Деметилаза - Demethylase
Деметилазы находятся ферменты это удалить метил (CH3-) группы из нуклеиновые кислоты, белки (в частности гистоны ) и другие молекулы. Ферменты деметилазы важны в эпигенетический механизмы модификации. Белки деметилазы изменяют транскрипционная регуляция из геном контролируя метилирование уровни, которые возникают на ДНК и гистонах и, в свою очередь, регулируют хроматин заявить на конкретных ген loci внутри организмов.
Окислительное деметилирование
Деметилирование гистонов

На протяжении многих лет метилирование гистонов считалось необратимым из-за того, что период полужизни метилирования гистонов был приблизительно равен периоду полужизни самих гистонов.[1] В 2004 году Shi et al. опубликовали свое открытие гистон-деметилазы LSD1 (позже классифицированной как KDM1A ), гомолог ядерной аминоксидазы.[2] С тех пор было обнаружено еще много гистоновых деметилаз. В зависимости от их механизмов существуют два основных класса гистоновых деметилаз: флавинадениндинуклеотид (FAD) -зависимый амин оксидаза, и Fe (II) и α-кетоглутарат-зависимая гидроксилаза. Оба действуют путем гидроксилирования метильной группы с последующей диссоциацией формальдегида. Деметилирование имеет значение для эпигенетика.[3]
Белки гистон-деметилазы имеют множество домены которые выполняют разные функции. Эти функции включают связывание с гистоном (или иногда ДНК на нуклеосома ), распознавая правильный метилированный аминокислота субстрат, катализирующий реакцию, и связывающие кофакторы. Кофакторы включают: альфа-кетоглутарат (JmjC-домен, содержащий деметилазы), CoREST (LSD), FAD, Fe (II) или NOG (N-оксалилглицин).[4] Домены включают:
- SWIRM1 (домен Swi3, Rsc и Мойра): Предлагаемый якорный сайт для молекул гистонов; найдено в нескольких хроматин модифицирующие комплексы; способствует развитию белка деметилазы и кофактора CoREST (связывание с нуклеосомным субстратом)[5]
- Jumonji (N / C терминальные домены): Связывающий домен ключа кофакторы Такие как альфа-кетоглутарат; связаны бета-шпилька / смешанный домен[4][6]
- PHD-палец: гидрофобная клетка остатков, которая связывает метилированные пептиды; играет ключевую роль в распознавании и селективности метилированных остатков гистонов[4]
- Цинковый палец: ДНК-связывающий домен[4]
- Аминоксидаза домен: каталитический активный центр белков LSD; больше, чем родственные белки, чтобы соответствовать размеру гистонового хвоста [4]


Есть несколько семейств гистоновых деметилаз, которые действуют на разные субстраты и играют разные роли в клеточной функции. Был разработан код для обозначения субстрата гистоновой деметилазы. Субстрат сначала определяется субъединицей гистона (H1, H2A, H2B, H3, H4), а затем однобуквенным обозначением и номером аминокислота то есть метилированный. Наконец, уровень метилирования иногда отмечается добавлением «me #» с номерами 1, 2 и 3 для монометилированных, диметилированных и триметилированных субстратов соответственно. Например, H3K9me2 - это гистон H3 с диметилированным лизином в девятом положении.
