Trp оперон - Trp operon
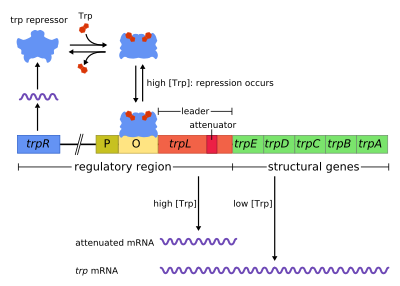
В trp оперон является оперон - группа генов, которые используются или транскрибируются вместе, - кодирующие компоненты для производства триптофан. В trp оперон присутствует во многих бактерии, но впервые был охарактеризован в кишечная палочка. Оперон регулируется таким образом, что, когда триптофан присутствует в окружающей среде, гены синтеза триптофана не экспрессируются. Это была важная экспериментальная система для изучения регуляции генов, и она обычно используется для обучения регуляции генов.
Оперон Trp содержит пять структурных генов: trpE, trpD, trpC, trpB и trpA, которые кодируют ферментативные части пути. Он также содержит репрессивный регуляторный ген называется trpR. trpR имеет промотор, в котором РНК-полимераза связывается и синтезирует мРНК для регуляторного белка. Белок, который синтезируется trpR, затем связывается с оператор что затем вызывает блокировку транскрипции. в trp оперон, триптофан связывается с белком-репрессором, эффективно блокируя транскрипцию гена. В этой ситуации репрессия - это репрессия РНК-полимеразы, транскрибирующей гены оперона. Также в отличие от лак оперон, то trp оперон содержит лидерный пептид и аттенюатор последовательность, которая позволяет ступенчатое регулирование.[1]
Это пример подавляемое отрицательное регулирование экспрессии генов. В регуляторной последовательности оперона оператор привязан к репрессор белок в присутствии триптофана (тем самым предотвращая транскрипция ) и высвобождается в отсутствие триптофана (тем самым позволяя транскрипцию).
Гены
Оперон Trp содержит пять структурных генов. Их роли:
- TrpE (P00895): Антранилатсинтаза производит антранилат.
- TrpD (P00904): Сотрудничает с TrpE.
- TrpC (P00909): Фосфорибозилантранилат изомераза домен сначала превращает N- (5-фосфо-β-D-рибозил) антранилат в 1- (2-карбоксифениламино) -1-дезокси-D-рибулозо-5-фосфат. В Индол-3-глицеринфосфатсинтаза на том же белке затем превращает продукт в (1S, 2R) -1-C- (индол-3-ил) глицерин-3-фосфат.
- TrpA (P0A877), TrpB (P0A879): две субъединицы триптофансинтетаза. Объединяет продукт TrpC с серином для производства триптофана.
Репрессия

В оперон действует по механизму отрицательной подавляемой обратной связи. Репрессор для оперона trp продуцируется перед геном trpR, который конститутивно экспрессируется на низком уровне. Синтезированные мономеры trpR объединяются в димеры. Когда триптофан присутствует, эти репрессор триптофана димеры связываются с триптофаном, вызывая изменение конформации репрессора, позволяя репрессору связываться с оператор. Это предотвращает РНК-полимераза от связывания и транскрипции оперона, поэтому триптофан не производится из его предшественника. Когда триптофан отсутствует, репрессор находится в неактивной конформации и не может связываться с областью оператора, поэтому транскрипция не ингибируется репрессором.
Затухание

Затухание это второй механизм отрицательной обратной связи в trp оперон. Система репрессии нацелена на внутриклеточную концентрацию trp, тогда как ослабление реагирует на концентрацию заряженной тРНК.trp.[2] Таким образом, репрессор trpR снижает экспрессию гена, изменяя инициацию транскрипции, в то время как аттенуация делает это, изменяя процесс транскрипции, который уже выполняется.[2] В то время как репрессор TrpR снижает транскрипцию в 70 раз, аттенуация может дополнительно уменьшить ее в 10 раз, что позволяет накопить репрессию примерно в 700 раз.[3] Затухание стало возможным благодаря тому, что в прокариоты (которые не имеют ядро ), рибосомы начинать Идет перевод то мРНК пока РНК-полимераза все еще расшифровка последовательность ДНК. Это позволяет процессу трансляции напрямую влиять на транскрипцию оперона.
