Железосвязывающие белки - Iron-binding proteins
Железосвязывающие белки находятся белки-носители и металлопротеины что важно в железе метаболизм[1] и иммунный ответ.[2][3] Железо необходимо для жизни.
Железозависимые ферменты катализируют множество различных биохимических реакций и могут быть разделены на три широких класса в зависимости от структуры их активного центра: моноядерные кластеры железа, центры Риске (Fe-S) или гемовые центры.[4] Хорошо известное семейство железозависимых ферментов включает оксигеназы, которые способствуют присоединению гидроксильной группы одного или обоих атомов двухатомного кислорода. Известные ферменты включают триптофандиоксигеназу, ферредоксин и 2-оксоглутаратдиоксигеназу (основной фермент биосинтеза флавоноидов).[5]
Биодоступность железа
Железо необходимо для жизни. Дефицит железа приводит к анемия. В окружающей среде железо очень распространено, но мало его биодоступно.[6] Чтобы быть полезным с биологической точки зрения, неорганическое железо обычно восстанавливают с трехвалентного до двухвалентного железа редуктазами трехвалентного железа. Металлы, как правило, более растворимы вблизи нейтрального pH. «Свободное железо» цитотоксично, поскольку участвует в Фентон химия производить активные формы кислорода. Таким образом, переизбыток железа также может быть вредным, что демонстрирует болезнь. гемохроматоз.
Люди обычно получают большую часть своего железа из растительных или животных источников. Это железо может быть гемовым (в мясе животных) или негемовым (в растениях). Железо в геме гораздо более биологически доступно для человека (усваивается на 15-35%), поэтому мясо может быть мощным источником диетического железа.[6] Негемовое железо обычно усваивается только на 2-20%. Однако большая часть железа в рационе среднего человека поступает из негемовых источников просто потому, что потребляется большее количество растительной пищи.[7] Железо прочно удерживается организмом, и единственный способ избавиться от лишнего железа для человека - это потеря крови. Следовательно, потребление железа регулируется, когда абсорбируется относительно низкая доля железа. Гепсидин является гормональным регулятором захвата железа в кишечнике. На клеточных мембранах кишечника переносчик двухвалентного металла-1 активно переносит железо в клетку.[8] Затем он улавливается ферритином для дальнейшего использования. Через гепсидин поглощение железа клетками кишечника может регулироваться физиологическими потребностями, такими как дефицит или избыток железа.
Гемовые белки

Гемовые протеины - это протеины, содержащие простетическую группу гема. Группа гема состоит из порфирин кольцо, координированное с ионом железа. Четыре атома азота в порфириновом кольце действуют как лиганд для железа в центре. Во многих случаях экваториальный порфирин дополняется одним или двумя аксиальными лигандами. Примером этого является гемоглобин, где порфирин работает вместе с боковой цепью гистидина и связанным O2 молекула, образующая восьмигранный сложный.
Синтез порфирина (т.е. анаболизм гема) высоко консервативен во всех сферах жизни. Он начинается с производства δ-аминолевулиновая кислота из глицина и сукцинил-КоА следует с образованием тетрапиррольного кольца и заканчивается присоединением атома железа к хелатирующему кольцу. Синтез порфирина негативно регулируется (т.е. из-за отсутствия / присутствия репрессора) повышенными концентрациями глюкозы и гема. Присутствие этих ингибиторов дестабилизирует комплекс мРНК-рибосома, а также предотвращает его захват митохондриями для биосинтеза митохондриального белка.
