градиентные гели.[18] Однопроцентные гели также называют линейными гелями.[20] Для линейных гелей выбранный процент обычно составляет от 7,5% до 20%.[18] Обычные процентные диапазоны для градиентных гелей составляют 4-15% и 10-20%. У каждого вида геля есть свои преимущества.[18] Например, линейные гели предпочтительны, когда несколько белков имеют одинаковые молекулярные массы; лучшее разделение между этими белками будет отображаться линейным гелем.[18] С другой стороны, градиентные гели являются лучшим выбором, когда представляющие интерес образцы содержат белки с сильно различающейся молекулярной массой или которые охватывают широкий диапазон молекулярных масс.[18][20]
Маркеры РНК
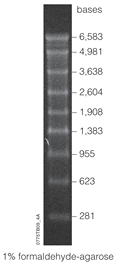
Разработка
РНК-лестницы, состоящие из маркеров размера молекулярной массы РНК, были первоначально разработаны с использованием метода синтетического круга.[21] для изготовления маркеров разного размера. Этот метод был усовершенствован изобретателем Эриком Т. Кулом для использования кольцевой ДНК. векторов как метод получения маркеров размера молекулярной массы РНК. Так называемый метод катящегося круга, усовершенствования этого метода связаны с его эффективностью в синтезе РНК. олигонуклеотиды. Из круглого шаблона ДНК, одноцепочечная РНК длиной от 4 до 1500 п.н. могут быть получены без использования праймеров и путем переработки нуклеотидтрифосфат. ДНК также можно синтезировать из круглого шаблона, что делает этот метод более универсальным. В сравнении с транскрипция стока, метод синтетического круга дает олигонуклеотиды РНК без стока. В сравнении с ПЦР, метод синтетического круга дает олигонуклеотиды РНК без необходимости использования полимеразы или термоциклер. Этот метод также является рентабельным, так как позволяет синтезировать большие количества продукта с меньшим количеством ошибок, чем машинные синтезаторы.[21]
Дизайн
Маркеры РНК состоят из транскриптов РНК различной увеличивающейся длины. Например, маркер Lonza 0.5-9 kbp[22] имеет полосы с маркировкой 0,5, 1, 1,5, 2, 2,5, 3, 4, 5, 6 и 9 килобаза пары. Маркеры растворяются в буфере для хранения, например в EDTA, и может иметь срок годности до 2 лет при хранении при -80 ° C. Чтобы использовать маркер, например, для Нозерн-блот-анализа, сначала размороженный, а затем окрашивают так, чтобы его можно было обнаружить при гель-электрофорезе. Одним из наиболее распространенных красителей, используемых для маркеров, является этидиум бромид.
Диапазон конкретного маркера относится к множеству диапазонов, которые он может отображать. «Высокий» диапазон относится к относительно большим фрагментам (измеряется в т.п.н.), тогда как «низкий» диапазон относится к маркерам, которые различают небольшие фрагменты (измеряются в п.о.). Некоторые маркеры даже можно охарактеризовать как «сверхнизкий диапазон»,[16] но еще более точным является маркер микроРНК. Маркер микроРНК можно использовать для измерения фрагментов РНК в пределах дюжины нуклеотидов, таких как маркер микроРНК 17-25 нуклеотидов.[23]
Использовать
При эквивалентной молекулярной массе РНК будет мигрировать быстрее, чем ДНК. Однако и РНК, и ДНК имеют отрицательный линейный склон между их расстоянием миграции и логарифмический молекулярный вес.[24] То есть образцы меньшего веса могут перемещаться на большее расстояние. Это соотношение необходимо учитывать при выборе РНК или ДНК-маркеров в качестве стандарта.