В начале транскрибируемых генов trp оперон представляет собой последовательность из не менее 130 нуклеотидов, называемую лидерным транскриптом (trpL; P0AD92).[4] Ли и Янофски (1977) обнаружили, что эффективность ослабления коррелирует со стабильностью вторичной структуры, встроенной в trpL,[5] и две составляющие шпильки структуры терминатора были позже выяснены Оксендером. и другие. (1979).[6] Этот транскрипт включает четыре короткие последовательности, обозначенные 1–4, каждая из которых частично комплементарна следующей. Таким образом, три отдельные вторичные структуры (заколки для волос ) может образовывать: 1–2, 2–3 или 3–4. Гибридизация последовательностей 1 и 2 с образованием структуры 1-2 происходит редко, поскольку РНК-полимераза ожидает присоединения рибосомы, прежде чем продолжить транскрипцию после последовательности 1, однако, если шпилька 1-2 образовалась, это предотвратило бы образование 2–3 строение (но не 3–4). Образование шпилечной петли между последовательностями 2–3 предотвращает образование шпилечной петли между последовательностями 1-2 и 3-4. Структура 3–4 представляет собой прекращение транскрипции последовательность (в большом количестве в G / C и сразу за ней следуют несколько остатков урацила), как только она образуется, РНК-полимераза отделяется от ДНК, и транскрипция структурных генов оперона не может происходить (более подробное объяснение см. ниже). Функциональное значение второй шпильки для терминации транскрипции иллюстрируется уменьшением частоты терминации транскрипции, наблюдаемым в экспериментах, дестабилизирующих центральное спаривание G + C этой шпильки.[5][7][8][9]
Часть кодов стенограммы руководителя для короткого полипептид из 14 аминокислот, названных лидерным пептидом. Этот пептид содержит два соседних остатка триптофана, что необычно, поскольку триптофан является довольно необычной аминокислотой (примерно один из ста остатков в типичном Кишечная палочка белок - триптофан). Нить 1 в trpL охватывает область, кодирующую концевые остатки лидерного пептида: Trp, Trp, Arg, Thr, Ser;[2] Консервация наблюдается в этих 5 кодонах, тогда как мутация вышележащих кодонов не изменяет экспрессию оперона.[2][10][11][12] Если рибосома пытается транслировать этот пептид, в то время как уровни триптофана в клетке низкие, она остановится на любом из двух кодонов trp. Во время остановки рибосома физически экранирует последовательность 1 транскрипта, предотвращая образование вторичной структуры 1-2. Последовательность 2 затем может свободно гибридизоваться с последовательностью 3, чтобы сформировать структуру 2–3, которая затем предотвращает образование шпильки с 3–4 концом, поэтому структура 2–3 называется шпилькой с концевым концом. При наличии структуры 2–3 РНК-полимераза может продолжать транскрибировать оперон. Мутационный анализ и исследования, включающие дополнительные олигонуклеотиды демонстрируют, что стабильность структуры 2–3 соответствует уровню экспрессии оперона.[10][13][14][15] Если уровни триптофана в клетке высоки, рибосома будет транслировать весь лидерный пептид без прерывания и остановится только во время завершения трансляции в точке стоп-кодон. В этот момент рибосома физически экранирует обе последовательности 1 и 2. Последовательности 3 и 4, таким образом, могут свободно образовывать структуру 3–4, которая завершает транскрипцию. Эта терминаторная структура формируется, когда ни одна рибосома не останавливается в непосредственной близости от тандема Trp (т.е. кодона Trp или Arg): либо лидерный пептид не транслируется, либо трансляция проходит плавно вдоль цепи 1 с обильно заряженной tRNAtrp.[2][10] Более того, предполагается, что рибосома блокирует только около 10 нт ниже по течению, таким образом, остановка рибосомы либо в вышестоящем Gly, либо в нижнем по течению Thr, по-видимому, не влияет на формирование терминальной шпильки.[2][10] Конечным результатом является то, что оперон будет транскрибироваться только тогда, когда триптофан недоступен для рибосомы, в то время как транскрипт trpL экспрессируется конститутивно.
Этот механизм затухания подтвержден экспериментально. Во-первых, прямо доказано, что трансляция лидерного пептида и задержка рибосом необходимы для ингибирования терминации транскрипции.[13] Более того, мутационный анализ, дестабилизирующий или нарушающий спаривание оснований шпильки-антитерминатора, приводит к усилению терминации нескольких складок; в соответствии с моделью аттенуации, эта мутация не может уменьшить аттенюацию даже с голодным Trp.[10][13] Напротив, комплементарные олигонуклеотиды, нацеленные на цепь 1, увеличивают экспрессию оперона, способствуя образованию антитерминатора.[10][14] Более того, в гистидиновом опероне компенсаторная мутация показывает, что способность к спариванию нитей 2–3 имеет большее значение, чем их первичная последовательность в ингибировании аттенуации.[10][15]
В аттенуации, где останавливается трансформирующаяся рибосома, определяется, будет ли сформирована оконечная шпилька.[10] Для того чтобы транскрибирующая полимераза одновременно захватила альтернативную структуру, временной масштаб структурной модуляции должен быть сопоставим с масштабом транскрипции.[2] Чтобы гарантировать, что рибосома связывается и начинает трансляцию лидерного транскрипта сразу после его синтеза, в последовательности trpL существует сайт паузы. Достигнув этого сайта, РНК-полимераза приостанавливает транскрипцию и, по-видимому, ожидает начала трансляции. Этот механизм позволяет синхронизировать транскрипцию и трансляцию, ключевой элемент ослабления.
