Диметродон - Dimetrodon
| Диметродон | |
|---|---|
 | |
| Скелет D. limbatus, Staatliches Museum für Naturkunde Karlsruhe | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Семья: | †Sphenacodontidae |
| Подсемейство: | †Sphenacodontinae |
| Род: | †Диметродон Справиться, 1878 |
| Типовой вид | |
| †Clepsydrops limbatus Коп, 1877 г. | |
| Разновидность | |
Видеть ниже | |
| Синонимы | |
Синонимия рода
Синонимия видов
| |
Диметродон (/dаɪˈмятрədɒп/ (![]() Слушать)[1] или же /dаɪˈмɛтрədɒп/,[2] означает «две меры зубов») - это вымерший род не-млекопитающее синапсид которые жили во время Цисуральский (ранняя пермь) около 295–272 миллионов лет назад (млн лет назад).[3][4][5] Это член семьи Sphenacodontidae. Самая яркая особенность Диметродон большой нейронный позвоночник на его спине образованы удлиненными шипами, отходящими от позвонки. Это ходил на четырех ногах и имел высокий изогнутый череп с большими зубами разного размера, расположенными вдоль челюстей. Большинство окаменелостей было найдено на юго-западе Соединенных Штатов, большинство из них происходит из геологического месторождения под названием Красные кровати Техаса и Оклахомы. Совсем недавно окаменелости были обнаружены в Германии. С тех пор было названо более десятка видов. род был впервые возведен в 1878 году.
Слушать)[1] или же /dаɪˈмɛтрədɒп/,[2] означает «две меры зубов») - это вымерший род не-млекопитающее синапсид которые жили во время Цисуральский (ранняя пермь) около 295–272 миллионов лет назад (млн лет назад).[3][4][5] Это член семьи Sphenacodontidae. Самая яркая особенность Диметродон большой нейронный позвоночник на его спине образованы удлиненными шипами, отходящими от позвонки. Это ходил на четырех ногах и имел высокий изогнутый череп с большими зубами разного размера, расположенными вдоль челюстей. Большинство окаменелостей было найдено на юго-западе Соединенных Штатов, большинство из них происходит из геологического месторождения под названием Красные кровати Техаса и Оклахомы. Совсем недавно окаменелости были обнаружены в Германии. С тех пор было названо более десятка видов. род был впервые возведен в 1878 году.
Диметродон часто ошибочно принимают за динозавр или как современник динозавров в популярной культуре, но он вымер за 40 миллионов лет до первого появления динозавров. По внешнему виду и физиологии похожи на рептилий, Диметродон тем не менее, он более тесно связан с млекопитающими, чем с современными рептилиями, хотя и не является прямым предком млекопитающих.[4] Диметродон относится к категории "не млекопитающих" синапсиды ", группа, традиционно называемая" млекопитающими рептилиями ".[4] Эта группа Диметродон вместе с млекопитающими в клады (эволюционная группа) под названием Synapsida, при размещении динозавры, рептилии и птицы в отдельной кладе, Sauropsida. Отдельные отверстия в черепе за каждым глазом, известные как височные отверстия, и другие особенности черепа различают Диметродон и млекопитающие с самых ранних завропсиды.
Диметродон вероятно был одним из высшие хищники экосистем Приуралья, питаясь рыбой и четвероногие, включая рептилий и амфибии. Меньше Диметродон виды могли иметь разные экологические роли. Парус Диметродон могли использоваться для стабилизации его позвоночника или для нагрева и охлаждения его тела в качестве формы терморегуляция. Некоторые недавние исследования[нужна цитата ] утверждают, что парус был бы неэффективен при отводе тепла от тела из-за того, что крупные виды были обнаружены с маленькими парусами, а мелкие виды были обнаружены с большими парусами, что по существу исключает регулирование тепла в качестве своей основной цели. Парус, скорее всего, использовался в дисплей ухаживания с такими методами, как угроза соперникам или хвастовство перед потенциальными партнерами.
Описание

Диметродон был четвероногий, парусный синапсид. Наиболее Диметродон виды имели длину от 1,7 до 4,6 метра (от 6 до 15 футов) и, по оценкам, весили от 28 до 250 килограммов (60 и 550 фунтов).[6] Самый крупный из известных видов Диметродон является D. angelensis около 4 метров (13 футов), а самый маленький D. teutonis на 60 см (24 дюйма).[7][8] Более крупные виды Диметродон были одними из крупнейших хищников ранней перми, хотя близкородственные Таппенозавр, известный по фрагментам скелета в немного более молодых породах, возможно, был даже больше - примерно 5,5 метров (18 футов) в общей длине тела.[9][10] Хотя некоторые Диметродон виды могли вырасти очень большими, известно много молодых особей.[11]
Череп
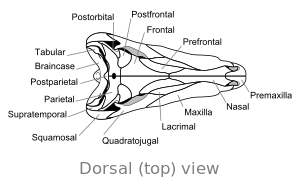 |  |
 |  |
А одно большое отверстие по обе стороны от задней части черепа звеньев Диметродон с млекопитающими и отличает его от большинства ранних сауропсид, которые либо не хватает отверстий или есть два отверстия. Такие особенности, как выступы на внутренней стороне носовая полость и гребень в задней части нижней челюсти считается частью эволюционного прогресса с раннего четвероногие наземные позвоночные к млекопитающие.
Череп Диметродон высокий и сжатый сбоку, или из стороны в сторону. Глазницы расположены высоко и далеко позади черепа. Позади каждой глазницы находится одно отверстие, называемое подвисочное отверстие. Дополнительное отверстие в черепе, надвисочное окно, видно при взгляде сверху. Задняя часть черепа ( затылок ) ориентирована под небольшим углом вверх, что является общей чертой всех других ранних синапсиды.[12] Верхний край черепа наклонен вниз по выпуклой дуге к кончику морды. Кончик верхней челюсти, образованный предчелюстная кость кость, возвышается над частью челюсти, образованной верхняя челюсть кость, чтобы сформировать верхнечелюстную «ступеньку». На этом этапе диастема, или разрыв в зубном ряду. Его череп был более прочным, чем динозавр с.
Зубы
Размер зубов сильно варьируется по длине челюстей, что дает Диметродон его название, которое означает «две меры зуба» применительно к множеству маленьких и больших зубов.[13] Одна или две пары клыков (большие заостренные собачий -подобные зубы) отходят от верхней челюсти. Большие резцы также присутствуют на концах верхней и нижней челюстей, уходят корнями в предчелюстные кости и зубные кости. Маленькие зубы присутствуют вокруг «ступеньки» верхней челюсти и позади клыков, которые становятся меньше в глубине челюсти.[14]