- KDM1
- Семейство KDM1 включает KDM1A и KDM1B. KDM1A (также называемый LSD1 / AOF2 / BHC110) может действовать на моно- и диметилированный H3K4 и H3K9, а KDM1B (также называемый LSD2 / AOF1) действует только на моно- и диметилированный H3K4. Эти ферменты могут играть решающую роль в эмбриогенез и тканеспецифичные дифференциация, а также рост ооцитов.[1] KDM1A была первой открытой деметилазой, и поэтому она изучена наиболее широко.[2]
- Делеция гена KDM1A может влиять на рост и дифференцировку эмбриональные стволовые клетки и может привести к эмбриональному летальность в нокаутные мыши, которые не продуцируют продукт гена KDM1A.[7][8] Считается, что KDM1A также играет роль в развитии рака, поскольку более неблагоприятные исходы могут быть коррелированы с более высокой экспрессией этого гена.[9][10] Следовательно, ингибирование KDM1A может быть возможным лечением рака.[11][12] KDM1A имеет много разных партнеров связывания, которые могут быть необходимы для его активности деметилирования.[13]
- KDM1B, однако, в основном участвует в ооцит разработка. Удаление этого гена приводит к материнский эффект летальность у мышей.[14] Ортологи KDM1 в D. melanogaster и C. elegans похоже, функционирует аналогично KDM1B, а не KDM1A.[15][16]
- KDM2
- Семейство KDM2 включает KDM2A и KDM2B. KDM2A (также известный как JHDM1A / FBXL11) может действовать на моно- и диметилированный H3K36 и триметилированный H3K4. KDM2B (также называемый JHDM1B / FBXL10) действует только на моно- и диметилированный H3K36. KDM2A играет роль либо в стимулировании, либо в подавлении функции опухоли, а KDM2B играет роль в онкогенез.[1]
- У многих эукариот белок KDM2A содержит домен цинкового пальца CXXC, способный связывать неметилированные CpG-островки. В настоящее время считается, что белки KDM2A могут связываться со многими регуляторными элементами генов без помощи специфичных для последовательности факторов транскрипции.[17] Хотя роль KDM2 в дифференцировке развития эукариот все еще остается загадкой, было показано, что как KDM2A, так и KDM2B играют роль в росте и подавлении опухолей. Было показано, что KDM2B чрезмерно экспрессируется у человека. лимфомы и аденокарциномы; однако при раке предстательной железы и глиобластомах наблюдается сниженная экспрессия как KDM2A, так и KDM2B. Кроме того, было показано, что KDM2B предотвращает старение в некоторых камерах через эктопическое выражение дальнейшее указание на его потенциал как онкогена.[18]
- KDM3
- Семейство KDM3 включает KDM3A, KDM3B и JMJD1C. KDM3A (также называемый JHDM2A / JMJD1A / TSGA) может действовать на моно- и диметилированный H3K9. Подложки для KDM3B (также обозначаемые как JHDM2B / JMJD1B) и JMJD1C (также обозначаемые как JHDM2C / TRIP8) неизвестны.[19] KDM3A играет роль в сперматогенез и метаболические функции; роли KDM3B и JMJD1C неизвестны.[1]
- Нокдаун-исследования KDM3A на мышах, где мышь продуцирует пониженные уровни KDM3A, привели к мужскому бесплодию и ожирению у взрослых. Дополнительные исследования показали, что KDM3A может играть роль в регуляции генов, зависимых от рецепторов андрогенов, а также генов, участвующих в плюрипотентность, что указывает на потенциальную роль KDM3A в онкогенезе.[20]
- KDM4
- Семейство KDM4 включает KDM4A, KDM4B, KDM4C, и KDM4D. Они также называются JMDM3A / JMJD2A, JMDM3B / JMJD2B, JMDM3C / JMJD2C и JMDM3D / JMJD2D соответственно. Эти ферменты могут действовать на ди- и триметилированные H3K9, H3K36, H1K26. KDM4B и KDM4C играют роль в туморогенез, а роль KDM4D неизвестна.[1]
- Семейство белков KDM4 связано со злокачественной трансформацией. В частности, амплификация KDM4C была задокументирована при плоскоклеточном раке пищевода, медуллобластоме и раке груди; Амплификация KDM4B также была обнаружена в медуллобластомах.[21][22][23][24] Другие данные по экспрессии генов также предполагают, что KDM4A, KDM4B и KDM4C чрезмерно экспрессируются при раке простаты.[25]
- KDM5