Подобный механизм ослабления регулирует синтез гистидин, фенилаланин и треонин.
Регулирование оперона trp в Bacillus subtilis
Гены оперона Trp расположены в одинаковом порядке у E. coli и Bacillus subtilis.[16] Регуляция оперонов trp у обоих организмов зависит от количества trp, присутствующего в клетке. Однако первичная регуляция биосинтеза триптофана в Б. subtilis происходит через ослабление, а не подавление транскрипции.[17] В Б. subtilis, триптофан связывается с одиннадцатимпонентным триптофаном-активируемым РНК-связывающим белком ослабления (TRAP), который активирует способность TRAP связываться с лидерной РНК trp.[18][19] Связывание активированного trp TRAP с лидерной РНК приводит к образованию структуры терминатора, которая вызывает терминацию транскрипции.[17]
Рекомендации
- ^ Клуг В.С., Каммингс М.Р., Спенсер С. (2006). Концепции генетики (8-е изд.). Нью-Джерси: Pearson Education Inc., стр. 394–402. ISBN 978-0-13-191833-7.
- ^ а б c d е ж грамм Яновский Ц. (февраль 1981 г.). «Аттенуация в контроле экспрессии бактериальных оперонов». Природа. 289 (5800): 751–8. Bibcode:1981Натура.289..751л. Дои:10.1038 / 289751a0. PMID 7007895. S2CID 4364204.
- ^ Ленингер А.Л., Нельсон Д.Л., Кокс М.М. (2008). Принципы биохимии (5-е изд.). Нью-Йорк, штат Нью-Йорк: W.H. Фримен и компания. п.1128. ISBN 978-0-7167-7108-1.
- ^ Бертран К., Сквайрс К., Янофски С. (май 1976 г.). «Прерывание транскрипции in vivo в лидерной области триптофанового оперона Escherichia coli». Журнал молекулярной биологии. 103 (2): 319–37. Дои:10.1016/0022-2836(76)90315-6. PMID 781269.
- ^ а б Ли Ф., Янофски С. (октябрь 1977 г.). «Терминация транскрипции на аттенюаторах оперона trp Escherichia coli и Salmonella typhimurium: вторичная структура РНК и регуляция терминации». Труды Национальной академии наук Соединенных Штатов Америки. 74 (10): 4365–9. Bibcode:1977PNAS ... 74.4365L. Дои:10.1073 / pnas.74.10.4365. ЧВК 431942. PMID 337297.
- ^ Оксендер Д.Л., Журавски Г., Янофски С. (ноябрь 1979 г.). «Аттенуация в опероне триптофана Escherichia coli: роль вторичной структуры РНК с участием области кодона триптофана». Труды Национальной академии наук Соединенных Штатов Америки. 76 (11): 5524–8. Bibcode:1979PNAS ... 76.5524O. Дои:10.1073 / пнас.76.11.5524. ЧВК 411681. PMID 118451.
- ^ Стройновский I, Яновский C (июль 1982 г.). «Вторичные структуры транскрипта регулируют терминацию транскрипции на аттенюаторе триптофанового оперона S. marcescens». Природа. 298 (5869): 34–8. Bibcode:1982Натура.298 ... 34С. Дои:10.1038 / 298034a0. PMID 7045685. S2CID 4347442.
- ^ Журавски Г., Яновский С. (сентябрь 1980 г.). «Мутации лидера триптофанового оперона Escherichia coli, которые облегчают терминацию транскрипции, являются цис-доминантными по отношению к лидерным мутациям trp, которые усиливают терминацию транскрипции». Журнал молекулярной биологии. 142 (1): 123–9. Дои:10.1016/0022-2836(80)90210-7. PMID 6159477.
- ^ Штауфер Г.В., Журавски Г., Янофски С. (октябрь 1978 г.). «Изменения одной пары оснований в лидерной области оперона trp Escherichia coli, которые снимают терминацию транскрипции на аттенюаторе trp». Труды Национальной академии наук Соединенных Штатов Америки. 75 (10): 4833–7. Bibcode:1978PNAS ... 75.4833S. Дои:10.1073 / пнас.75.10.4833. ЧВК 336215. PMID 368800.