Многие зубы имеют наибольшую ширину в средней части и сужаются ближе к челюстям, что придает им вид слезы. Каплевидные зубы уникальны для Диметродон и другие тесно связанные сфенакодонтиды, и помочь отличить их от других ранних синапсидов.[8] Как и во многих других ранних синапсиды, зубы большинства Диметродон по краям зубчатые.[8] Зубцы Диметродон зубы были такими прекрасными, что напоминали крошечные трещинки.[15] Динозавр Альбертозавр имел аналогичные трещиноподобные зубцы, но в основании каждого зубца был круглый пустота, который функционировал бы для распределения силы по большему площадь поверхности и предотвратить распространение трещины по зубу из-за стресса от кормления. В отличие от Альбертозавр, Диметродон у зубов не было приспособлений, которые предотвращали бы образование трещин на их зубцах.[15] Зубы D. teutonis без зубцов, но все же имеют острые края.[8]
Исследование 2014 года показывает, что Диметродон был в гонке вооружений против своей добычи. Меньшие виды, Д. Миллери, не имел зубцов, так как ел мелкую добычу. По мере того, как добыча росла, несколько Диметродон виды начали развивать зубцы на зубах и увеличиваться в размерах. Например, D. limbatus имел эмалевые зубцы, которые помогали ему прорезать плоть (которые были похожи на зубцы, которые можно найти на Секодонтозавр ). Второй по величине вид, D. grandis, имеет зубцы, похожие на зубцы акул и теропод динозавров, что делает их зубы еще более специализированными для разрезания плоти. Это исследование демонстрирует, что, поскольку ДиметродонДобыча росла, различные виды в ответ эволюционировали в более крупные размеры и развили все более острые зубы.[16]
Полость носа
На внутренней поверхности носового отдела черепа расположены гребни, называемые назотурбиналы, который мог поддерживать хрящ, увеличивавший площадь обонятельный эпителий, слой ткани, улавливающий запахи. Эти гребни намного меньше, чем у более поздних синапсиды из поздней перми и триаса, чьи большие носотурбиналы рассматриваются как свидетельство теплокровности, поскольку они, возможно, поддерживали слизистые оболочки, которые согревали и увлажняли поступающий воздух. Таким образом, полость носа Диметродон является переходный между теми из ранней земли позвоночные и млекопитающие.[17]
Челюстной сустав и ухо
Еще одна переходная особенность Диметродон это выступ в задней части челюсти, называемый отраженной пластиной. Отраженная пластинка находится на суставной кость, которая соединяется с квадратная кость черепа с образованием челюстного сустава. У более поздних предков млекопитающих суставная и квадратная части тела отделились от челюстного сустава, в то время как суставная часть превратилась в суставную. молоточек кость среднее ухо. Отраженная пластинка стала частью кольца, называемого барабанным кольцом, которое поддерживает барабанная перепонка у всех живых млекопитающих.[18]
Хвост

Хвост Диметродон составляет большую часть его общей длины тела и включает около 50 хвостовые позвонки. Хвосты отсутствовали или были неполными у первых описанных скелетов Диметродон; единственными известными хвостовыми позвонками были одиннадцать ближайших к бедру. Поскольку эти первые несколько хвостовых позвонков быстро сужаются по мере удаления от бедра, многие палеонтологи в конце девятнадцатого и начале двадцатого веков считали, что Диметродон имел очень короткий хвост. Лишь в 1927 г. Диметродон был описан.[19]
Плыть

Парус Диметродон образуется удлиненной нервные отростки выступающие из позвонков. Каждый шип варьируется по форме поперечного сечения от основания до кончика в так называемой дифференциации «диметродонта».[20] Рядом с телом позвонка поперечное сечение позвоночника сжато с боков до прямоугольной формы, а ближе к кончику принимает форму восьмерки, поскольку бороздка проходит по обеим сторонам позвоночника. Считается, что форма восьмерки укрепляет позвоночник, предотвращая сгибание и переломы.[21] Поперечный разрез позвоночника одного экземпляра Dimetrodon giganhomogenes имеет прямоугольную форму, но сохраняет кольца в форме восьмерки близко к центру, что указывает на то, что форма шипов может меняться с возрастом.[22] Микроскопическая анатомия каждого позвоночника варьируется от основания к кончику, что указывает на то, где он был встроен в мышцы спины, а где - как часть паруса. Нижняя или проксимальный часть позвоночника имеет шероховатую поверхность, которая служила бы точкой крепления для эпаксиальные мышцы спины, а также имеет сеть соединительных тканей, называемую Волокна Шарпея что указывает на то, что он был встроен в тело. Выше на дистальный (внешняя) часть позвоночника, поверхность кости более гладкая. В надкостница слой ткани, окружающий кость, покрыт небольшими бороздками, которые предположительно поддерживают кровеносные сосуды, васкуляризирующие парус.[23]
Когда-то считалось, что большая бороздка, проходящая по всей длине позвоночника, является каналом для кровеносных сосудов, но, поскольку кость не содержит сосудистых каналов, парус не считается столь сильно васкуляризованным, как считалось раньше. Некоторые образцы Диметродон сохранить деформированные участки позвоночника, которые выглядят как залеченные переломы. В кортикальная кость , который вырос через эти разрывы, сильно васкуляризован, что позволяет предположить, что мягкие ткани должны были присутствовать на парусе, чтобы снабжать это место кровеносный сосуд.[21] Слоистый пластинчатая кость составляет большую часть площади поперечного сечения нервного отдела позвоночника и содержит линии задержки роста, которые можно использовать для определения возраста каждого человека на момент смерти.[24] Во многих экземплярах D. gigashomogenes дистальные части шипов резко изгибаются, указывая на то, что при жизни парус имел неправильный профиль. Их изогнутость предполагает, что мягкие ткани, возможно, не простирались до кончиков хребтов, а это означает, что лямка паруса могла быть не такой обширной, как ее обычно представляют.[20]
Кожа

Нет ископаемых свидетельств Диметродоншкуру еще не нашли. Отпечатки кожи родственного животного, Estemmenosuchus, указывают на то, что она была бы гладкой и хорошо снабженной железами, однако эта форма кожи могла не применяться к Диметродон поскольку его происхождение довольно далекое.[25] Диметродон также могли иметь большие щитки на нижней стороне его хвоста и живота, как и другие синапсиды.[26][27] Свидетельства из варанопид Ascendonanus предполагает, что некоторые ранние синапсиды могли иметь чешуйчатый чешуйки.[28] Однако недавние исследования поставили варанопидов таксономически ближе к диапсид рептилии.[29][30]
История классификации
Первые описания Копа
Окаменелости теперь приписываются Диметродон были впервые изучены американским палеонтологом Эдвард Дринкер Коуп в 1870-х гг. Коп получил окаменелости вместе с окаменелостями многих других пермских четвероногие от нескольких коллекционеров, которые исследовали группу скал в Техасе, названную Красные Кровати. Среди этих коллекционеров были швейцарские натуралисты. Джейкоб Болл, Техасский геолог В. Ф. Камминс, и палеонтолог-любитель Чарльз Хазелиус Штернберг.[31] Большинство образцов Коупа отправились в Американский музей естественной истории или в Чикагский университет Уокера (большая часть коллекции окаменелостей Уокера сейчас находится в Полевой музей естественной истории ).
Штернберг отправил несколько своих образцов немецкому палеонтологу. Фердинанд Бройли в Мюнхенский университет, хотя Бройли не был таким плодовитым, как Коп, когда дело доходило до описания образцов. Соперник Коупа Отниэль Чарльз Марш также собрал некоторые кости Диметродон, который он отправил в Музей Уокера.[32] Первое использование имени Диметродон появился в 1878 году, когда Коуп назвал вид Диметродон инцизивус, Диметродон прямой, и Диметродон гигас в научном журнале Труды Американского философского общества.[33]
Однако первое описание Диметродон окаменелость появилась годом ранее, когда Коуп назвал вид Clepsydrops limbatus от Кровати Texas Red.[34] (Название Клепсидропс был впервые придуман Копом в 1875 году для сфенакодонтид остается от Округ Вермилион, Иллинойс, и позже был использован для многих образцов сфенаконтид из Техаса; многие новые виды сфенакодонтид из Техаса были отнесены к Клепсидропс или же Диметродон в конце девятнадцатого и начале двадцатого веков.) C. limbatus был реклассифицирован как вид Диметродон в 1940 году, а это означает, что статья Коупа 1877 года была первой записью Диметродон.
Коп был первым, кто описал парусный синапсид с наименованием Клепсидропс наталис в своей статье 1878 года, хотя он назвал парус плавником и сравнил его с гербами современных ящерица василиск (Basilicus). Паруса не сохранились у экземпляров D. incisivus и D. gigas что Коп описал в своей статье 1878 года, но удлиненные шипы присутствовали в D. rectiformis экземпляр, который он описал.[33] Коуп прокомментировал назначение паруса в 1886 году, написав: «Трудно представить себе полезность этого паруса. Если животное не имело водных привычек и не плавало на спине, гребень или плавник должны были мешать активным движениям ... Конечности недостаточно длинные, а когти недостаточно острые, чтобы демонстрировать древесные привычки, как у существующего рода. Basilicus, где есть подобный гребень ".[14]
Описания начала 20 века