- Семейство KDM5 включает KDM5A, KDM5B, KDM5C, и KDM5D. Их также называют JARID1A / RBP2, JARID1B / PLU-1, JARID1C / SMCX и JARID1D / SMCY соответственно. Эти ферменты могут действовать на ди- и триметилированный H3K4.[1]
- Семейство белков KDM5, по-видимому, играет ключевые функции развития. Удаление домена JmjC ретинобластомы связывающего белка, родственного 2 (RBR-2) в C. elegans выраженные дефекты формирования вульвы.[26] Мутации домена JmjC в Дрозофила вызывает либо смертельные последствия для личинок, либо многие дефекты развития у выживших.[27]
- KDM5A в системах клеточных культур также продемонстрировал связь с регуляцией дифференцировки, митохондриальный функция клеточный цикл прогрессия.[28][29][30][31][32][33] KDM5B и KDM5C также показали взаимодействие с белками PcG, которые участвуют в репрессии транскрипции. Мутации KDM5C (обнаруженные на Х-хромосоме) также были обнаружены у пациентов с Х-связанный умственная отсталость.[34] Истощение гомологов KDM5C в D. rerio показали дефекты формирования паттерна мозга и гибель нейрональных клеток.[35]
- KDM6
- Семейство KDM6 включает KDM6A, KDM6B, и UTY. KDM6A (также обозначаемый как UTX) и KDM6B (также обозначаемый как JMJD3) действуют на ди- и триметилированный H3K27 и играют роль в развитии; субстрат и роль UTY неизвестны.[1] В целом, и KDM6A, и KDM6B обладают опухолевыми характеристиками. KDM6A нокдауны в фибробласты привести к немедленному увеличению популяции фибробластов. KDM6B, экспрессируемый в фибробластах, индуцирует онкогены пути RAS_RAF.[36] Делеции и точечные мутации KDM6A были идентифицированы как одна из причин Синдром Кабуки, врожденное нарушение, приводящее к умственной отсталости.[37][38]
- Для KDM6B были предложены другие возможные роли. В частности, в одном исследовании мутирующие гомологи KDM6B нарушали развитие гонад у C.elegans.[39] Другие исследования показали, что экспрессия KDM6B активируется в активированных макрофаги и динамически выражается при дифференциации стволовые клетки.[40][41]
- С другой стороны, истощение гомологов KDM6A в D. rerio показали снижение экспрессии генов HOX, которые играют роль в регуляции рисунок тела во время разработки.[42] В исследованиях на млекопитающих было показано, что KDM6A также регулирует гены HOX.[39][43]
Деметилирование сложного эфира

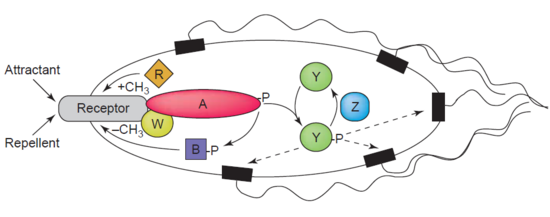
Другой пример деметилазы - это протеин-глутаматметилэстераза, также известный как белок CheB (EC 3.1.1.61), который деметилирует MCP (мэтил-принимающий cгемотаксис пroteins) посредством гидролиза сложноэфирных связей карбоновых кислот. Ассоциация хемотаксис рецептор с агонистом приводит к фосфорилированию CheB. Фосфорилирование белка CheB усиливает его каталитическую активность деметилирования МСР, что приводит к адаптации клетки к раздражителям окружающей среды.[44] МСК реагируют на внеклеточные аттрактанты и репелленты у бактерий, таких как Кишечная палочка в хемотаксис регулирование. CheB более конкретно называют метилэстераза, так как он удаляет метильные группы из метилглутамат остатки, расположенные на MCP в результате гидролиза, с образованием глутамат сопровождается выпуском метанол.[45]
CheB представляет особый интерес для исследователей, поскольку может быть терапевтической мишенью для смягчения распространения бактериальных инфекций.[46]
Смотрите также
Рекомендации
- ^ а б c d е ж грамм Педерсен М.Т., Хелин К. (ноябрь 2010 г.). «Гистоновые деметилазы в развитии и болезни». Тенденции в клеточной биологии. 20 (11): 662–71. Дои:10.1016 / j.tcb.2010.08.011. PMID 20863703.
- ^ а б Ши Ю., Лан Ф., Матсон С., Маллиган П., Ветстайн Дж. Р., Коул П.А., Касеро Р.А., Ши И. (декабрь 2004 г.). «Деметилирование гистонов, опосредованное ядерным гомологом аминоксидазы LSD1». Клетка. 119 (7): 941–53. Дои:10.1016 / j.cell.2004.12.012. PMID 15620353.
- ^ Klose RJ, Zhang Y (апрель 2007 г.). «Регулирование метилирования гистонов путем деметилирования и деметилирования». Обзоры природы Молекулярная клеточная биология. 8 (4): 307–18. Дои:10.1038 / nrm2143. PMID 17342184.