При нанесении маркеров РНК и образцов РНК на гель важно предотвратить нуклеаза загрязнение, так как РНК очень чувствительна к рибонуклеаза (РНКазы) деградация через катализ.[25][26] Таким образом, необходимо учитывать все материалы, которые будут использоваться в процедуре. Любая стеклянная посуда, которая должна контактировать с РНК, должна быть предварительно обработана диэтилпирокарбонат (DEPC) и пластмассовые материалы должны быть одноразовыми.[25]
Маркеры размера молекулярной массы и SDS-PAGE

Одно из наиболее распространенных применений маркеров размера молекулярной массы - электрофорез в геле. Цель гель-электрофореза - разделить белки физический или же химические свойства, которые включают заряд, размер молекулы и pH. <При разделении по размеру идеальным методом является SDS-PAGE или электрофорез в полиакриламидном геле и маркеры размера молекулярной массы являются подходящими стандартами для использования.
Гели могут различаться по размеру. Подходящий размер геля будет зависеть от количества исследуемых образцов. Все гели разделены на полосы, которые проходят параллельно через гель. Каждая дорожка будет содержать определенный образец. Обычно стандарты размера молекулярной массы размещаются на внешней дорожке. Если гель имеет особенно большое количество полосок, то для большей прозрачности можно разместить несколько лестниц поперек геля.
Белки и стандарты пипетированный на геле в соответствующих полосах. Додецилсульфат натрия (SDS) взаимодействует с белками, денатурирующий их и придавая им отрицательный заряд. Поскольку все белки имеют одинаковое отношение заряда к массе, подвижность белка через гель будет зависеть исключительно от молекулярной массы. Как только электрическое поле будет включено, начнется миграция белка. По завершении можно использовать механизм обнаружения, такой как вестерн-блоттинг, который выявит наличие полос. Каждая полоса представляет определенный белок. Расстояние путешествия зависит исключительно от молекулярной массы; следовательно, молекулярная масса каждого белка может быть определена путем сравнения расстояния между неизвестным белком и стандартом известной молекулярной массы.[27]
Различное использование маркеров размера молекулярной массы
Существует множество типов маркеров размера молекулярной массы, и каждый из них обладает уникальными характеристиками, что позволяет использовать их в ряде биологических методов. Выбор маркера размера молекулярной массы зависит от типа маркера (ДНК, РНК или белок) и предлагаемого диапазона длин (например, 1 килобайт). Прежде чем выбрать маркер размера молекулярной массы, важно ознакомиться с этими характеристиками и свойствами. В конкретном случае один тип может быть более подходящим, чем другой. Хотя конкретные маркеры могут различаться в разных протоколах для данной техники, в этом разделе будут описаны общие маркеры и их роли.
Аллозимы
Был разработан первый тип молекулярного маркера, который был исследован на гель-электрофорезе. аллозимы. Эти маркеры используются для обнаружения изменений белка. Слово «аллозим» (также известное как «аллоэнзим») происходит от «аллельный варианты ферменты."[28] При работе с гелем белки разделяются по размеру и заряду. Хотя аллозимы могут показаться устаревшими по сравнению с другими доступными маркерами, они все еще используются сегодня, в основном из-за их низкой стоимости. Одним из основных недостатков является то, что, поскольку доступно только ограниченное количество, специфичность является проблемой.[28]
Маркеры на основе ДНК (1960-е)
Хотя аллозимы могут обнаруживать вариации ДНК, это косвенный метод и не очень точный. Маркеры на основе ДНК были разработаны в 1960-х годах.[28] Эти маркеры намного более эффективны при различении вариантов ДНК. Сегодня это наиболее часто используемые маркеры. Маркеры на основе ДНК работают, исследуя нуклеотиды, которые могут выполнять множество функций, таких как обнаружение различий в нуклеотидах или даже количественное определение количества нуклеотидов. мутации.[28]
- RFLP
- Полиморфизма длин рестрикционных фрагментов это метод, используемый для обнаружения изменений в гомологичный ДНК.[29] Особое ограничение эндонуклеазы используются для переваривания ДНК. Молекулярный маркер RFLP специфичен для одного фрагмента. Наряду с аллельными маркерами RFLP, маркер размера молекулярной массы, в данном случае ДНК-маркер,[30] также включен в электорфорезированный агарозный гель. Маркер ДНК позволяет оценить размер рестрикционных фрагментов.