В первые несколько десятилетий двадцатого века американский палеонтолог E. C. Дело автор многих исследований по Диметродон и описал несколько новых видов. Он получил финансирование от Институт Карнеги за изучение многих Диметродон экземпляры в коллекциях Американский музей естественной истории и несколько других музеев.[32] Многие из этих окаменелостей были собраны Копом, но не были подробно описаны, поскольку Коп был известен созданием новых видов на основе всего нескольких фрагментов костей.
С конца 1920-х гг. Палеонтолог. Альфред Ромер пересмотрел многие Диметродон экземпляров и назвал несколько новых видов. В 1940 году Ромер в соавторстве с Ллевеллин Айвор Прайс называется "Обзор пеликозавров", в котором вид Диметродон названные Коупом и Кейсом были переоценены.[35] Большинство названий видов, которые Ромер и Прайс считают действительными, используются и сегодня.[23]
Новые экземпляры
В течение десятилетий после монографии Ромера и Прайса многие Диметродон экземпляры описаны из населенных пунктов вне Техас и Оклахома. Первый был описан из Четыре угла регион Юта в 1966 году[36] и еще один был описан из Аризоны в 1969 году.[37] В 1975 году Олсон сообщил Диметродон материал из Огайо.[38] Новый вид Диметродон называется D. occidentalis (что означает "западный Диметродон") был назван в 1977 году из Нью-Мексико.[39] Образцы, найденные в Юте и Аризоне, вероятно, также принадлежат D. occidentalis.[40]
До этих открытий существовала теория о том, что Срединно-континентальный морской путь отделял то, что сейчас является Техасом и Оклахомой, от более западных земель в течение ранней перми, изолируя Диметродон в небольшой регион Северной Америки, в то время как меньший сфенакодонтид называл Sphenacodon доминировал в западной области. Хотя этот морской путь, вероятно, действительно существовал, открытие окаменелостей за пределами Техаса и Оклахомы показывает, что его протяженность была ограничена и что он не был эффективным препятствием для распространения Диметродон.[39][41]
В 2001 г. появился новый вид Диметродон называется D. teutonis был описан из местонахождения Бромакер в нижней перми в Тюрингенском лесу в Германии, расширяя географический ареал Диметродон за пределами Северной Америки впервые.[6]
Разновидность

20 разновидность из Диметродон были названы с род впервые был описан в 1878 году. Многие из них были синонимизированный с более старыми названными видами, а некоторые теперь принадлежат к разным родам.
Резюме
| Разновидность | Орган власти | Место расположения | Положение дел | Синонимы | Изображений |
|---|---|---|---|---|---|
| Dimetrodon angelensis | Олсон, 1962 год. | Техас | Действительный |  | |
| Dimetrodon borealis | Лейди, 1854 г. | Остров Принца Эдуарда | Действительный | Ранее известный как Батигнат бореалис |  |
| Dimetrodon booneorum | Ромер, 1937 год. | Техас | Действительный | ||
| Диметродон долловиан | Дело, 1907 год | Техас | Действительный | Эмболофор долловиан Коп, 1888 г. | |
| Диметродон гигахомоген | Дело, 1907 год | Техас | Действительный |  | |
| Диметродон большой | Ромер и Прайс, 1940 | Оклахома Техас | Действительный | Clepsydrops gigas Коп, 1878 г. Диметродон гигас Коп, 1878 г. Theropleura grandis Дело, 1907 год Батиглипт теодори Дело, 1911 год Диметродон максимальный Ромер 1936 |  |
| Dimetrodon kempae | Ромер, 1937 год. | Техас | Возможный номен дубиум | ||
| Диметродон лимбатус | Ромер и Прайс, 1940 | Оклахома Техас | Действительный | Clepsydrops limbatus Коп, 1877 г. Диметродон инцизивус Коп, 1878 г. Диметродон прямой Коп, 1878 г. Диметродон полурадикатус Коп, 1881 г. | |
| Диметродон лумизи | Ромер, 1937 год. | Техас Оклахома | Действительный |  | |
| Dimetrodon macrospondylus | Дело, 1907 год | Техас | Действительный | Clepsydrops macrospondylus Коп, 1884 г. Dimetrodon platycentrus Дело, 1907 год | |
| Диметродон миллери | Ромер, 1937 год. | Техас | Действительный |  | |
| Диметродон наталис | Ромер, 1936 год. | Техас | Действительный | Клепсидропс наталис Коп, 1878 г. |  |
| Диметродон западный | Берман, 1977 г. | Аризона Нью-Мексико Юта | Действительный | ||
| Диметродон тевтонис | Берман и другие., 2001 | Германия | Действительный |
Диметродон лимбатус

Диметродон лимбатус был впервые описан Эдвардом Дринкером Коупом в 1877 году как Clepsydrops limbatus.[34] (Название Клепсидропс был впервые придуман Копом в 1875 году для останков сфенакодонтидов из Округ Вермилион, Иллинойс, и позже был использован для многих образцов сфенаконтид из Техаса; многие новые виды сфенакодонтид из Техаса были отнесены к Клепсидропс или же Диметродон в конце девятнадцатого и начале двадцатого веков.) На основе образца из Красные кровати Техаса, это был первый известный синапсид с парусной спинкой. В 1940 г. палеонтологи Альфред Ромер и Ллевеллин Айвор Прайс переназначен C. limbatus к роду Диметродон, изготовление D. limbatus в типовой вид из Диметродон.[35]
Диметродон инцизивус
Первое использование имени Диметродон появился в 1878 году, когда Коуп назвал вид Диметродон инцизивус вместе с Диметродон прямой и Диметродон гигас.[33]
Диметродон прямой
Диметродон прямой был назван рядом с Диметродон инцизивус в статье Коупа 1878 года, и был единственным из трех названных видов, сохранившим удлиненные нервные отростки.[33] В 1907 г. палеонтолог E. C. Дело взолнованный D. rectiformis в вид D. incisivus.[32] D. incisivus позже был синонимом типового вида Диметродон лимбатус, изготовление D. rectiformis синоним D. limbatus.[23]
Диметродон полурадикатус
Описан в 1881 г. на основании костей верхней челюсти, Диметродон полурадикатус был последним видом, названным Копом.[42] В 1907 году Э. К. Кейс назвал D. semiradicatus с D. incisivus на основе сходства формы зубов и костей черепа.[32] D. incisivus ' и D. semiradicatus теперь считаются синонимами D. limbatus.[23]
Диметродон долловиан
Диметродон долловиан был впервые описан Эдвардом Дринкером Коупом в 1888 году как Эмболофор долловиан. В 1903 году Э. К. Кейс опубликовал подробное описание E. dollovianus, о котором он позже упоминал Диметродон.[43]
Диметродон большой

Палеонтолог Э. К. Кейс назвал новый вид синапсидов с парусной спинкой, Theropleura grandis, в 1907 г.[32] В 1940 году Альфред Ромер и Ллевеллин Айвор Прайс переназначили Theropleura grandis к Диметродон, возводя виды D. grandis.[35]
Диметродон гигас
В своей статье 1878 года об окаменелостях из Техаса Коуп назвал Clepsydrops gigas вместе с первыми названными видами Диметродон, D. limbatus, D. incisivus, и D. rectiformis.[33] Дело реклассифицировано C. gigas как новый вид Диметродон в 1907 г.[32] Кейс также описал очень хорошо сохранившийся череп Диметродон в 1904 г., причисляя его к виду Диметродон гигас.[44] В 1919 г. Чарльз В. Гилмор приписал почти полный образец Диметродон к D. gigas.[45] Диметродон гигас теперь признан синонимом D. grandis.[46]
Dimetrodon giganhomogenes

Dimetrodon giganhomogenes был назван Э. К. Кейсом в 1907 году и до сих пор считается допустимым видом Диметродон.[32][23]
Dimetrodon macrospondylus
Dimetrodon macrospondylus был впервые описан Коупом в 1884 году как Clepsydrops macrospondylus. В 1907 году Кейс реклассифицировал его как Dimetrodon macrospondylus.[32]
Dimetrodon platycentrus
Dimetrodon platycentrus был впервые описан Кейсом в его монографии 1907 года. Сейчас это считается синонимом Dimetrodon macrospondylus.[23]
Диметродон наталис