- ^ а б c d е Мосаммапараст Н, Ши Й (2010). «Обращение метилирования гистонов: биохимические и молекулярные механизмы гистоновых деметилаз». Ежегодный обзор биохимии. 79: 155–79. Дои:10.1146 / annurev.biochem.78.070907.103946. PMID 20373914.
- ^ Tochio N, Umehara T, Koshiba S, Inoue M, Yabuki T, Aoki M, Seki E, Watanabe S, Tomo Y, Hanada M, Ikari M, Sato M, Terada T, Nagase T, Ohara O, Shirouzu M, Tanaka A , Кигава Т., Ёкояма С. (март 2006 г.). «Структура раствора домена SWIRM гистоновой деметилазы LSD1 человека». Структура. 14 (3): 457–68. Дои:10.1016 / j.str.2005.12.004. PMID 16531230.
- ^ Апреликова О., Чен К., Эль Туни Л.Х., Бриньяц-Гиттард С., Хан Дж., Цю Т., Ян Х.Х., Ли М.П., Чжу М., Грин Дж. Э. (апрель 2016 г.). «Эпигенетический модификатор JMJD6 амплифицируется в опухолях молочной железы и взаимодействует с c-Myc для усиления клеточной трансформации, прогрессирования опухоли и метастазирования». Clin Epigenetics. 8 (38). Дои:10.1186 / s13148-016-0205-6. ЧВК 4831179. PMID 27081402.
- ^ Ван Дж., Хеви С., Кураш Дж. К., Лей Х., Гей Ф., Байко Дж., Су Х, Сун В., Чанг Х, Сюй Г., Гаудет Ф, Ли Э, Чен Т. (январь 2009 г.). «Лизиндеметилаза LSD1 (KDM1) необходима для поддержания глобального метилирования ДНК». Природа Генетика. 41 (1): 125–9. Дои:10,1038 / нг.268. PMID 19098913.
- ^ Ван Дж., Скалли К., Чжу Х, Цай Л., Чжан Дж., Префонтейн Г. Г., Кронес А., Охги К. А., Чжу П., Гарсия-Бассет И., Лю Ф., Тейлор Х., Лозач Дж., Джейс Флорида, Корах К. С., Гласс К. К., Fu XD, Розенфельд MG (апрель 2007 г.). «Противоположные комплексы LSD1 действуют в программах активации и репрессии генов развития». Природа. 446 (7138): 882–7. Дои:10.1038 / природа05671. PMID 17392792.
- ^ Kahl P, Gullotti L, Heukamp LC, Wolf S, Friedrichs N, Vorreuther R, Solleder G, Bastian PJ, Ellinger J, Metzger E, Schüle R, Buettner R (декабрь 2006 г.). «Коактиваторы рецептора андрогенов, лизин-специфическая гистоновая деметилаза 1 и четыре с половиной домена LIM-белка 2, предсказывают риск рецидива рака простаты». Исследования рака. 66 (23): 11341–7. Дои:10.1158 / 0008-5472.CAN-06-1570. PMID 17145880.
- ^ Лим С., Янцер А., Беккер А., Циммер А., Шюле Р., Бюттнер Р., Кирфель Дж. (Март 2010 г.). «Лизин-специфическая деметилаза 1 (LSD1) высоко экспрессируется при ER-негативном раке молочной железы и является биомаркером, предсказывающим агрессивную биологию». Канцерогенез. 31 (3): 512–20. Дои:10.1093 / carcin / bgp324. PMID 20042638.
- ^ Мецгер Э., Виссманн М., Инь Н., Мюллер Дж. М., Шнайдер Р., Петерс А. Х., Гюнтер Т., Бюттнер Р., Шюле Р. (сентябрь 2005 г.). «LSD1 деметилирует репрессивные гистоновые метки, чтобы способствовать транскрипции, зависимой от андрогенных рецепторов». Природа. 437 (7057): 436–9. Дои:10.1038 / природа04020. PMID 16079795.
- ^ Schulte JH, Lim S, Schramm A, Friedrichs N, Koster J, Versteeg R, Ora I, Pajtler K, Klein-Hitpass L, Kuhfittig-Kulle S, Metzger E, Schüle R, Eggert A, Buettner R, Kirfel J (Mar 2009 г.). «Лизин-специфическая деметилаза 1 сильно экспрессируется в низкодифференцированной нейробластоме: значение для терапии». Исследования рака. 69 (5): 2065–71. Дои:10.1158 / 0008-5472.CAN-08-1735. PMID 19223552.