- Мини-спутники
- Подобно RFLP, этот метод также использует эндонуклеазы рестрикции для переваривания геномной ДНК. Мини-спутники представляют собой короткие последовательности тандемных повторов, приблизительно 10-60 пар оснований. Минисателлиты можно использовать для снятия отпечатков ДНК и в качестве регуляторов генного контроля.[28]
Маркеры на основе ПЦР (1980-е годы)
Успех маркеров на основе ДНК привел к развитию ПЦР. ПЦР (полимеразной цепной реакции ) является ДНК усиление техника, которая может применяться к различным типам фрагментов. До этой разработки для амплификации ДНК ее нужно было клонировать или изолировать. Вскоре после открытия ПЦР возникла идея использовать маркеры на основе ПЦР для гель-электрофореза. Этот тип маркеров основан на ПЦР. грунтовки и относятся к категории последовательности ДНК полиморфизм.[28]
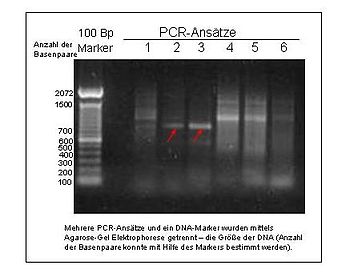
- Микроспутники
- Также известен как SSR (простая последовательность повторений ) или STR (короткие тандемные повторы ), микроспутники отличаются от минисателлитов тем, что они короче, обычно 2-6 пар оснований. Это свойство микроспутников позволяет легко изолировать их. Микроспутники чаще всего используются в популяционная генетика. Микросателлиты имеют высокую и сложную частоту мутаций, что является их основным недостатком.[28]
- AFLP
- Полиморфизм длины амплифицированного фрагмента на основе ПЦР Дактилоскопия ДНК техника. ДНК сначала переваривается эндонуклеазами. В рестрикционные фрагменты затем лигируют вместе.[31] Затем создается молекулярный маркер, когда выбираются определенные фрагменты для усиление. Маркеры AFLP наносятся вместе с маркером ДНК на гель. Обычный ДНК-маркер AFLP имеет длину 30–330 п.н.[32] Фрагменты этого маркера лежат с интервалом 10 п.н. для повышения точности.
- RAPD
- Случайно амплифицированная полиморфная ДНК это метод, который проводится аналогично AFLP. Разница в том, что молекулярные маркеры генерируются случайным образом.[31] Наиболее распространенным маркером размера молекулярной массы для этого метода является лестница ДНК размером 1 КБ.[33][34]
Полиморфизм последовательности ДНК
Хотя технически говоря, полиморфизм последовательности ДНК имеет место с момента использования RFLP в 1960-х годах, анализ значительно изменился с годами. Полиморфизм последовательности ДНК использует более старые методы, такие как RFLP, но в более крупном масштабе. Секвенирование происходит намного быстрее и эффективнее. Анализ автоматизирован, поскольку он использует метод, известный как секвенирование дробовика. Этот высокопроизводительный метод обычно используется в популяционной генетике.[28]
- SNP
- SNP (однонуклеотидный полиморфизм ), используются для обнаружения вариаций одиночных нуклеотидов. Методика очень похожа на RFLP. SNP часто используются для популяционных генетических исследований.[35] После амплификации с помощью ПЦР эти небольшие фрагменты можно визуализировать с помощью гель-электрофореза, и снова ДНК-маркеры играют роль в определении длины фрагмента.