Гемоглобин
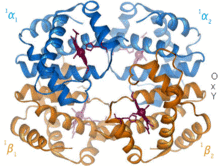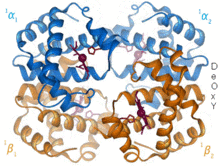
Гемоглобин - это белок, переносящий кислород, который встречается практически у всех позвоночных. Гемоглобин А - основной тип, обнаруживаемый у взрослых людей. Это тетрамер, состоящий из двух альфа и двух бета-субъединиц. Каждая из четырех мономерных единиц содержит простетическую группу гема, в которой катион трехвалентного железа связан между четырьмя атомами азота порфиринового кольца. Наряду с гистидином апоформа имеет пять лигандов, окружающих атом железа. Кислород связывается с пустой шестой позицией, образуя октаэдрический комплекс в голо форме.[9] Связывание кислорода полностью кооперативно для каждой из субъединиц, потому что, когда первый кислород связывается с одной из четырех гемовых групп, белок претерпевает резкие конформационные изменения, которые резко увеличивают сродство к кислороду трех других субъединиц.[10]
Гемоглобин имеет различное сродство в зависимости от pH, структуры и CO.2 частичное давление. Гемоглобин плода - это вариант, содержащий две гамма-субъединицы вместо двух бета-субъединиц. Гемоглобин плода является преобладающей формой до тех пор, пока ребенку не исполнится несколько месяцев, и он имеет большее сродство к кислороду, чтобы компенсировать низкое давление кислорода в поставляемой материнской крови во время беременности.[11] Гемоглобин имеет более низкое сродство к кислороду при низком pH. Это обеспечивает быструю диссоциацию, поскольку насыщенный кислородом гемоглобин транспортируется к клеткам по всему телу. Из-за CO2 При производстве и водном образовании угольной кислоты в дышащих клетках оксигенированный гемоглобин диссоциирует, чтобы доставить в клетки необходимый кислород.[12] Гемоглобин имеет сродство к связыванию монооксид углерода это в 250 раз больше, чем для кислорода. Это основа отравление угарным газом, поскольку гемоглобин больше не может транспортировать кислород к клеткам.
Цитохромы
Цитохромы - это гемсодержащие ферменты, которые действуют как переносчики одиночных электронов, прежде всего как переносчики электронов при окислительном фосфорилировании и фотосинтезе. Типы хорошо изученных цитохромов включают цитохромы a-c, цитохромоксидазу и цитохром P450.[13] Эти белки действуют как электронные челноки, переключая степень окисления атома железа гема между двухвалентным (Fe2+) и трехвалентное железо (Fe3+). Различные цитохромы в сочетании с другими окислительно-восстановительными молекулами образуют градиент стандартных восстановительных потенциалов, который увеличивает эффективность взаимодействия энергии во время событий переноса электронов.
Белки железо-сера
Белки железо-сера - это белки со структурой железа, которая включает серу. Железо и сера могут принимать различные формы в белках, но наиболее распространенными являются [2Fe 2S] и [4Fe 4S]. Кластеры часто связаны с остатками цистеина в белковой цепи.[14]
Негемовые белки
Трансферрин
Трансферрин содержится в плазме крови человека и используется для транспортировки и импорта негемового железа.[15] Он свободно перемещается во внеклеточном пространстве.[16] Когда его железо необходимо клетке, оно попадает в цитозоль по рецептор трансферрина. Трансферрин может связывать два иона Fe (III) вместе с анионом (обычно карбонатом). Чтобы высвободить железо, карбонатный анион протонируется. Это изменяет взаимодействие карбоната с белком, изменяя конформацию и позволяя переносить Fe (III).
Трансферрин имеет молекулярную массу около 80 кДа. Это гликопротеин, что означает, что к его аминокислотной цепи присоединены сахара.
Лактоферрин
Лактоферрин является членом семейства трансферринов и является преобладающим белком, обнаруживаемым в экзокринных секретах млекопитающих, таких как слезы, молоко и слюна. Он состоит приблизительно из 700 остатков и существует в основном в виде тетрамера, причем соотношение мономер: тетрамер составляет 1: 4 при концентрации белка 10 мкМ.[17] Третичная структура состоит из двух долей, называемых долями N и C, каждая из которых содержит один карман для связывания железа. Каждый карман содержит четыре аминокислоты (два тирозина, один гистидин и один аспартат) и, вместе с двумя карбонатными или бикарбонатными анионами, образует шестичленную координату вокруг катиона железа. Именно эта специфическая комбинация делает лактоферрин сродством к железу в 300 раз больше, чем трансферрин.[18]

Лактоферрин обладает значительными антимикробными свойствами. Его самая высокая концентрация - 150 нг / мл - содержится в молозиве человека (тип молока, вырабатываемого на последних стадиях беременности), обеспечивая столь необходимую иммунную поддержку новорожденным.[19] Было широко распространено мнение, что лактоферрин является только бактериостатическим агентом из-за его высокого сродства к железу и его способности изолировать свободные атомы железа от патогенных микробов. Однако теперь известно, что основная движущая сила противомикробного действия заключается в бактерицидных свойствах связанного с железом кармана и специфического пептида лактоферрицина, расположенного в N-доле. Лактоферрин способен связываться с ЛПС (липополисахарид ) слой бактерий, и в его голо форме атом железа окисляет липополисахариды, лизируя внешнюю мембрану и одновременно производя токсичный перекись водорода.[20] Кроме того, при расщеплении лактоферрина трипсином образуется пептид лактоферрицин, который связывается с H+-АТФаза, нарушающая транслокацию протонов и в конечном итоге убивающая клетку.[21]
Ферритин

Ферритин - это железный резервуар для отдельной клетки. Он обнаружен во всех типах клеток и локализуется в цитозоле. Ферритин - это большой белок, состоящий из 24 субъединиц, окружающих ядро, полное атомов железа. Он способен удерживать 0-4500 атомов железа,[22] который можно использовать как резервуар для клеточных нужд. Железо хранится в избытке и извлекается, когда железо снова требуется.[15] Субъединицы представляют собой смесь H (тяжелого или сердечного) и L (легкого или печеночного). Субъединицы образуют кластер шириной 70-80 ангстрем, который затем заполняется ферригидритом железа.[23]
Ферритин - это высококонсервативный белок во всех сферах жизни. Он настолько консервативен, что субъединицы лошадей и людей могут собираться вместе в функциональный белок.[15] Каждая субъединица состоит из пяти альфа-спиралей.
