Электрофорез в агарозном геле - Agarose gel electrophoresis
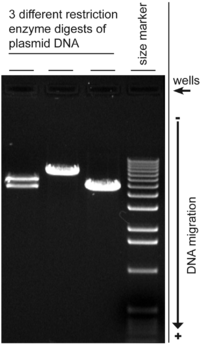
Электрофорез в агарозном геле это метод гель-электрофорез используется в биохимия, молекулярная биология, генетика, и клиническая химия для разделения смешанной популяции макромолекул, таких как ДНК или белки, в матрице агароза, один из двух основных компонентов агар. Белки могут быть разделены по заряду и / или размеру (изоэлектрическая фокусировка электрофорез в агарозе по существу не зависит от размера), а ДНК и РНК фрагменты по длине.[1] Биомолекулы разделяются путем нанесения электрическое поле для перемещения заряженных молекул через матрицу агарозы, и биомолекулы разделяются по размеру в матрице геля агарозы.[2]
Гель из агарозы легко отливать, имеет относительно меньше заряженных групп и особенно подходит для разделения ДНК в диапазоне размеров, наиболее часто встречающихся в лабораториях, что объясняет популярность его использования. Отделенную ДНК можно рассматривать с помощью красителя, чаще всего в УФ-свете, и фрагменты ДНК можно относительно легко экстрагировать из геля. Большинство используемых гелей агарозы растворены на 0,7–2% в подходящем буфере для электрофореза.
Свойства агарозного геля

Агароза гель представляет собой трехмерную матрицу, образованную спиральными молекулами агарозы в суперспиральных пучках, которые агрегированы в трехмерные структуры с каналами и порами, через которые могут проходить биомолекулы.[3] Трехмерная структура удерживается вместе водородными связями и поэтому может быть разрушена путем нагревания до жидкого состояния. Температура плавления отличается от температуры гелеобразования, в зависимости от источников, агарозный гель имеет температуру гелеобразования 35–42 ° C и температуру плавления 85–95 ° C. Также доступны агарозы с низкой температурой плавления и с низким содержанием геля, полученные путем химических модификаций.
Гель агарозы имеет большой размер пор и хорошую прочность геля, что делает его подходящим в качестве противоконвекционной среды для электрофореза ДНК и больших молекул белка. Размер пор 1% геля оценивается от 100 нм до 200–500 нм.[4][5] а его прочность геля позволяет гелям с разбавлением до 0,15% образовывать пластину для гель-электрофореза.[6] Однако гели с низкой концентрацией (0,1–0,2%) хрупкие, и поэтому с ними трудно обращаться. Гель агарозы имеет более низкую разрешающую способность, чем гель полиакриламида, для ДНК, но имеет больший диапазон разделения, и поэтому используется для фрагментов ДНК размером обычно 50–20 000 п.н. Предел разрешения для стандартного электрофореза в агарозном геле составляет около 750 кб, но разрешение более 6 мб возможно с гель-электрофорез в импульсном поле (PFGE).[7] Его также можно использовать для разделения крупных белков, и он является предпочтительной матрицей для гель-электрофореза частиц с эффективными радиусами более 5-10 нм. 0,9% -ный гель агарозы имеет достаточно большие поры для проникновения бактериофаг Т4.[6]
Полимер агарозы содержит заряженные группы, в частности пируват и сульфат.[8] Эти отрицательно заряженные группы создают поток воды в направлении, противоположном движению ДНК в процессе, называемом электроэндосмос (EEO), и, следовательно, может замедлять движение ДНК и вызывать размытие полос. Гели с более высокой концентрацией будут иметь более высокий электроэндосмотический поток. Поэтому агароза с низким EEO обычно предпочтительна для использования в агарозе. гель-электрофорез нуклеиновых кислот, но агароза с высоким EEO может использоваться для других целей. Более низкое содержание сульфата в агарозе с низким EEO, особенно в агарозе с низкой температурой плавления (LMP), также полезно в тех случаях, когда ДНК, выделенная из геля, должна использоваться для дальнейших манипуляций, поскольку присутствие загрязняющих сульфатов может повлиять на некоторые последующие процедуры, такие как в качестве перевязка и ПЦР. Однако агарозы с нулевым ЭЭО нежелательны для некоторых применений, поскольку они могут быть получены путем добавления положительно заряженных групп, и такие группы могут влиять на последующие ферментативные реакции.[9] Электроэндосмос - причина, по которой агароза используется вместо агар как агаропектин Компонент в агаре содержит значительное количество отрицательно заряженных сульфатных и карбоксильных групп. Удаление агаропектина в агарозе существенно снижает EEO, а также снижает неспецифическую адсорбцию биомолекул на гелевой матрице. Однако для некоторых применений, таких как электрофорез белков сыворотки, может быть желательным высокий EEO, и в используемый гель можно добавлять агаропектин.[10]