Палеонтолог Альфред Ромер установил вид Диметродон наталис в 1936 г., ранее описанный как Клепсидропс наталис. D. natalis был самым маленьким из известных видов Диметродон в то время, и был найден рядом с останками более крупных D. limbatus.[47]
Dimetrodon booneorum
Dimetrodon booneorum был впервые описан Альфредом Ромером в 1937 году на основе останков из Техаса.[47]
Кемпаэ "Диметродон"
Dimetrodon kempae был назван Ромером в 1937 году в той же статье, что и D. booneorum, D. loomisi, и Д. Миллери.[47] Dimetrodon kempae был назван на основе одной плечевой кости и нескольких позвонков, и поэтому может быть номен дубиум которые нельзя выделить как уникальный вид Диметродон.[6] В 1940 году Ромер и Прайс высказали предположение, что D. kempae не может относиться к роду Диметродон, предпочитая классифицировать его как Sphenacodontidae incertae sedis.[35]
Диметродон лумизи

Диметродон лумизи был впервые описан Альфредом Ромером в 1937 году вместе с D. booneorum, D. kempae, и Д. Миллери.[47] Останки были найдены в Техасе и Оклахоме.
Диметродон миллери

Диметродон миллери был описан Ромером в 1937 году.[47] Это один из самых мелких видов Диметродон в Северной Америке и может быть тесно связан с D. occidentalis, еще один мелкотелый вид.[40] Д. Миллери известен по двум скелетам, один почти полный (MCZ 1365), а другой менее полный, но более крупный (MCZ 1367). Д. Миллери самый старый из известных видов Диметродон.
Помимо небольшого размера, Д. Миллери отличается от других видов Диметродон в том, что его нервные отростки имеют форму круга, а не восьмерки в поперечном сечении. Его позвонки также короче по высоте относительно остальной части скелета, чем у других Диметродон разновидность. Череп высокий, а морда короткая относительно височной области. У этого вида также видны короткие позвонки и высокий череп. D. booneorum, D. limbatus и D. grandis, предлагая, что Д. Миллери может быть первым в эволюционной прогрессии между этими видами.
Dimetrodon angelensis
Dimetrodon angelensis был назван палеонтологом Эверетт К. Олсон в 1962 г.[48] Об образцах этого вида сообщалось из Формация Сан-Анджело Техаса.[49] Это также самый крупный вид Dimetrodon.
Диметродон западный
Диметродон западный был назван в 1977 году из Нью-Мексико.[39] Его название означает "западный" Диметродон"потому что это единственный североамериканский вид Диметродон известен к западу от Техаса и Оклахомы. Он был назван на основании единственного скелета, принадлежащего относительно небольшой особи. Небольшой размер D. occidentalis похож на Д. Миллери, предполагая близкие отношения. Диметродон образцы, найденные в Юте и Аризоне, вероятно, также принадлежат D. occidentalis.[40]
Диметродон тевтонис
Диметродон тевтонис был назван в 2001 году из Бассейн Саар Нахе Германии и был первым видом Диметродон будет описано за пределами Северной Америки. Также это самый мелкий вид Диметродон.[6]
Виды, отнесенные к разным родам
Диметродон крестоцвет
В 1878 году Коуп опубликовал статью под названием «Theromorphous Reptilia», в которой описал Диметродон крестоцвет.[50] D. cruciger отличался небольшими выступами, которые тянулись с обеих сторон каждого нервного отростка, как ветви дерева.[51] В 1886 году Коуп переехал D. cruciger к роду Наозавр потому что он считал его шипы такими отличными от шипов других Диметродон вид, что вид заслужил свой собственный род.[52] Наозавр позже будет синонимом Эдафозавр, род, который Коп назвал в 1882 году на основании черепов, которые, очевидно, принадлежали травоядным животным, учитывая их тупые дробящие зубы.[53]
Диметродон лонгирамус
Э. К. Кейс назвал вид Диметродон лонгирамус в 1907 г. на основе лопатки и удлиненной нижней челюсти из Формация Белль-Плейнс Техаса.[32] В 1940 году Ромер и Прайс признали, что D. longiramus материал принадлежал к тому же таксону, что и другой экземпляр, описанный палеонтологом. Сэмюэл Венделл Уиллистон в 1916 г., который включал такую же удлиненную нижнюю челюсть и длинную верхнюю челюсть.[35] Уиллистон не считал свой экземпляр принадлежащим Диметродон но вместо этого классифицировал его как офиакодонтид.[54] Ромер и Прайс отнесли образцы Кейса и Уиллистона к недавно выведенному роду и виду, Секодонтозавр longiramus, который был тесно связан с Диметродон.[35][55]
Филогенетическая классификация
Диметродон является одним из первых участников группы под названием синапсиды, к которым относятся млекопитающие и многие их вымершие родственники, хотя они не являются предком какого-либо млекопитающего (появившегося миллионы лет спустя[56]). В популярной культуре его часто принимают за динозавра, несмотря на то, что он вымер примерно за 40 миллионов лет (млн лет назад) до первого появления динозавров в Триасовый период. Как синапсид, Диметродон более тесно связан с млекопитающими, чем с динозаврами или любыми живыми рептилиями. К началу 1900-х годов большинство палеонтологов назвали Диметродон рептилия в соответствии с Линнеевская таксономия, который оценил Reptilia как учебный класс и Диметродон как род внутри этого класса. Млекопитающие были выделены в отдельный класс, и Диметродон был описан как «рептилия, похожая на млекопитающее». Палеонтологи предположили, что млекопитающие произошли от этой группы в ходе (так они называли) перехода от рептилий к млекопитающим.
Филогенетическая таксономия Synapsida

Под филогенетическая систематика, потомки последний общий предок из Диметродон и все живые рептилии будут включать всех млекопитающих, потому что Диметродон более тесно связан с млекопитающими, чем с любой живой рептилией. Таким образом, если желательно избежать клады, содержащей как млекопитающих, так и живых рептилий, тогда Диметродон не должны входить в эту кладу - или какие-либо другие «звероподобные рептилии». Потомки последнего общего предка млекопитающих и рептилий (который появился около 310 млн лет назад в Поздний карбон ) поэтому делятся на две клады: Synapsida, которая включает Диметродон и млекопитающие, и Sauropsida, который включает живых рептилий и всех вымерших рептилий, более близких к ним, чем к млекопитающим.[4]
Внутри клады Synapsida, Диметродон является частью клады Sphenacodontia, которая была впервые предложена как ранняя группа синапсидов в 1940 году палеонтологами Альфредом Ромером и Ллевеллином Айвором Прайсом вместе с группами Офиакодонтия и Эдафозаврия.[35] Все три группы известны из позднего карбона и ранней перми. Ромер и Прайс выделяли их прежде всего посткраниальный такие особенности, как форма конечностей и позвонков. Ophiacodontia считалась самой примитивной группой, потому что ее члены казались самыми рептильными, а Sphenacodontia была самой продвинутой, потому что ее члены больше всего походили на группу под названием Терапсида, в которую вошли ближайшие родственники млекопитающих. Ромер и Прайс поместили еще одну группу ранних синапсидов, названную варанопиды внутри Sphenacodontia, считая их более примитивными, чем другие сфенакодонты, такие как Диметродон.[57] Они думали, что варанопиды и Диметродон-подобные сфенакодонты были тесно связаны между собой, поскольку обе группы были плотоядными, хотя варанопиды намного меньше по размеру и более похожи на ящериц, у них нет парусов.
Современный взгляд на взаимоотношения синапсидов был предложен палеонтологом. Роберт Р. Рейс в 1986 году, чье исследование включало особенности, в основном обнаруженные в черепе, а не в посткраниальном скелете.[58] Диметродон до сих пор считается сфенакодонтом в соответствии с этим филогения, но варанодонтиды теперь считаются более базальный синапсиды, выходящие за пределы клады Sphenacodontia. Внутри Sphenacodontia находится группа Sphenacodontoidea, который, в свою очередь, содержит Sphenacodontidae и Терапсида. Sphenacodontidae - это группа, содержащая Диметродон и несколько других синапсидов с парусом, например Sphenacodon и Секодонтозавр, пока Терапсида включает млекопитающих и их преимущественно пермских и Триасовый родственники.
Ниже кладограмма Clade Synapsida, которая следует этой филогении Synapsida как изменено на основе анализа Benson (2012).[57]

| Амниота |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
На следующей кладограмме показаны отношения нескольких Диметродон виды, из Brink и другие., (2015).[59]
| Sphenacodontidae |
| ||||||||||||||||||||||||||||||||||||
Палеобиология
Функция нервных отростков