Ферритин используется для диагностики низкого уровня железа у людей.[22] Его можно использовать для определения уровня биодоступного железа, что полезно для диагностики анемии. Обычный диапазон для мужчин составляет 18–270 нг / мл, а для женщин - 18–160 нг / мл.[24]
Смотрите также
внешние ссылки
- Железосвязывающие + белки в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
использованная литература
- ^ Брок Дж. Х. (1989). «Железосвязывающие белки». Acta Paediatrica Scandinavica. Дополнение. 361: 31–43. Дои:10.1111 / apa.1989.78.s361.31. PMID 2485582. S2CID 44752615.
- ^ Де Соуза М., Бридвелт Ф., Динезиус-Трентам Р., Трентэм Д., Лам Дж. (1988). «Железо, железосвязывающие белки и клетки иммунной системы». Летопись Нью-Йоркской академии наук. 526 (1): 310–22. Bibcode:1988НЯСА.526..310С. Дои:10.1111 / j.1749-6632.1988.tb55515.x. PMID 3291685. S2CID 12756539.
- ^ Каплан Дж., DM Ward (август 2013 г.). «Сущность использования и регулирования железа». Текущая биология. 23 (15): R642-6. Дои:10.1016 / j.cub.2013.05.033. ЧВК 3928970. PMID 23928078.
- ^ Соломон Э.И., Брунольд Т.С., Дэвис М.И., Кемсли Дж. Н., Ли С. К., Ленерт Н., Низ Ф., Скулан А. Дж., Ян Ю. С., Чжоу Дж. (Январь 2000 г.). «Геометрическая и электронная структура / взаимосвязь функций в негемовых ферментах железа». Химические обзоры. 100 (1): 235–350. Дои:10.1021 / cr9900275. PMID 11749238.
- ^ Cheng AX, Han XJ, Wu YF, Lou HX (январь 2014 г.). «Функция и катализ 2-оксоглутарат-зависимых оксигеназ, участвующих в биосинтезе флавоноидов растений». Международный журнал молекулярных наук. 15 (1): 1080–95. Дои:10.3390 / ijms15011080. ЧВК 3907857. PMID 24434621.
- ^ а б Аббаспур Н., Харрелл Р., Келишади Р. (февраль 2014 г.). «Обзор железа и его значения для здоровья человека». Журнал исследований в области медицинских наук. 19 (2): 164–74. ЧВК 3999603. PMID 24778671.
- ^ Monsen ER, Hallberg L, Layrisse M, Hegsted DM, Cook JD, Mertz W., Finch CA (январь 1978 г.). «Оценка доступного пищевого железа». Американский журнал клинического питания. 31 (1): 134–41. Дои:10.1093 / ajcn / 31.1.134. PMID 619599.
- ^ «Поглощение железа». course.washington.edu. Получено 2018-11-17.
- ^ Менис, Оскар (1968). «Техническая записка 454». Секция аналитической координационной химии: обзор деятельности с июля 1967 г. по июнь 1968 г.. Гейтерсбург, доктор медицины: Отдел аналитической координационной химии Отдел аналитической химии Институт исследования материалов. Дои:10.6028 / nbs.tn.454.