Миграция нуклеиновых кислот в агарозном геле
Факторы, влияющие на миграцию нуклеиновой кислоты в геле
На миграцию нуклеиновых кислот может влиять ряд факторов: размер пор геля (концентрация геля), размер ДНК, подвергаемой электрофорезу, используемое напряжение, ионная сила буфера и концентрация интеркалирующего красителя, такого как бромид этидия. если используется во время электрофореза.[11]
Молекулы меньшего размера перемещаются в геле быстрее, чем молекулы большего размера, а двухцепочечная ДНК движется со скоростью, которая обратно пропорциональна логарифму числа пар оснований. Однако эта взаимосвязь нарушается с очень большими фрагментами ДНК, и разделение очень больших фрагментов ДНК требует использования гель-электрофорез в импульсном поле (PFGE), который применяет переменный ток с двух разных направлений, и большие фрагменты ДНК разделяются, поскольку они переориентируются с изменяющимся током.[12]
Для стандартного электрофореза в агарозном геле более крупные молекулы лучше разделяются при использовании геля с низкой концентрацией, в то время как молекулы меньшего размера лучше разделяются при использовании геля с высокой концентрацией. Однако гель с высокой концентрацией требует более длительного времени работы (иногда дней).
На движение ДНК может влиять конформация молекулы ДНК, например, сверхспиральная ДНК обычно движется быстрее, чем расслабленная ДНК, потому что она плотно свернута и, следовательно, более компактна. В препарате нормальной плазмидной ДНК может присутствовать несколько форм ДНК.[13] Гель-электрофорез плазмид обычно показывает отрицательно свернутую форму в качестве основной полосы, в то время как разорванная ДНК (открытая круглая форма) и расслабленная закрытая круговая форма проявляются как второстепенные полосы. Однако скорость, с которой перемещаются различные формы, может изменяться при различных условиях электрофореза.[14] и на подвижность более крупной кольцевой ДНК может сильнее влиять размер пор геля, чем на линейную ДНК.[15]
Бромид этидия, который внедряется в кольцевую ДНК, может изменять заряд, длину, а также суперсильность молекулы ДНК, поэтому его присутствие в геле во время электрофореза может повлиять на ее движение. Например, положительный заряд бромистого этидия может уменьшить движение ДНК на 15%.[12] Электрофорез в агарозном геле можно использовать для разделения кольцевой ДНК с различной топологией суперспирализации.[16]
Повреждение ДНК из-за повышенного сшивание также уменьшит электрофоретическую миграцию ДНК дозозависимым образом.[17][18]
Скорость миграции ДНК пропорциональна приложенному напряжению, то есть чем выше напряжение, тем быстрее движется ДНК. Однако разрешение больших фрагментов ДНК ниже при высоком напряжении. Подвижность ДНК также может изменяться в нестабильном поле - в поле, которое периодически меняется, подвижность ДНК определенного размера может значительно снижаться при определенной частоте циклов.[4] Это явление может привести к инверсии полосы при гель-электрофорезе с инверсией поля (FIGE), в результате чего более крупные фрагменты ДНК перемещаются быстрее, чем более мелкие.
Миграционные аномалии
- Гели "Smiley" - этот краевой эффект возникает, когда приложенное напряжение слишком велико для используемой концентрации геля.[19]
- Перегрузка ДНК - перегрузка ДНК замедляет миграцию фрагментов ДНК.
- Загрязнение - присутствие примесей, таких как соли или белки, может повлиять на движение ДНК.