Палеонтологи предложили множество способов, которыми парус мог функционировать при жизни. Некоторые из первых, кто задумывался о его назначении, предположили, что парус мог служить камуфляжем среди тростников во время Диметродон ждали добычи, или как настоящий парус, похожий на лодку, чтобы поймать ветер, пока животное находилось в воде.[60] Другой заключается в том, что длинные нервные отростки могли стабилизировать туловище, ограничивая движение вверх и вниз, что позволило бы более эффективно перемещаться из стороны в сторону при ходьбе.[21]
Терморегуляция
В 1940 г. Альфред Ромер и Ллевеллин Айвор Прайс предположили, что парус выполняет терморегулирующую функцию, позволяя людям согревать свое тело солнечным теплом. В последующие годы было создано множество моделей для оценки эффективности терморегуляции в Диметродон. Например, в статье 1973 года в журнале Природа палеонтологи К. Д. Брамвелл и П. Б. Феллгетт подсчитали, что человеку весом 200 кг (440 фунтов) требуется около полутора часов, чтобы температура его тела поднялась с 26 до 32 ° C (79–90 ° F).[61] В 1986 году Стивен К. Хаак пришел к выводу, что потепление происходило медленнее, чем считалось ранее, и что процесс, вероятно, длился четыре часа. Использование модели, основанной на различных факторах окружающей среды и предполагаемых физиологических аспектах Диметродон, Хаак обнаружил, что парус позволяет Диметродон чтобы согреваться быстрее утром и достичь немного более высокой температуры тела в течение дня, но это было неэффективно для выделения избыточного тепла и не позволяло Диметродон для сохранения повышенной температуры тела ночью.[62] В 1999 году группа инженеров-механиков создала компьютерную модель для анализа способности паруса регулировать температуру тела в разное время года и пришла к выводу, что парус полезен для улавливания и выделения тепла в любое время года.[63]

Большинство этих исследований отводят пару терморегулирующих свойств парусу. Диметродон: один как средство быстрого согрева по утрам, а другой как средство для охлаждения, когда температура тела становится высокой. Диметродон и все остальные раннепермские наземные позвоночные были хладнокровными или хладнокровными. пойкилотермный, полагаясь на солнце для поддержания высокой температуры тела. Из-за большого размера Диметродон был высокий тепловая инерция, что означает, что изменение температуры тела у него происходило медленнее, чем у животных с меньшим размером тела. По мере того, как по утрам температура повышалась, мелкая добыча Диметродон могут согреть их тела намного быстрее, чем что-то размером с Диметродон. Многие палеонтологи, включая Хаака, предположили, что парус Диметродон мог позволить ему быстро согреться утром, чтобы успевать за своей добычей.[62] Большая площадь поверхности паруса также означала, что тепло могло быстро рассеиваться в окружающей среде, что было полезно, если животному нужно было выделять избыточное тепло, производимое метаболизмом или поглощаемое солнцем. Диметродон мог отклонить свой парус от солнца, чтобы охладиться, или ограничил приток крови к парусу, чтобы поддерживать тепло ночью.[60]
В 1986 году Дж. Скотт Тернер и К. Ричард Трейси предположили, что эволюция паруса в Диметродон был связан с эволюцией теплокровности у предков млекопитающих. Они думали, что парус Диметродон позволил этому быть теплый, поддерживая постоянную, хоть и низкую, температуру тела. Млекопитающие также теплокровны, хотя и отличаются от Диметродон в бытии эндотермический, контролируя температуру своего тела изнутри за счет повышенного метаболизма. Тернер и Трейси отметили, что ранние терапсиды, более развитая группа синапсидов, тесно связанных с млекопитающими, имели длинные конечности, которые могли выделять тепло так же, как и парус парусника. Диметродон. Гомеотермия, развившаяся у таких животных, как Диметродон могли быть перенесены на терапсиды через изменение формы тела, что в конечном итоге переросло в теплокровность млекопитающих.[64]

Недавние исследования паруса Диметродон и другие сфенакодонтиды поддерживают утверждение Хаака 1986 года о том, что парус был плохо приспособлен к выделению тепла и поддержанию стабильной температуры тела. Наличие парусов у мелкотелых видов Диметродон Такие как Д. Миллери и D. teutonis не соответствует идее о том, что целью паруса была терморегуляция, потому что паруса меньшего размера менее способны передавать тепло и потому что маленькие тела могут легко поглощать и выделять тепло самостоятельно. Более того, близкие родственники Диметродон Такие как Sphenacodon имеют очень низкие гребни, которые были бы бесполезны в качестве терморегулирующих устройств.[23] Большой парус Диметродон Считается, что он постепенно развивался из этих меньших гребней, а это означает, что на протяжении большей части эволюционной истории паруса терморегуляция не могла выполнять важную функцию.[65]
Более крупные экземпляры Диметродон имеют большие паруса относительно их размера, пример положительная аллометрия. Положительная аллометрия может принести пользу терморегуляции, потому что это означает, что по мере роста человека площадь поверхности увеличивается быстрее, чем масса. Животные с более крупным телом выделяют много тепла в процессе обмена веществ, и количество тепла, которое должно рассеиваться с поверхности тела, значительно больше, чем то, что должно рассеиваться животными с меньшим телом. Эффективное рассеивание тепла можно предсказать для множества разных животных с помощью единственного отношения между массой и площадью поверхности. Однако исследование аллометрии в 2010 г. Диметродон обнаружили иную взаимосвязь между его парусом и массой тела: фактический масштабный показатель паруса был намного больше, чем показатель, ожидаемый у животного, приспособленного к рассеиванию тепла. Исследователи пришли к выводу, что парус Диметродон росли гораздо быстрее, чем это было необходимо для терморегуляции, и предположили, что половой отбор был основной причиной его эволюции.[65]
Половой отбор
Аллометрический показатель для высоты паруса по величине аналогичен масштабированию межвидовой длины рога на высоту плеча в шейки матки. Более того, как заметил Баккер (1970) в контексте Диметродонмногие виды ящериц поднимают спинной гребень кожи во время демонстрации угрозы и ухаживания, а у современных ящериц присутствуют положительно аллометрические, сексуально диморфные оборки и подвесы (Echelle et al. 1978; Christian et al. 1995). Есть также свидетельства полового диморфизма, как в прочности скелета, так и в относительной высоте шипов. D. limbatus (Ромер и Прайс, 1940).[65]
Половой диморфизм
Диметродон могло быть сексуально диморфный, что означает, что мужчины и женщины имели немного разные размеры тела. Некоторые образцы Диметродон были выдвинуты как мужчины, потому что у них более толстые кости, большие паруса, более длинные черепа и более выраженные верхнечелюстные «ступеньки», чем у других. Исходя из этих различий, смонтированные скелеты в Американский музей естественной истории (AMNH 4636) и Полевой музей естественной истории могут быть мужчины и скелеты в Денверский музей природы и науки (MCZ 1347) и Музей естественной истории Мичиганского университета могут быть самки.[35]


