Транс-действующая миРНК - Trans-acting siRNA
Транс-активная миРНК (сокращенно "та-миРНК" или "tasiRNA") являются классом малая интерферирующая РНК (миРНК), которые подавляют экспрессия гена через посттранскрипционное молчание генов в наземные растения.[1][2][3] Транскрипты прекурсоров из ТАС локусы полиаденилированный и превращаются в двухцепочечную РНК, а затем процессируются в дуплексы РНК длиной 21 нуклеотид с выступами.[1] Эти сегменты включены в РНК-индуцированный комплекс сайленсинга (RISC) и направляют специфичное для последовательности расщепление мРНК-мишени. Ta-миРНК классифицируются как миРНК, потому что они возникают из двухцепочечная РНК (дцРНК).[4]
Открытие
ta-siRNA были впервые обнаружены в 2004 году в цветковых растениях. Arabidopsis thaliana.[1][2] Первоначальные описания обнаружили участие растительного белка, супрессора сайленсинга гена 3 (SGS3), и фермента РНК-зависимой РНК-полимеразы 6 (RDR6).
Биосинтез
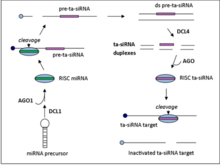
Ta-миРНК генерируются из некодирующих транскриптов посредством Аргонавт -опосредованный miRNA управляемое расщепление с последующим преобразованием в двухцепочечную РНК с помощью RDR6.[5] Полученная дцРНК далее процессируется дайсер-подобным ферментом 4 (DCL4) для получения фазированной матрицы из 21 нуклеотида. миРНК из позиций, примыкающих к сайту расщепления miRNA.[6]
Существует четыре семейства локусов, генерирующих та-миРНК (ТАС гены) в A. thaliana. Семейства TAS1, TAS2 и TAS4 требуют одного сайта связывания miRNA для того, чтобы произошло расщепление, тогда как TAS3 требует двух сайтов связывания.[7]Номера семейств генов TAS обычно не указывают ортология, например мох TAS1 семейство генов не имеет общего гена предка с Arabidopsis thaliana TAS1 генная семья.
TAS1 и TAS2
Транскрипты TAS1 / 2 подвергаются начальному AGO1-опосредованному расщеплению на 5 ’конце, которое управляется miR173. Затем RDR6 преобразует транскрипт в двухцепочечный фрагмент РНК, который затем обрабатывается DCL4 с образованием 21-нуклеотидной миРНК с 2'-нуклеотидными выступами, которые нацелены на комплементарные мРНК в транс.[7]
TAS4
Начальные шаги для семейства та-миРНК TAS4 аналогичны таковым для TAS1 и TAS2. Семейство транскриптов TAS4 сначала подвергается управляемому miR828, опосредованному AGO1 расщеплению с последующим синтезом дцРНК и процессингом с помощью DCL4.[7]
TAS3
В отличие от семейства связывания одиночной мРНК, TAS3 требует, чтобы направляющая мРНК miR390 связала транскрипт в двух сайтах. Затем транскрипт расщепляется только по 3 ’сайту связывания с помощью AGO7. Как и в случае семейств TAS1, TAS2 и TAS3, RDR6 затем синтезирует фрагмент дцРНК, который далее обрабатывается DCL4.[7]
Механизм
Эндогенные та-миРНК действуют посредством гетеро-сайленсинга, что означает, что гены, на которые они нацелены для расщепления и репрессии, не имеют большого сходства с генами, от которых происходят миРНК. Это отличается от других эндогенных миРНК которые являются цис-действующими и осуществляют автоматическое замалчивание, подавляя экспрессию генов, которые являются такими же или имеют большое сходство с генами, от которых они происходят. Ранее считалось, что только миРНК проявляют гетеро-сайленсинг.[1] Как и другие миРНК, та-миРНК включены в РНК-индуцированные комплексы сайленсинга (RISC), где они направляют комплекс на расщепление целевых мРНК в середине единственного комплементарного сайта и репрессию трансляции.[1][2][8]
Член Аргонавт Семейство белков является компонентом всех эффекторных комплексов, подавляющих молчание РНК, включая RISC, которые катализируют расщепление мРНК.[8][9] В частности, у арабидопсиса, по-видимому, AGO7 / ZIPPY играет роль в пути та-миРНК, действуя во время TAS3-производная ta-siRNA-опосредованная регуляция. AGO7 / ZIPPY не играет роли в механизмах TAS1 или TAS2 Биогенез та-миРНК.[9] ta-siRNA могут быть загружены в комплексы AGO1 для управления расщеплением целевой мРНК.[10]
Наличие в растениях
Помимо присутствия в A. thaliana,[6] доказательства ta-siRNA были также обнаружены во мхе. Physcomitrella patens,[5] кукуруза,[11] Oryza sativa (рис),[12] и другие растения. TAS3 транс-действующая коротко интерферирующая РНК, нацеленная на факторы ответа на ауксин («tasiR-ARF»), является примером та-миРНК, которая, как было показано, присутствует не только в арабидопсисе, но и во всех предыдущих примерах. TasiR-ARF отвечает за регулирование сигнальной молекулы ауксин. Это достигается путем воздействия на мРНК, кодирующую несколько генов фактора ауксинового ответа (ARF), для деградации.[11]
Рекомендации
- ^ а б c d е Васкес Ф., Вошерет Х., Раджагопалан Р., Леперс С., Гашиолли В., Мэллори А.С., Гильберт Дж.Л., Бартель Д.П., Крете П. (октябрь 2004 г.). «Эндогенные транс-действующие миРНК регулируют накопление мРНК Arabidopsis». Мол. Клетка. 16 (1): 69–79. Дои:10.1016 / j.molcel.2004.09.028. PMID 15469823.