Механизм миграции и отделения
Отрицательный заряд его фосфатного скелета перемещает ДНК к положительно заряженному аноду во время электрофореза. Однако миграция молекул ДНК в растворе в отсутствие гелевой матрицы не зависит от молекулярной массы во время электрофореза.[4][20] Таким образом, гелевая матрица отвечает за разделение ДНК по размеру во время электрофореза, и существует ряд моделей для объяснения механизма разделения биомолекул в гелевой матрице. Широко распространена модель Огстона, в которой полимерная матрица рассматривается как сито. Глобулярный белок или случайный катушки ДНК движется через соединенные между собой поры, и движение более крупных молекул с большей вероятностью будет затруднено и замедлено из-за столкновений с гелевой матрицей, поэтому молекулы разных размеров могут быть разделены в этом процессе просеивания.[4]
Однако модель Огстона не работает для больших молекул, в результате чего поры значительно меньше размера молекулы. Для молекул ДНК размером более 1 т.п.н. отказ модель (или ее варианты) наиболее часто используются. Эта модель предполагает, что ДНК может ползать «змеиным» образом (отсюда и «рептация») через поры в виде удлиненной молекулы. Модель смещения рептаций применяется при более высокой напряженности электрического поля, в результате чего передний конец молекулы становится сильно смещенным в прямом направлении и тянет за собой остальную часть молекулы.[21] Однако флуоресцентная микроскопия окрашенных молекул в реальном времени показала более тонкую динамику во время электрофореза, при этом ДНК демонстрирует значительную эластичность, поскольку она попеременно растягивается в направлении приложенного поля, а затем сжимается в шар или зацепляется в U-образную форму. когда он зацепляется за полимерные волокна.[22][23]
Общая процедура
Детали эксперимента с электрофорезом в агарозном геле могут варьироваться в зависимости от методов, но большинство из них следует общей процедуре.

Отливка геля

Гель готовят растворением порошка агарозы в подходящем буфере, таком как ТАЕ или ТВЕ, для использования в электрофорезе.[24] Агарозу диспергируют в буфере перед нагреванием до температуры, близкой к температуре кипения, но избегайте кипения. Расплавленной агарозе дают остыть в достаточной степени, прежде чем заливать раствор в гипс, так как он может деформироваться или треснуть, если раствор агарозы слишком горячий. В гипс помещается гребешок для создания лунок для загрузки образца, и гель должен полностью затвердеть перед использованием.
Концентрация геля влияет на разрешение разделения ДНК. Гель агарозы состоит из микроскопических пор, через которые проходят молекулы, и существует обратная зависимость между размером пор геля агарозы и его концентрацией - размер пор уменьшается по мере увеличения плотности волокон агарозы. Высокая концентрация геля улучшает разделение более мелких молекул ДНК, а снижение концентрации геля позволяет разделять большие молекулы ДНК. Этот процесс позволяет разделять фрагменты от 50 пар оснований до нескольких мега оснований в зависимости от используемой концентрации геля.[25] Концентрация измеряется в соотношении веса агарозы к объему использованного буфера (г / мл). Для стандартного электрофореза в агарозном геле 0,8% гель дает хорошее разделение или разрешение больших фрагментов ДНК размером 5–10 килобайт, а 2% гель дает хорошее разрешение для небольших фрагментов размером 0,2–1 килобайт. Для стандартного электрофореза часто используют 1% гели.[26] Гели с высоким процентным содержанием часто бывают хрупкими и не могут схватываться равномерно, в то время как гели с низким процентным содержанием (0,1-0,2%) являются хрупкими и с ними нелегко обращаться. Гели агарозы с низкой температурой плавления (LMP) также более хрупкие, чем гель нормальной агарозы. Агароза с низкой температурой плавления может использоваться отдельно или одновременно со стандартной агарозой для разделения и выделения ДНК.[27] PFGE и FIGE часто выполняются с использованием гелей с высоким содержанием агарозы.
Загрузка образцов
Как только гель затвердеет, гребенку удаляют, оставляя лунки, куда можно загрузить образцы ДНК. Буфер для загрузки смешивают с образцом ДНК перед загрузкой смеси в лунки. Загрузочный буфер содержит плотное соединение, которое может быть глицерином, сахарозой или Фиколл, что увеличивает плотность образца, так что образец ДНК может опуститься на дно лунки.[28] Если после приготовления образец ДНК содержит остаточный этанол, он может выплыть из лунки. Буфер загрузки также включает цветные красители, такие как ксилол цианол и бромфеноловый синий используется для наблюдения за ходом электрофореза. Образцы ДНК загружаются с помощью пипетка.