Палеоэкология

Окаменелости Диметродон известны из США (Техас, Оклахома, Нью-Мексико, Аризона, Юта и Огайо) и Германии, областей, которые были частью суперконтинента Euramerica в ранней перми. В Соединенных Штатах почти все материалы, приписываемые Диметродон происходит из трех геологических групп в северо-центральном Техасе и южно-центральной Оклахоме: Очистить группу вилок, то Wichita Group, а Pease River Group.[66][67] Большинство находок окаменелостей относится к экосистемам низменностей, которые в пермский период представляли собой обширные водно-болотные угодья. В частности, Красные пласты Техаса - район большого разнообразия ископаемых. четвероногие, или четырехногие позвоночные. В добавление к Диметродон, наиболее распространенные четвероногие в красных пластах и во всех раннепермских отложениях на юго-западе США, являются амфибиями. Archeria, Диплокаулус, Эриопс, и Trimerorhachis, то рептилиоморф Сеймурия, рептилия Captorhinus, а синапсиды Офиакодон и Эдафозавр. Эти четвероногие составляли группу животных, которую палеонтологи Эверетт К. Олсон названный «хронофауной пермо-карбона», фауна которые доминировали в континентальной евразийской экосистеме в течение нескольких миллионов лет.[68] Основываясь на геологии таких месторождений, как Красные пласты, считается, что фауна населяла хорошо заросшие низины. дельтовая экосистема.[69]
Пищевой сети