- ^ Михайлеску М.Р., Руссу И.М. (март 2001 г.). «Признак перехода T ---> R в гемоглобине человека». Труды Национальной академии наук Соединенных Штатов Америки. 98 (7): 3773–7. Bibcode:2001ПНАС ... 98.3773М. Дои:10.1073 / pnas.071493598. ЧВК 31128. PMID 11259676.
- ^ Берг Дж. М., Тимочко Дж. Л., Страйер Л. (2002). Биохимия (5-е изд.). Нью-Йорк: W.H. Фримен. ISBN 0716730510. OCLC 48055706.
- ^ Хассельбалх К.А. (декабрь 1964 г.). «Расчет концентрации ионов водорода в крови в зависимости от связывания свободного и связанного диоксида углерода и кислорода в зависимости от Ph». Обзор анестезиологии. 8 (6): 607–32. Дои:10.1097/00132586-196412000-00059.
- ^ Л., Ленингер, Альберт (2000). Принципы биохимии Ленингера. Нельсон, Дэвид Л. (David Lee), 1942-, Кокс, Майкл М. (3-е изд.). Нью-Йорк: Worth Publishers. ISBN 1572591536. OCLC 42619569.
- ^ Джонсон, Дебора С.; Дин, Деннис Р .; Смит, Арчер Д .; Джонсон, Майкл К. (18 февраля 2005 г.). «Структура, функции и формирование биологических кластеров железа и серы». Ежегодный обзор биохимии. 74 (1): 247–281. Дои:10.1146 / annurev.biochem.74.082803.133518. ISSN 0066-4154. PMID 15952888.
- ^ а б c Айзен П., Эннс С., Весслинг-Резник М. (октябрь 2001 г.). «Химия и биология метаболизма железа в эукариотах». Международный журнал биохимии и клеточной биологии. 33 (10): 940–59. Дои:10.1016 / с 1357-2725 (01) 00063-2. PMID 11470229.
- ^ «TF - предшественник серотрансферрина - Homo sapiens (человек) - ген и белок TF». www.uniprot.org. Получено 2018-11-11.
- ^ Бейкер Э. Н., Бейкер Х. М. (ноябрь 2005 г.). «Молекулярная структура, связывающие свойства и динамика лактоферрина». Клеточные и молекулярные науки о жизни. 62 (22): 2531–9. Дои:10.1007 / s00018-005-5368-9. PMID 16261257.
- ^ Мазурье Дж., Спик Дж. (Май 1980 г.). «Сравнительное исследование железосвязывающих свойств трансферринов человека. I. Полное и последовательное насыщение железом и десатурация лактотрансферрина». Biochimica et Biophysica Acta. 629 (2): 399–408. Дои:10.1016/0304-4165(80)90112-9. PMID 6770907.
- ^ Санчес Л., Кальво М., Брок Дж. Х. (май 1992 г.). «Биологическая роль лактоферрина». Архив детских болезней. 67 (5): 657–61. Дои:10.1136 / adc.67.5.657. ЧВК 1793702. PMID 1599309.
- ^ Фарно С., Эванс Р. В. (ноябрь 2003 г.). «Лактоферрин - многофункциональный белок с антимикробными свойствами». Молекулярная иммунология. 40 (7): 395–405. Дои:10.1016 / S0161-5890 (03) 00152-4. PMID 14568385.
- ^ Kuwata H, Yip TT, Yip CL, Tomita M, Hutchens TW (апрель 1998 г.). «Бактерицидный домен лактоферрина: обнаружение, количественное определение и характеристика лактоферрицина в сыворотке с помощью аффинной масс-спектрометрии SELDI». Сообщения о биохимических и биофизических исследованиях. 245 (3): 764–73. Дои:10.1006 / bbrc.1998.8466. PMID 9588189.
- ^ а б Эндрюс NC (июль 2008 г.). «Ковка поля: золотой век биологии железа». Кровь. 112 (2): 219–30. Дои:10.1182 / кровь-2007-12-077388. ЧВК 2442739. PMID 18606887.
- ^ Crichton RR, Charloteaux-Wauters M (май 1987 г.). «Транспортировка и хранение железа». Европейский журнал биохимии. 164 (3): 485–506. Дои:10.1111 / j.1432-1033.1987.tb11155.x. PMID 3032619.
- ^ «Что такое анализ крови на ферритин? Что означают результаты?». WebMD. Получено 2018-11-11.