- ^ а б c d е ж грамм час Кольтер Р., Яновский С. (1982). «Аттенуация в оперонах биосинтеза аминокислот». Ежегодный обзор генетики. 16: 113–34. Дои:10.1146 / annurev.ge.16.120182.000553. PMID 6186194.
- ^ Ли Ф, Бертран К., Беннетт Дж., Янофски С. (май 1978 г.). «Сравнение нуклеотидных последовательностей исходных транскрибируемых областей триптофановых оперонов Escherichia coli и Salmonella typhimurium». Журнал молекулярной биологии. 121 (2): 193–217. Дои:10.1016 / с0022-2836 (78) 80005-9. PMID 351195.
- ^ Миоццари Г., Янофски С. (ноябрь 1978 г.). «Встречающаяся в природе отрицательная мутация промотора: нуклеотидная последовательность промотора / оператора / лидерной области trp Shigella dysenteriae 16». Труды Национальной академии наук Соединенных Штатов Америки. 75 (11): 5580–4. Bibcode:1978ПНАС ... 75.5580М. Дои:10.1073 / пнас.75.11.5580. ЧВК 393010. PMID 364484.
- ^ а б c Zurawski G, Elseviers D, Stauffer GV, Yanofsky C (декабрь 1978 г.). «Трансляционный контроль терминации транскрипции на аттенюаторе триптофанового оперона Escherichia coli». Труды Национальной академии наук Соединенных Штатов Америки. 75 (12): 5988–92. Bibcode:1978PNAS ... 75.5988Z. Дои:10.1073 / pnas.75.12.5988. ЧВК 393102. PMID 366606.
- ^ а б Винклер М.Э., Маллис К., Барнетт Дж., Стройновский И., Янофски С. (апрель 1982 г.). «Терминация транскрипции в аттенюаторе триптофанового оперона снижается in vitro олигомером, комплементарным сегменту лидерного транскрипта». Труды Национальной академии наук Соединенных Штатов Америки. 79 (7): 2181–5. Bibcode:1982PNAS ... 79.2181W. Дои:10.1073 / pnas.79.7.2181. ЧВК 346154. PMID 6179092.
- ^ а б Джонстон Х.М., Рот-младший (февраль 1981 г.). «Изменения последовательности ДНК мутаций, изменяющих контроль ослабления гистидинового оперона Salmonella typhimurium». Журнал молекулярной биологии. 145 (4): 735–56. Дои:10.1016/0022-2836(81)90312-0. PMID 6167727.
- ^ Мерино Э., Дженсен Р.А., Янофски С. (апрель 2008 г.). «Эволюция бактериальных trp-оперонов и их регуляция». Текущее мнение в микробиологии. 11 (2): 78–86. Дои:10.1016 / j.mib.2008.02.005. ЧВК 2387123. PMID 18374625.
- ^ а б Голлник П., Бабицке П., Антсон А., Янофски С. (2005-11-14). «Сложность регуляции биосинтеза триптофана у Bacillus subtilis». Ежегодный обзор генетики. 39 (1): 47–68. Дои:10.1146 / annurev.genet.39.073003.093745. PMID 16285852.
- ^ Эллиотт МБ, Готтлиб П.А., Голлник П. (январь 2001 г.). «Механизм связывания РНК с TRAP: инициация и кооперативные взаимодействия». РНК. 7 (1): 85–93. Дои:10.1017 / S135583820100173X. ЧВК 1370072. PMID 11214184.
- ^ Антсон А.А., Отридж Дж., Бжозовски А.М., Додсон Э.Дж., Додсон Г.Г., Уилсон К.С. и др. (Апрель 1995 г.). «Структура trp РНК-связывающего белка аттенуации». Природа. 374 (6524): 693–700. Bibcode:1995Натура 374..693А. Дои:10.1038 / 374693a0. PMID 7715723. S2CID 4340136.
дальнейшее чтение
- Морзе Д.Е., Мостеллер Р.Д., Яновский С. (1969). «Динамика синтеза, трансляции и деградации матричной РНК оперона trp в E. coli». Симпозиумы Колд-Спринг-Харбор по количественной биологии. 34: 725–40. Дои:10.1101 / sqb.1969.034.01.082. PMID 4909527.
- Яновский Ц. (февраль 1981 г.). «Аттенуация в контроле экспрессии бактериальных оперонов». Природа. 289 (5800): 751–8. Bibcode:1981Натура.289..751л. Дои:10.1038 / 289751a0. PMID 7007895. S2CID 4364204.