Олсон сделал много выводов о палеоэкологии Кровати Texas Red и роль Диметродон в своей экосистеме. Он предложил несколько основных типов экосистем, в которых жили самые ранние четвероногие. Диметродон принадлежал к наиболее примитивной экосистеме, которая возникла из водных пищевых сетей. В нем водные растения были первичные производители и в основном питались рыбы и водные беспозвоночные. Большинство наземных позвоночных питались этими основными водными потребителями. Диметродон вероятно был главный хищник экосистемы Red Beds, питаясь различными организмами, такими как акула Ксенакант[нужна цитата ], водные амфибии Trimerorhachis и Диплокаулус, а наземные четвероногие Сеймурия и Трематопы. Насекомые известны из раннепермских красных пластов и, вероятно, в какой-то степени участвовали в той же пищевой сети, что и Диметродон, кормление мелких рептилий, таких как Captorhinus. Комплекс Red Beds также включал некоторых из первых крупных наземных травоядных животных, таких как Эдафозавр и Диадекты. Питаясь в основном наземными растениями, эти травоядные не получали энергию от водных пищевых сетей. По мнению Олсона, лучший современный аналог экосистемы Диметродон обитаемый Эверглейдс.[69] Точный образ жизни Диметродон (от земноводных до наземных) долгое время оставалось спорным, но микроанатомия костей поддерживает наземный образ жизни,[70] что означает, что он питался бы в основном на суше, на берегах или на очень мелководье. Также существуют доказательства Диметродон охотясь на праздники Диплокаулус во время засухи с тремя частично съеденными молодыми Диплокаулус в норе из восьми несущих зубов следы от Диметродон которые откопали и убили их.[71]
Единственный вид Диметродон найден за пределами юго-запада США, D. teutonis из Германии. Его останки были найдены в Тамбачская свита в ископаемом месте под названием Бромакер. Ансамбль Бромакера ранней перми четвероногие необычен тем, что существует несколько крупных синапсидов, выполняющих роль высших хищников. D. teutonis по оценкам, был всего 1,7 метра (5,6 фута) в длину, слишком мал, чтобы охотиться на крупных диадектида травоядные животные, которые многочисленны в сообществе бромакеров. Он, скорее, ел мелких позвоночных и насекомых. Только три окаменелости могут быть отнесены к крупным хищникам, и считается, что они были либо крупными, либо крупными. варанопиды или маленький сфенакодонты, оба из которых потенциально могут охотиться на D. teutonis. В отличие от низменности дельтовый Красные пласты Техаса, месторождения Бромакер, как полагают, представляли собой возвышенную среду без водных видов. Возможно, что крупные плотоядные животные не были частью сообщества Бромакеров, потому что они зависели от крупных водных организмов. амфибии для пищевых продуктов.[6]
Рекомендации
- ^ «Диметродон». Оксфордские словари Британский словарь. Oxford University Press. Получено 2016-01-21.
- ^ Диметродон. Dictionary.com. Dictionary.com Без сокращений. Random House, Inc. http://www.dictionary.com/browse/dimetrodon (дата обращения: 12 февраля 2018 г.).
- ^ «Диметродон». База данных палеобиологии. Получено 23 августа 2012.
- ^ а б c d Ангельчик, К. Д. (2009). «Диметродон - не динозавр: использование древовидного мышления для понимания древних родственников млекопитающих и их эволюции». Эволюция: образование и пропаганда. 2 (2): 257–271. Дои:10.1007 / s12052-009-0117-4.
- ^ Huttenlocker, A.K .; Рега, Э. (2012). "Палеобиология и микроструктура костей пеликозавровских синапсид". В Чинсами, А. (ред.). Предшественники млекопитающих: радиация, гистология, биология. Издательство Индианского университета. С. 90–119. ISBN 978-0-253-35697-0.
- ^ а б c d е Berman, D.S .; Reisz, R.R .; Martens, T .; Хенрици, A.C. (2001). "Новый вид Диметродон (Synapsida: Sphenacodontidae) из нижней перми Германии регистрирует первое появление рода за пределами Северной Америки » (PDF). Канадский журнал наук о Земле. 38 (5): 803–812. Bibcode:2001CaJES..38..803B. Дои:10.1139 / cjes-38-5-803.
- ^ «Кунгурский век». Палеос. Архивировано из оригинал 12 ноября 2013 г.. Получено 29 сентября 2013.
- ^ а б c d Fröbisch, J .; Schoch, R.R .; Müller, J .; Шиндлер, Т .; Швейс, Д. (2011). «Новый базальный синапсид сфенакодонтида из позднего карбона бассейна Саар-Наэ, Германия» (PDF). Acta Palaeontologica Polonica. 56 (1): 113–120. Дои:10.4202 / app.2010.0039. S2CID 45410472.
- ^ Olson, E.C .; Бирбауэр, Дж. Р. (1953). "Формация Сан-Анджело, Пермь Техаса, и ее позвоночные". Журнал геологии. 61 (5): 389–423. Bibcode:1953JG ..... 61..389O. Дои:10.1086/626109. S2CID 128681671.
- ^ Олсон, Э. К. (1955). «Параллелизм в эволюции фаун пермских рептилий Старого и Нового Света». Fieldiana. 37 (13): 385–401.
- ^ Штернберг, C.W. (1942). "Скелет незрелого пеликозавра, Диметродон ср. grandis, из перми Техаса ». Журнал палеонтологии. 16 (4): 485–486. JSTOR 1298848.
- ^ Лаурин, М .; Рейс, Р.Р. (2012). «Синапсиды: млекопитающие и их вымершие родственники». Веб-проект "Древо жизни". Получено 24 августа 2012.
- ^ «Экспонаты: Диметродон». Американский музей естественной истории. Архивировано из оригинал 4 июля 2012 г.. Получено 2 июля 2012.
- ^ а б Baur, G .; Кейс, E.C. (1899). «История пеликозавров с описанием рода Диметродон, Справиться". Труды Американского философского общества. 20 (1): 5–62. Дои:10.2307/1005488. HDL:2027 / uc1.32106020416696. JSTOR 1005488.
- ^ а б Abler, W.L. 2001. Модель зубцов тираннозавра в виде пропилов и сверлений. п. 84-89. В: Мезозойская жизнь позвоночных. Под ред. Танке, Д. Х., Карпентер, К., Скрепник, М. В. Издательство Индианского университета.
- ^ «Великий энергетический вызов». Среда.
- ^ Кемп, Т. (2006). «Происхождение и раннее облучение терапсидных рептилий, подобных млекопитающим: палеобиологическая гипотеза». Журнал эволюционной биологии. 19 (4): 1231–1247. Дои:10.1111 / j.1420-9101.2005.01076.x. PMID 16780524. S2CID 3184629.
- ^ Лаурин, М .; Reisz, R.R. (1997). «Аутапоморфии основных клад синапсидов». Веб-проект "Древо жизни". Получено 24 августа 2012.
- ^ Ромер, А. (1927). "Заметки о пермо-каменноугольных рептилиях" Диметродон". Журнал геологии. 35 (8): 673–689. Bibcode:1927JG ..... 35..673R. Дои:10.1086/623462. JSTOR 30060393. S2CID 140679339.
- ^ а б Rega, E. A .; Noriega, K .; Sumida, S. S .; Huttenlocker, A .; Ли, А .; Кеннеди, Б. (2012). «Исцеленные переломы в нервных отростках связанного скелета диметродона: последствия для морфологии и функции спинного паруса». Филдиана Науки о жизни и Земле. 5: 104–111. Дои:10.3158/2158-5520-5.1.104. S2CID 108887164.
- ^ а б c Rega, E .; Сумида, S .; Noriega, K .; Pell, C .; Ли, А. (2005). "Доказательная палеопатология I: онтогенетические и функциональные последствия спинных парусов в Диметродон". Журнал палеонтологии позвоночных. 25 (S3): 103A. Дои:10.1080/02724634.2005.10009942. S2CID 220413556.
- ^ Сумида, S .; Rega, E .; Норьега, К. (2005). "Доказательная палеопатология II: влияние на филогенетический анализ рода" Диметродон". Журнал палеонтологии позвоночных. 25 (S3): 120А. Дои:10.1080/02724634.2005.10009942. S2CID 220413556.
- ^ а б c d е ж грамм Huttenlocker, A.K .; Rega, E .; Сумида, С.С. (2010). «Сравнительная анатомия и остеогистология гипер удлиненных нервных отростков сфенакодонтид». Sphenacodon и Диметродон (Амниота: Synapsida) ". Журнал морфологии. 271 (12): 1407–1421. Дои:10.1002 / jmor.10876. PMID 20886514. S2CID 40899700.
- ^ Rega, E.A .; Noriega, K .; Сумида, S .; Ли, А. (2004). «Гистологический анализ травматического повреждения множественных нервных отростков связанного скелета Диметродон: Влияние на реакцию заживления, морфологию спинного паруса и возраст смерти в синапсидах нижней перми ». Интегративная и сравнительная биология. 44: 628.
- ^ Константинович Чудинов, Петр (1965). «Новые факты о фауне верхней перми СССР». Журнал геологии. 73: 117–30. Дои:10.1086/627048. S2CID 129428120.
- ^ Botha-Brink, J .; Модесто, С. П. (2007). «Скопление« пеликозавров »разновозрастного класса из Южной Африки: самые ранние свидетельства родительской заботы о амниотах?». Труды Королевского общества B. 274 (1627): 2829–2834. Дои:10.1098 / rspb.2007.0803. ЧВК 2288685. PMID 17848370.
- ^ Niedźwiedzki, G .; Бояновский, М. (2012). «Предполагаемый отпечаток тела евпеликозавра из ранней перми внутри Судетского бассейна, Польша». Ичнос. 19 (3): 150–155. Дои:10.1080/10420940.2012.702549. S2CID 129567176.
- ^ Шпиндлер, Фредерик; Вернебург, Ральф; Schneider, Joerg W .; Luthardt, Людвиг; Аннакер, Фолькер; Рёсслер, Ронни (2018). «Первые древесные« пеликозавры »(Synapsida: Varanopidae) из ранней перми Chemnitz Fossil Lagerstätte, SE Германия, с обзором филогении варанопидов». PalZ. 92 (2): 315–364. Дои:10.1007 / s12542-018-0405-9. S2CID 133846070.
- ^ Модесто, Шон П. (январь 2020 г.). «Болеют за рептилии». Природа Экология и эволюция. 4 (1): 10–11. Дои:10.1038 / s41559-019-1074-0. ISSN 2397-334X. PMID 31900449. S2CID 209672518.
- ^ Форд, Дэвид П .; Бенсон, Роджер Б. Дж. (2019). «Повторное описание Orovenator mayorum (Sauropsida, Diapsida) с использованием микроконтактной томографии высокого разрешения и последствия для ранней филогении амниот». Статьи по палеонтологии. 5 (2): 197–239. Дои:10.1002 / spp2.1236. ISSN 2056-2802.