Электрофорез

Электрофорез в агарозном геле чаще всего проводят горизонтально в подводном режиме, когда пластинчатый гель полностью погружается в буфер во время электрофореза. Также возможно, но реже, проводить электрофорез как в вертикальном, так и в горизонтальном положении с гелем, приподнятым на ножках агарозы, с использованием подходящего устройства.[29] Буфер, используемый в геле, такой же, как рабочий буфер в резервуаре для электрофореза, поэтому электрофорез в подводном режиме возможен с гелем агарозы.
Для оптимального разрешения ДНК более 2 kb в стандартном гель-электрофорезе рекомендуется от 5 до 8 В / см (расстояние в см относится к расстоянию между электродами, поэтому это рекомендуемое напряжение должно быть от 5 до 8, умноженное на расстояние между электродами в см).[14] Напряжение также может быть ограничено тем фактом, что он нагревает гель и может вызвать плавление геля, если он работает под высоким напряжением в течение длительного периода, особенно если используемый гель представляет собой гель агарозы LMP. Слишком высокое напряжение может также снизить разрешение, а также вызвать появление полос для больших молекул ДНК. Слишком низкое напряжение может привести к расширению полосы для небольших фрагментов ДНК из-за дисперсии и диффузии.[30]
Поскольку ДНК не видна при естественном свете, за ходом электрофореза следят с помощью цветных красителей. Ксилолцианол (голубой цвет) объединяет большие фрагменты ДНК, в то время как бромфеноловый синий (темно-синий) объединяет более мелкие фрагменты. К менее часто используемым красителям относятся: Крезол Красный и Оранжевый G которые мигрируют впереди бромфенолового синего. А Маркер ДНК также выполняется вместе для оценки молекулярной массы фрагментов ДНК. Обратите внимание, однако, что размер кольцевой ДНК-подобной плазмиды нельзя точно измерить с помощью стандартных маркеров, если он не был линеаризован с помощью ограничительный дайджест в качестве альтернативы можно использовать маркер суперспиральной ДНК.
Окрашивание и визуализация

ДНК, а также РНК обычно визуализируются путем окрашивания этидиум бромид, который внедряется в основные бороздки ДНК и флуоресцирует в УФ-свете. Интеркаляция зависит от концентрации ДНК, и, таким образом, полоса с высокой интенсивностью будет указывать на большее количество ДНК по сравнению с полосой с меньшей интенсивностью.[12] Бромид этидия может быть добавлен к раствору агарозы до образования геля, или гель ДНК может быть окрашен позже после электрофореза. Обесцвечивание геля не обязательно, но может улучшить изображение. Доступны другие методы окрашивания; примеры SYBR Зеленый, GelRed, метиленовый синий, блестящий крезиловый синий, Нильский синий сульфат и кристально-фиолетовый.[31] SYBR Green, GelRed и другие аналогичные коммерческие продукты продаются как более безопасные альтернативы бромистому этидию, поскольку, как было показано, он мутагенный в Тест Эймса, Хотя канцерогенность бромистого этидия фактически не установлено. SYBR Green требует использования трансиллюминатора синего света. ДНК, окрашенную кристаллическим фиолетовым, можно рассматривать при естественном освещении без использования УФ-трансиллюминатора, что является преимуществом, однако он может не давать четкой полосы.
При окрашивании бромистым этидием гель просматривается с ультрафиолетовый (УФ) трансиллюминатор. УФ-свет возбуждает электроны в ароматическом кольце бромистого этидия, и как только они возвращаются в основное состояние, высвобождается свет, заставляя ДНК и комплекс бромида этидия флуоресцировать.[12] Стандартные трансиллюминаторы используют длины волн 302/312 нм (УФ-В), однако воздействие УФ-излучения на ДНК в течение всего 45 секунд может привести к повреждению ДНК и повлиять на последующие процедуры, например, снизить эффективность трансформация, in vitro транскрипция, и ПЦР.[32] Следовательно, следует ограничить воздействие УФ-излучения на ДНК. Использование более высокой длины волны 365 нм (диапазон УФ-А) вызывает меньшее повреждение ДНК, но также дает гораздо более слабую флуоресценцию с бромидом этидия. Если в трансиллюминторе можно выбрать несколько длин волн, более короткая длина волны будет использоваться для захвата изображений, тогда как более длинная длина волны должна использоваться, если необходимо работать с гелем в течение любого продолжительного периода времени.
Устройство трансиллюминатора может также содержать устройства захвата изображения, такие как цифровая или поляроидная камера, которые позволяют снимать или печатать изображение геля.
Для гель-электрофореза белка полосы можно визуализировать с помощью Кумасси или же серебряные пятна.