Анализ полисахаридов методом электрофореза в углеводном геле
Углеводы маркеры используются в технике, известной как полисахарид анализ методом электрофореза в углеводном геле (ПАСЕ), который представляет собой измеримый метод разделения.[36]Это позволяет анализировать фермент гидролиз товары.[36] Он использовался в таких приложениях, как характеристика ферментов, участвующих в гемицеллюлоза деградация, определение структуры полисахаридов гемицеллюлозы и анализ ферментативного расщепления целлюлоза товары.[36]
PACE зависит от дериватизации, которая представляет собой преобразование химического соединения в производная.[36][37] Здесь моносахариды, олигосахариды, и полисахариды представляют интерес. Они помечены на их переходных концах знаком флуоресцентная метка (т.е. флуорофор ).[36] Эта дериватизация с помощью флуорофора позволяет как разделение на геле при желаемых обстоятельствах, так и флуоресценция визуализация геля. В этом случае используется полиакриламидный гель.[36]
Как и в случае электрофореза ДНК, РНК и белков, маркеры анализируются вместе с интересующими образцами при электрофорезе в углеводном геле.[36] Маркеры состоят из олигосахаридов с известной молекулярной массой. Подобно интересующим образцам, маркер также дериватизируется с помощью флуорофора (обычно с 8-аминогруппой).нафталин -1,3,6-трисульфоновая кислота (ANTS) или 2-аминоакридон ).[36]
Рекомендации
- ^ Карлсон, Дэвид П. «Маркеры размера для электрофоретического анализа ДНК». Патент США № 5316908A.. Патенты Google. Получено 30 октября 2013.
- ^ Карлсон, Дэвид П. «Маркеры размера для электрофоретического анализа ДНК». Патент EP № 0466404B1.. Патенты Google. Получено 30 октября 2013.
- ^ а б c d е ж грамм час я j k л м п о п Блабер, Майк. «Лекция 20: Гель-электрофорез». BCH5425 Молекулярная биология и биотехнология.
- ^ Боуэн, Р. (20 октября 1999 г.). «Лигирование ДНК». Биотехнология и генная инженерия. Получено 12 ноября 2013.
- ^ а б c Лан, Во Тхи Тхуонг; Ссуда, Фам Тхи Тхань; Дуонг, Фам Ань Туи; Тхань, Ле Тхи; Ха, Нго Тхи; Туан, Та Бич (2012). «Простая процедура лабораторного производства ДНК-лестницы». Журнал нуклеиновых кислот. 2012: 254630. Дои:10.1155/2012/254630. ЧВК 3306978. PMID 22496965.
- ^ а б c d е ж грамм Боуэн, Р. (2000). «Электрофорез ДНК в агарозном геле». Гипертексты для биомедицинских наук - Государственный университет Колорадо.
- ^ «Буфер трис-борат-ЭДТА и трис-ацетат-ЭДТА (TAE и TBE, pH 8,3)» (PDF). Аниара.
- ^ а б c "Советы и приемы электрофореза в агарозном геле". Технологии жизни.
- ^ Хартли, Джеймс. «Маркерная лестница размера белка». Патент США № 5449758A.. Патенты Google. Получено 30 октября 2013.
- ^ Ченг, Тиан Лу. «Саморазвивающийся и регулярно взвешиваемый набор маркеров молекулярной массы белка и метод его приготовления». Патент США № 20130217133A1.. Патенты Google. Получено 30 октября 2013.
- ^ «Маркер молекулярной массы предварительно окрашенного белка». Термонаучный. Получено 12 ноября 2013.
- ^ а б c d е ж Ингельман, Маргарета (2004). «Разделение и анализ белков». KE7001 Биохимические лаборатории. Получено 12 ноября 2013.
- ^ «Маркеры молекулярной массы белка». Помощь Биотех. 2011. Получено 12 ноября 2013.
- ^ а б c d е ж грамм час «Руководство по сравнению и выбору маркеров молекулярной массы белков». Получено 16 ноября 2013.
- ^ а б «Биотинилированный маркер молекулярной массы». Получено 16 ноября 2013.
- ^ а б «Маркеры молекулярной массы». Получено 16 ноября 2013.
- ^ а б «Маркер молекулярной массы предварительно окрашенного протеина Pierce». Получено 16 ноября 2013.
- ^ а б c d е ж грамм час я j k л м п "Электрофорез - руководство по электрофорезу и детекции в полиакриламидном геле" (PDF). Био-Рад.
- ^ «Руководство по сравнению и выбору маркеров молекулярной массы белков». Термонаучный. Получено 12 ноября 2013.