- ^ Мэтью, W.D. (1908). "Обзор ревизии Кейса пеликозавров Северной Америки"'". Наука. 27 (699): 816–818. Bibcode:1908Научный .... 27..816М. Дои:10.1126 / science.27.699.816.
- ^ а б c d е ж грамм час я Кейс, E.C. (1907). Ревизия пеликозавров Северной Америки. Вашингтон, округ Колумбия: Вашингтонский институт Карнеги. стр.1 –176.
- ^ а б c d е Коуп, Э. (1878). «Описание вымерших Batrachia и Reptilia из пермских формаций Техаса». Труды Американского философского общества. 17 (101): 505–530. JSTOR 982652.
- ^ а б Коуп, Э. (1877). «Описание вымерших позвоночных из пермских и триасовых формаций США». Труды Американского философского общества. 17 (100): 182–193. JSTOR 982295.
- ^ а б c d е ж грамм час Romer, A.S .; Прайс, Л. (1940).«Обзор пеликозавров». Специальный доклад Геологического общества Америки. Специальные статьи Геологического общества Америки. 28: 1–538. Дои:10.1130 / spe28-p1.
- ^ Вон, П. (1966). «Сравнение раннепермской фаун позвоночных в районе Четырех углов и северо-центральной части Техаса» (PDF). Вклад в науку. 105. Музей естественной истории округа Лос-Анджелес: 1–13. Архивировано из оригинал (PDF) на 2017-01-10. Получено 2016-04-21. Цитировать журнал требует
| журнал =(помощь) - ^ Вон, П. (1969). «Раннепермские позвоночные с юга Нью-Мексико и их палеозоогеографическое значение» (PDF). Вклад в науку. 166. Музей естественной истории округа Лос-Анджелес: 1–22. Архивировано из оригинал (PDF) на 2017-01-10. Получено 2016-04-21. Цитировать журнал требует
| журнал =(помощь) - ^ Олсон, Э. К. (1975). «Пермо-карбоновая палеоэкология и морфотипическая серия». Американский зоолог. 15 (2): 371–389. Дои:10.1093 / icb / 15.2.371.
- ^ а б c Берман, Д.С. (1977). "Новый вид Диметродон (Reptilia, Pelycosauria) из недельтовой фации в нижней перми северо-центральной части штата Нью-Мексико ». Журнал палеонтологии. 51 (1): 108–115. JSTOR 1303466.
- ^ а б c Madalena, K .; Сумида, S .; Zeigler, K .; Рега, Э. (2007). "Новый рекорд раннепермского синапсида класса пеликозавров. Диметродон (Eupelycosauria: Sphenacodontidae) из нижней группы Катлера (ранняя пермь) Джемес-Пуэбло, северо-центральная часть Нью-Мексико ". Журнал палеонтологии позвоночных. 27 (3, Прил.): 110А. Дои:10.1080/02724634.2007.10010458.
- ^ Вон, П. (1973). Позвоночные животные группы Катлер Долины монументов и окрестностей (PDF). Путеводитель Геологического общества Нью-Мексико. 24. Геологическое общество Нью-Мексико. С. 99–105.
- ^ Коуп, Э. (1881). «О некоторых новых Batrachia и Reptilia из пермских красных пластов Техаса». Бюллетень Геологической службы США. 6: 79–82.
- ^ Кейс, E.C. (1903). "Остеология Эмболофор долловиан, Справились, с попыткой восстановления ". Журнал геологии. 11 (1): 1–28. Bibcode:1903JG ..... 11 .... 1С. Дои:10.1086/621055. JSTOR 30056735.
- ^ Кейс, E.C. (1904). «Остеология черепа из рода пеликозавров, Диметродон". Журнал геологии. 12 (4): 304–311. Bibcode:1904JG ..... 12..304C. Дои:10.1086/621157. JSTOR 30055825.
- ^ Гилмор, C.W. (1919). "Навесной скелет Диметродон гигас в Национальном музее США, с заметками об анатомии скелета " (PDF). Труды Национального музея США. 56 (2300): 525–539. Дои:10.5479 / si.00963801.56-2300.525.
- ^ Henrici, A.C .; Berman, D.S .; Lucas, S.G .; Heckert, A.B .; Rinehart, L.F .; Цейглер, К. (2005). "Запястья и предплюсны раннепермских синапсидов. Sphenacodon ferox (Eupelycosauria: Sphenacodontidae) " (PDF). В Lucas, S.G .; Цейглер, К. (ред.). Неморская пермь. Бюллетень Музея естественной истории и науки Нью-Мексико. 30. Альбукерке: Музей естественной истории и науки Нью-Мексико. С. 106–110.
- ^ а б c d е Ромер, А. (1937). «Новые роды и виды рептилий пеликозавров» (PDF). Труды Зоологического клуба Новой Англии. 16: 89–97.
- ^ Олсон, Э. К. (1962). «Позднепермские наземные позвоночные, США и СССР». Труды Американского философского общества. 52 (2): 1–224. Дои:10.2307/1005904. JSTOR 1005904.
- ^ Баттейл, Б. (2000). «Сравнение позднепермской гондванской и лауразийской фаун амниот». Журнал африканских наук о Земле. 31: 165–174. Дои:10.1016 / S0899-5362 (00) 00081-6.
- ^ Коуп, Э. (1878). "Тероморфные рептилии". Американский натуралист. 12 (12): 829–830. Дои:10.1086/272251.
- ^ Коуп, Э. (1880 г.). «Второй вклад в историю позвоночных пермской формации Техаса». Труды Американского философского общества. 19 (107): 38–58. JSTOR 982605.
- ^ Коуп, Э. (1886). "Длинноиглая тероморфа пермской эпохи". Американский натуралист. 20 (6): 544–545. Дои:10.1086/274275.
- ^ Коуп, Э. (1882 г.). «Третий вклад в историю позвоночных пермской формации Техаса». Труды Американского философского общества. 20 (112): 447–461. JSTOR 982692.
- ^ Уиллистон, С. (1916). «Остеология некоторых американских пермских позвоночных, II». Вклад музея Уокера. 1: 165–192.
- ^ Reisz, R. R .; Берман, Д. С .; Скотт, Д. (1992). «Черепная анатомия и взаимоотношения Secodontosaurus, необычного млекопитающего рептилий (Synapsida: Sphenacodontidae) из ранней перми Техаса». Зоологический журнал Линнеевского общества. 104 (2): 127–184. Дои:10.1111 / j.1096-3642.1992.tb00920.x. S2CID 56425294.
- ^ Появляются млекопитающие
- ^ а б Бенсон, Р.Дж. (2012). «Взаимоотношения базальных синапсидов: черепные и посткраниальные морфологические перегородки предполагают разную топологию». Журнал систематической палеонтологии. 10 (2): 601–624. Дои:10.1080/14772019.2011.631042. S2CID 84706899.
- ^ Рейс, Р. Р. (1986). «Пеликозаврия». В Sues, H.-D. (ред.). Handbuch der Paläoherpetologie. 17А. Густав Фишер Верлаг. С. 1–102. ISBN 978-3-89937-032-4. ПР 12985656M.
- ^ Бринк, Кирстин С .; Maddin, Hillary C .; Эванс, Дэвид С .; Reisz, Robert R .; Сьюз, Ханс-Дитер (2015). "Переоценка исторической канадской окаменелости Батигнат borealis из ранней перми острова Принца Эдуарда ". Канадский журнал наук о Земле. 52 (12): 1109–1120. Bibcode:2015CaJES..52.1109B. Дои:10.1139 / cjes-2015-0100.
- ^ а б Florides, G.A .; Kalogirou, S.A .; Tassou, S.A .; Вробель, Л. (2001). «Природная среда и термическое поведение Диметродон лимбатус". Журнал термобиологии. 26 (1): 15–20. Дои:10.1016 / S0306-4565 (00) 00019-X. PMID 11070340.
- ^ Bramwell, C.D .; Феллгетт, П. (1973). «Терморегуляция у парусных ящериц». Природа. 242 (5394): 203–205. Bibcode:1973Натура.242..203Б. Дои:10.1038 / 242203a0. S2CID 4159825.
- ^ а б Хаак, С.С. (1986). «Тепловая модель парусного пеликозавра». Палеобиология. 12 (4): 450–458. Дои:10.1017 / S009483730000316X.
- ^ Florides, G.A .; Wrobel, L.C .; Kalogirou, S.A .; Тассу, С.А. (1999). «Тепловая модель рептилий и пеликозавров». Журнал термобиологии. 24 (1): 1–13. Дои:10.1016 / S0306-4565 (98) 00032-1.
- ^ Тернер, J.S .; Трейси, C.R. (1986). «Размер тела, гомеотермия и контроль теплообмена у рептилий, подобных млекопитающим» (PDF). В Хоттоне, Н. III .; MacLean, P.D .; Roth, J.J .; Рот, E.C. (ред.). Экология и биология млекопитающих рептилий. Вашингтон, округ Колумбия: Smithsonian Institution Press. С. 185–194.
- ^ а б c Tomkins, J.L .; LeBas, N.R .; Witton, M.P .; Martill, D.M .; Хамфрис, С. (2010). «Позитивная аллометрия и предыстория полового отбора» (PDF). Американский натуралист. 176 (2): 141–148. Дои:10.1086/653001. PMID 20565262. S2CID 36207. Архивировано из оригинал (PDF) на 2016-04-12.
- ^ Lucas, S.G .; Spielmann, J.A .; Rinehart, L.F .; Мартенс, Т. (2009). Диметродон (Amniota: Synapsida: Sphenacodontidae) из нижнепермской формации Або, округ Сокорро, Нью-Мексико (PDF). Путеводитель Геологического общества Нью-Мексико. 60. Геологическое общество Нью-Мексико. С. 281–284.
- ^ Нельсон, Джон В., Роберт В. Хук и Дэн С. Чейни (2013). Литостратиграфия нижнепермской (леонардовской) формации прозрачной вилки Северо-Центрального Техаса из Каменноугольно-пермский переход: Бюллетень 60, изд. Спенсер Г. Лукас и др. Музей естественной истории и науки Нью-Мексико, стр. 286-311. Проверено 28 декабря 2017 года.
- ^ Салливан, К .; Reisz, R.R .; Мэй, W.J. (2000). «Крупные диссорофоидные элементы скелета из трещин Ричардс Спур в нижней перми, Оклахома, и их палеоэкологические последствия». Журнал палеонтологии позвоночных. 20 (3): 456–461. Дои:10.1671 / 0272-4634 (2000) 020 [0456: LDSEFT] 2.0.CO; 2. JSTOR 4524117.
- ^ а б Олсон, E.C. (1966). «Эволюция сообщества и происхождение млекопитающих». Экология. 47 (2): 291–302. Дои:10.2307/1933776. JSTOR 1933776.
- ^ Kriloff, A .; Germain, D .; Canoville, A .; Vincent, P .; Sache, M .; Лаурин, М. (2008). «Эволюция костной микроанатомии большеберцовой кости четвероногих и ее использование в палеобиологических выводах». Журнал эволюционной биологии. 21 (3): 807–826. Дои:10.1111 / j.1420-9101.2008.01512.x. PMID 18312321. S2CID 6102313.
- ^ "Плавниковое чудовище отрубило головы древним амфибиям".
внешняя ссылка
- Диметродон Палеос страница на Диметродон
- Знакомство с пеликозаврами Музей палеонтологии Калифорнийского университета веб-страница на ранних синапсидах, включая Диметродон
- «Диметродон: наш самый маловероятный предок». PBS Eons. 21 августа 2017 г. - через YouTube.
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||