Последующие процедуры
Разделенные полосы ДНК часто используют для дальнейших процедур, и полосу ДНК можно вырезать из геля в виде среза, растворить и очистить. Однако загрязняющие вещества могут влиять на некоторые последующие процедуры, такие как ПЦР, и в некоторых случаях может быть предпочтительна агароза с низкой температурой плавления, поскольку она содержит меньше сульфатов, которые могут влиять на некоторые ферментативные реакции. Гели также можно использовать для блоттинга.
Буферы
В общем, идеальный буфер должен иметь хорошую проводимость, выделять меньше тепла и иметь долгий срок службы.[33] Для электрофореза в агарозе используется ряд буферов; общие для нуклеиновых кислот включают Трис / Ацетат / ЭДТА (TAE) и Трис / борат / ЭДТА (TBE). Используемые буферы содержат ЭДТА для инактивации многих нуклеаз, для функционирования которых требуется двухвалентный катион. Борат в буфере TBE может быть проблематичным, поскольку борат может полимеризоваться и / или взаимодействовать с цис-диолами, такими как те, которые содержатся в РНК. TAE имеет самую низкую буферную емкость, но обеспечивает лучшее разрешение для ДНК большего размера. Это означает более низкое напряжение и больше времени, но более качественный продукт.
Было предложено много других буферов, например борат лития (LB), изоэлектрический гистидин, буферы для согласованных товаров pK и т.д .; в большинстве случаев предполагаемое объяснение - более низкий ток (меньше тепла) и / или согласованная подвижность ионов, что приводит к увеличению срока службы буфера. Трис-фосфатный буфер обладает высокой буферной способностью, но его нельзя использовать, если экстрагированная ДНК будет использоваться в реакции, чувствительной к фосфату. LB является относительно новым и неэффективен при разрешении фрагментов размером более 5 кб; Однако из-за его низкой проводимости можно было использовать гораздо более высокое напряжение (до 35 В / см), что означает более короткое время анализа для обычного электрофореза. Разница в размере всего одной пары оснований может быть устранена в 3% агарозном геле со средой с чрезвычайно низкой проводимостью (1 мМ борат лития).[34]
Другая буферная система может быть использована в конкретных приложениях, например, барбитуровая кислота-барбитурат натрия или трис-барбитурат Буферы можно использовать для электрофореза белков в агарозном геле, например, для обнаружения аномального распределения белков.[35]
Приложения
- Оценка размера молекул ДНК после переваривания рестрикционные ферменты, например, в отображение ограничений клонированной ДНК.
- Анализ продукции полимеразной цепной реакции (ПЦР), например, в молекулярных генетическая диагностика или же генетическая дактилоскопия
- Разделение фрагментов ДНК для экстракции и очистки.
- Разделение рестриктированной геномной ДНК до Южный трансфер, или РНК до Северный трансфер.
- Разделение белков, например, скрининг аномалий белков в клиническая химия.[36]
Гели агарозы легко отливаются и обрабатываются по сравнению с другими матрицами, а нуклеиновые кислоты химически не изменяются во время электрофореза. Образцы также легко восстанавливаются. После окончания эксперимента полученный гель можно хранить в полиэтиленовом пакете в холодильнике.
Электрофорез проводится в буферных растворах, чтобы уменьшить изменения pH из-за электрического поля, что важно, потому что заряд ДНК и РНК зависит от pH, но слишком долгая работа может истощить буферную способность раствора. Кроме того, разные препараты генетического материала могут не мигрировать согласованно друг с другом по морфологическим или другим причинам.
Смотрите также
- Гель-электрофорез
- Иммунодиффузия, Иммуноэлектрофорез
- SDD-AGE
- Нозерн-блот
- Электрофорез в SDS-полиакриламидном геле
- Саузерн-блот
Рекомендации
- ^ Крындушкин Д.С., Александров И.М., Тер-Аванесян М.Д., Кушниров В.В. (декабрь 2003 г.). «Агрегаты прионов [PSI +] дрожжей образованы небольшими полимерами Sup35, фрагментированными Hsp104». Журнал биологической химии. 278 (49): 49636–43. Дои:10.1074 / jbc.M307996200. PMID 14507919.
- ^ Сэмбрук Дж., Рассел Д.В. (2001). Молекулярное клонирование: лабораторное руководство, 3-е изд. Лабораторный пресс Колд-Спринг-Харбор. Колд-Спринг-Харбор, Нью-Йорк.