- ^ а б c d е ж грамм час я «Справочник по белкам 2013» (PDF). Технологии жизни.
- ^ а б Кул, Эрик Т. «Круговые ДНК-векторы для синтеза РНК и ДНК». Патент США № 6096880A.. Патенты Google. Получено 27 ноября 2013.
- ^ Лонза. «Маркеры РНК 0,5–9 т.п.н.» (PDF). Документ № 18123-0807-06.. Lonza Rockland Inc. Получено 27 ноября 2013.
- ^ Биолаборатории Новой Англии. «Маркер микроРНК». Биолаборатории Новой Англии. Получено 27 ноября 2013.
- ^ Уикс, Ричард Дж. (1986).«Определение молекулярной массы РНК с помощью электрофореза в агарозном геле с использованием формальдегида в качестве денатурирующего агента: Сравнение маркеров молекулярной массы РНК и ДНК». Международный журнал биохимии. 18 (3): 277–278. Дои:10.1016 / 0020-711x (86) 90118-7. PMID 2937672.
- ^ а б "Каталожный №: R0004". РНК-маркер High Easy. Абнова. Получено 14 декабря 2013.
- ^ «Электрофорез РНК: Введение». РНК-электрофорез. Thermo Fisher Scientific Inc.. Получено 14 декабря 2013.
- ^ «Определение молекулярной массы белков» (PDF). Получено 14 декабря 2013.
- ^ а б c d е ж грамм час Шлоттерер, Кристиан. «Эволюция молекулярных маркеров» (PDF). Получено 26 ноября 2013.
- ^ «Благоустройство населения». Получено 30 октября 2013.
- ^ Хиггинс, Л. (апрель 2012 г.). «ДНК-лестница для гель-электрофореза». Колледж Льюиса и Кларка. Получено 15 ноября 2013.
- ^ а б Мюллер, Ульрих (1999). «Генотипирование AFLP и дактилоскопия» (PDF). Тенденции в экологии и эволюции. 14 (10): 389–394. Дои:10.1016 / s0169-5347 (99) 01659-6. PMID 10481200. Получено 30 октября 2013.
- ^ Корпорация Invitrogen (2003 г.). «30–330 п.н. AFLP® DNA Ladder» (PDF). Руководство. Корпорация Life Technologies. Получено 15 ноября 2013.
- ^ Джаннини, Кристина; и другие. (Май 2004 г.). «unRAPD анализ мтДНК из цветков томата без ядерных артефактов ДНК» (PDF). Биотехнологии. 36 (5): 772–776. Дои:10.2144 / 04365BM04. PMID 15152595. Получено 15 ноября 2013.
- ^ Робертс, М. А .; Кроуфорд, Д. Л. (1 июня 2000 г.). «Использование случайно усиленной полиморфной ДНК в качестве средства разработки ДНК-зондов Streptomyces, специфичных для определенного рода и штамма». Прикладная и экологическая микробиология. 66 (6): 2555–2564. Дои:10.1128 / AEM.66.6.2555-2564.2000. ЧВК 110581. PMID 10831438.
- ^ МакКлин, Филипп. «Классы молекулярных маркеров». Получено 30 октября 2013.
- ^ а б c d е ж грамм час Косик, Ондрей; Бромли, Дженнифер Р .; Буссе-Вичер, Марта; Чжан, Чжинун; Дюпри, Поль (2012). «Исследования ферментативного расщепления целлюлозы с использованием анализа полисахаридов с помощью электрофореза в углеводном геле (PACE)». Методы в энзимологии. 510: 51–67. Дои:10.1016 / B978-0-12-415931-0.00004-5. ISBN 9780124159310. ISSN 0076-6879. PMID 22608721.
- ^ Cammack R, Attwood TK, Campbell PN, Parish HJ, Smith A, Stirling JL, Vella F (2006). «Флуорофор». Оксфордский словарь биохимии и молекулярной биологии (Второе изд.). Издательство Оксфордского университета.