- ^ а б c Перагин А., Йошикава М., Ву Г., Альбрехт Х.Л., Поэтиг Р.С. (октябрь 2004 г.). «SGS3 и SGS2 / SDE1 / RDR6 необходимы для развития молоди и продукции транс-действующих миРНК у Arabidopsis». Genes Dev. 18 (19): 2368–79. Дои:10.1101 / gad.1231804. ЧВК 522987. PMID 15466488.
- ^ Axtell MJ, Ян К., Раджагопалан Р., Бартель Д.П. (ноябрь 2006 г.). «Триггер с двумя ударами для биогенеза siRNA в растениях». Клетка. 127 (3): 565–77. Дои:10.1016 / j.cell.2006.09.032. PMID 17081978.
- ^ Акстелл, Майкл Дж. (29 апреля 2013 г.). «Классификация и сравнение малых РНК из растений». Ежегодный обзор биологии растений. 64 (1): 137–159. Дои:10.1146 / annurev-arplant-050312-120043.
- ^ а б Талмор-Нейман М., Став Р., Клипкан Л., Буксдорф К., Баулкомб, округ Колумбия, Арази Т. (ноябрь 2006 г.). «Идентификация транс-действующих миРНК в мхах и РНК-зависимой РНК-полимеразы, необходимой для их биогенеза». Завод J. 48 (4): 511–21. Дои:10.1111 / j.1365-313X.2006.02895.x. PMID 17076803.
- ^ а б Аллен Э., Се З., Густафсон А.М., Кэррингтон Дж.С. (апрель 2005 г.). «МикроРНК-направленная фазировка во время транс-действующего биогенеза миРНК в растениях». Клетка. 121 (2): 207–21. Дои:10.1016 / j.cell.2005.04.004. PMID 15851028.
- ^ а б c d Аллен Э., Доктор медицины Хауэлл (октябрь 2010 г.). «миРНК в биогенезе транс-действующих миРНК у высших растений». Семин. Cell Dev. Биол. 21 (8): 798–804. Дои:10.1016 / j.semcdb.2010.03.008. PMID 20359543.
- ^ а б Томари Ю., Заморе П.Д. (март 2005 г.). «Перспектива: машины для РНКи». Genes Dev. 19 (5): 517–29. Дои:10.1101 / gad.1284105. PMID 15741316.
- ^ а б Адено X, Эльмаян Т., Лауссерг Д., Буте С., Буше Н., Гашиолли В., Вошере Х. (май 2006 г.). "DRB4-зависимые транс-действующие миРНК TAS3 контролируют морфологию листа посредством AGO7". Curr. Биол. 16 (9): 927–32. Дои:10.1016 / j.cub.2006.03.035. PMID 16682354.
- ^ У Л., Мао Л., Ци И (октябрь 2012 г.). «Роль dicer-like и argonaute белков в TAS-производном малой интерферирующей РНК-триггерной ДНК метилировании». Физиология растений. 160 (2): 990–9. Дои:10.1104 / стр.112.200279. ЧВК 3461571. PMID 22846193.
- ^ а б Уильямс Л., Карлес СС, Осмонт К.С., Флетчер Дж.С. (июль 2005 г.). «Метод анализа базы данных идентифицирует эндогенную транс-действующую короткую интерферирующую РНК, которая нацелена на гены ARF2, ARF3 и ARF4 арабидопсиса». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 102 (27): 9703–8. Дои:10.1073 / pnas.0504029102. ЧВК 1172271. PMID 15980147.
- ^ Heisel SE, Zhang Y, Allen E, Guo L, Reynolds TL, Yang X, Kovalic D, Roberts JK (2008). «Характеристика уникальных популяций малых РНК из рисового зерна». PLoS ONE. 3 (8): e2871. Дои:10.1371 / journal.pone.0002871. ЧВК 2518513. PMID 18716673.
Виды нуклеиновые кислоты | |||||||
|---|---|---|---|---|---|---|---|
| Избиратели | |||||||
| Рибонуклеиновые кислоты (кодирование, некодирование ) |
| ||||||
| Дезоксирибонуклеиновые кислоты | |||||||
| Аналоги | |||||||
| Клонирование векторов | |||||||
| |||||||