- ^ Джозеф Сэмбрук; Дэвид Рассел. «Глава 5, протокол 1». Молекулярное клонирование - лабораторное руководство. 1 (3-е изд.). п. 5.4. ISBN 978-0-87969-577-4.
- ^ а б c d Zimm BH, Levene SD (май 1992 г.). «Проблемы и перспективы теории гель-электрофореза ДНК» (PDF). Ежеквартальные обзоры биофизики. 25 (2): 171–204. Дои:10,1017 / с0033583500004662. PMID 1518924.
- ^ Жан-Луи Виови (2000). «Электрофорез ДНК и других полиэлектролитов: физические механизмы». Обзоры современной физики. 72 (3): 813–872. Bibcode:2000РвМП ... 72..813В. Дои:10.1103 / RevModPhys.72.813.
- ^ а б Филип Сервер (1983). «Агарозные гели: свойства и применение для электрофореза». Электрофорез. 4 (6): 375–382. Дои:10.1002 / elps.1150040602. S2CID 97819634.
- ^ Джозеф Сэмбрук; Дэвид Рассел. «Глава 5, протокол 1». Молекулярное клонирование - лабораторное руководство. 1 (3-е изд.). п. 5.2–5.3. ISBN 978-0-87969-577-4.
- ^ «Приложение B: Физическая химия агарозы» (PDF). Lonza Group.
- ^ Джозеф Сэмбрук; Дэвид Рассел. «Глава 5, протокол 1». Молекулярное клонирование - лабораторное руководство. 1 (3-е изд.). п. 5.7. ISBN 978-0-87969-577-4.
- ^ Керен, Дэвид (26 сентября 2003 г.). Белковый электрофорез в клинической диагностике. CRC Press. С. 7–8. ISBN 978-0340812136.
- ^ Г. Люкотт; Ф. Банейкс (1993). Введение в методы молекулярного клонирования. Вили-Блэквелл. п. 32. ISBN 978-0471188490.
- ^ а б c d Ли PY, Costumbrado J, Hsu CY, Kim YH (апрель 2012 г.). «Электрофорез в агарозном геле для разделения фрагментов ДНК». Журнал визуализированных экспериментов (62). Дои:10.3791/3923. ЧВК 4846332. PMID 22546956.
- ^ Ричард Р. Синден (1994-11-24). Структура и функции ДНК. Academic Press Inc. стр. 97. ISBN 978-0126457506.
- ^ а б Джозеф Сэмбрук; Дэвид Рассел. «Глава 5, протокол 1». Молекулярное клонирование - лабораторное руководство. 1 (3-е изд.). п. 5.5–5.6. ISBN 978-0-87969-577-4.
- ^ Aaij C, Borst P (май 1972 г.). «Гель-электрофорез ДНК». Biochimica et Biophysica Acta (BBA) - нуклеиновые кислоты и синтез белков. 269 (2): 192–200. Дои:10.1016/0005-2787(72)90426-1. PMID 5063906.
- ^ Дональд Воет; Джудит Г. Воет (1995). Биохимия (2-е изд.). Джон Вили и сыновья. стр.877–878. ISBN 978-0471586517.
- ^ Blasiak J, Trzeciak A, Malecka-Panas E, Drzewoski J, Wojewódzka M (август 2000 г.). «Генотоксичность этанола и ацетальдегида in vitro в лимфоцитах человека и клетках слизистой оболочки желудочно-кишечного тракта». Токсикология in vitro. 14 (4): 287–95. Дои:10.1016 / S0887-2333 (00) 00022-9. PMID 10906435.
- ^ Лу И, Моримото К. (июль 2009 г.). «Связано ли обычное употребление алкоголя со снижением миграции электрофоретической ДНК в лейкоцитах периферической крови у мужчин японцев с дефицитом ALDH2?». Мутагенез. 24 (4): 303–8. Дои:10.1093 / mutage / gep008. PMID 19286920.
- ^ Г. Люкотт; Ф. Банейкс (1993). Введение в методы молекулярного клонирования. Вили-Блэквелл. п. 41. ISBN 978-0471188490.
- ^ Роберт В. Олд; Сэнди Б. Примроуз (27 сентября 1994). Принцип манипуляции генами - введение в генную инженерию (5-е изд.). Blackwell Scientific. п.9. ISBN 9780632037124.
- ^ Ли Чжу; Хун Ван (2009-03-02). «Глава 4 - Генетический анализ в миниатюрных системах электрофореза». Инь Тянь, Вэй-Чэн; Finehout, Эрин (ред.). Микрофлюидика для биологических приложений. Springer. п. 125. ISBN 978-0-387-09480-9.
- ^ Смит С.Б., Олдридж П.К., Каллис Дж. Б. (январь 1989 г.). «Наблюдение за отдельными молекулами ДНК, подвергающимися гель-электрофорезу». Наука. 243 (4888): 203–6. Bibcode:1989Sci ... 243..203S. Дои:10.1126 / science.2911733. PMID 2911733.
- ^ Шварц Д.К., Коваль М. (апрель 1989 г.). «Конформационная динамика отдельных молекул ДНК при гель-электрофорезе». Природа. 338 (6215): 520–2. Bibcode:1989Натура.338..520С. Дои:10.1038 / 338520a0. PMID 2927511. S2CID 4249063.
- ^ Пей Юн Ли; Джон Костумбрадо; Чжи-Юань Сюй; Ён Хун Ким (2012). «Электрофорез в агарозном геле для разделения фрагментов ДНК». Журнал визуализированных экспериментов (62): e3923. Дои:10.3791/3923. PMID 22546956.
- ^ Магдельдин, Самех (2012). Гель-электрофорез. InTech. С. 35–40.
- ^ «Электрофорез в агарозном геле (основной метод)». Биологические протоколы. Получено 23 августа 2011.
- ^ Фотадар У., Шапиро Л.Е., Суркс М.И. (февраль 1991 г.). «Одновременное использование стандартной и легкоплавкой агарозы для разделения и выделения ДНК электрофорезом». Биотехнологии. 10 (2): 171–2. PMID 2059440.
- ^ Ли PY, Costumbrado J, Hsu CY, Kim YH (апрель 2012 г.). «Электрофорез в агарозном геле для разделения фрагментов ДНК». Журнал визуализированных экспериментов. 62 (62). Дои:10.3791/3923. ЧВК 4846332. PMID 22546956.
- ^ Дэвид Фрайфельдер (1982). Физическая биохимия: приложения к биохимии и молекулярной биологии (2-е изд.). WH Freeman. С. 292–293. ISBN 978-0716714446.
- ^ «Раздел III: Загрузка и запуск ДНК в агарозных гелях» (PDF). Lonza Group.
- ^ "ДНК раскрыта" (PDF). Национальный центр биотехнологического образования. Университет Ридинга. Архивировано из оригинал (PDF) на 2012-03-04.
- ^ Грюндеманн Д., Шёмиг Э. (ноябрь 1996 г.). «Защита ДНК во время препаративного электрофореза в агарозном геле от повреждений, вызванных ультрафиолетом» (PDF). Биотехнологии. 21 (5): 898–903. Дои:10.2144 / 96215rr02. PMID 8922632. Архивировано из оригинал (PDF) на 2016-03-04. Получено 2013-07-03.
- ^ Самех Магдельдин, изд. (2012). Гель-электрофорез - принципы и основы. InTech. ISBN 978-953-51-0458-2.
- ^ Броуди-младший, Керн С.Е. (октябрь 2004 г.). «История и принципы проводящих сред для стандартного электрофореза ДНК» (PDF). Аналитическая биохимия. 333 (1): 1–13. Дои:10.1016 / j.ab.2004.05.054. PMID 15351274. Архивировано из оригинал (PDF) 24 декабря 2012 г.
- ^ Jeppsson JO, Laurell CB, Franzén B (апрель 1979 г.). «Электрофорез в агарозном геле». Клиническая химия. 25 (4): 629–38. Дои:10.1093 / Clinchem / 25.4.629. PMID 313856.
- ^ Джо, Жан-Франсуа (2010). «Электрофорез в агарозном геле: применение в клинической химии» (PDF). Журнал медицинской биохимии. 29: 9–14. Дои:10.2478 / v10011-009-0033-8. S2CID 94646249.
внешняя ссылка
- Как запустить гель ДНК или РНК
- Анимация гель-анализа рестрикционных фрагментов ДНК
- Видео и статья по электрофорезу в агарозном геле
- Пошаговые фотографии запуска геля и извлечения ДНК
- Электрофорез на соломинке!
- Типичный метод из викиверситета
- Создание камеры для гель-электрофореза