- ^ Wang Y, Zhang H, Chen Y, Sun Y, Yang F, Yu W, Liang J, Sun L, Yang X, Shi L, Li R, Li Y, Zhang Y, Li Q, Yi X, Shang Y (август 2009 г.) ). "LSD1 является субъединицей комплекса NuRD и нацелен на программы метастазирования при раке груди". Клетка. 138 (4): 660–72. Дои:10.1016 / j.cell.2009.05.050. PMID 19703393.
- ^ Чикконе Д.Н., Су Х., Хеви С., Гей Ф., Лей Х., Байко Дж., Сюй Г., Ли Э, Чен Т. (сентябрь 2009 г.). «KDM1B представляет собой гистоновую H3K4 деметилазу, необходимую для установления материнских геномных отпечатков». Природа. 461 (7262): 415–8. Дои:10.1038 / природа08315. PMID 19727073.
- ^ Рудольф Т., Йонезава М., Лейн С., Гейдрих К., Кубичек С., Шефер С., Фальке С., Вальтер М., Шмидт А., Йенувейн Т., Рейтер Г. (апрель 2007 г.). «Образование гетерохроматина у дрозофилы инициируется активным удалением метилирования H3K4 гомологом LSD1 SU (VAR) 3-3». Молекулярная клетка. 26 (1): 103–15. Дои:10.1016 / j.molcel.2007.02.025. PMID 17434130.
- ^ Ди Стефано Л., Джи Дж.Й., Мун Н.С., Герр А., Дайсон Н. (май 2007 г.). «Мутация Lsd1 дрозофилы нарушает метилирование H3-K4, что приводит к тканеспецифическим дефектам во время развития». Текущая биология. 17 (9): 808–12. Дои:10.1016 / j.cub.2007.03.068. ЧВК 1909692. PMID 17462898.
- ^ Блэкледж Н.П., Чжоу Джей Си, Толсторуков М.Ю., Фаркас А.М., Парк П.Дж., Клозе Р.Дж. (апрель 2010 г.). «CpG-островки привлекают гистон H3 лизин 36 деметилазу». Молекулярная клетка. 38 (2): 179–90. Дои:10.1016 / j.molcel.2010.04.009. ЧВК 3098377. PMID 20417597.
- ^ Хэ Дж, Каллин Э.М., Цукада Й, Чжан И (ноябрь 2008 г.). «Деметилаза H3K36 Jhdm1b / Kdm2b регулирует пролиферацию и старение клеток через p15 (Ink4b)». Структурная и молекулярная биология природы. 15 (11): 1169–75. Дои:10.1038 / nsmb.1499. ЧВК 2612995. PMID 18836456.
- ^ Ямане К., Тумазоу К., Цукада Ю., Эрдджумент-Бромаж Х, Темпст П., Вонг Дж., Чжан Ю. (май 2006 г.). «JHDM2A, JmjC-содержащая деметилаза H3K9, способствует активации транскрипции рецептором андрогенов». Клетка. 125 (3): 483–95. Дои:10.1016 / j.cell.2006.03.027. PMID 16603237.
- ^ Ло Й.Х., Чжан В., Чен Х, Джордж Дж., Нг Х.Х. (октябрь 2007 г.). «Деметилазы Jmjd1a и Jmjd2c гистона H3 Lys 9 регулируют самообновление в эмбриональных стволовых клетках». Гены и развитие. 21 (20): 2545–57. Дои:10.1101 / gad.1588207. ЧВК 2000320. PMID 17938240.
- ^ Эрбрехт А., Мюллер Ю., Вольтер М., Хойшен А., Кох А., Радлвиммер Б., актер Б., Минчева А., Пич Т., Лихтер П., Райфенбергер Г., Вебер Р. Г. (март 2006 г.). «Комплексный геномный анализ десмопластических медуллобластом: идентификация новых амплифицированных генов и отдельная оценка различных гистологических компонентов». Журнал патологии. 208 (4): 554–63. Дои:10.1002 / path.1925. PMID 16400626.
- ^ Лю Дж., Боллиг-Фишер А., Крайке Б., ван де Виджвер М.Дж., Абрамс Дж., Этье С.П., Ян З.К. (декабрь 2009 г.). «Геномная амплификация и онкогенные свойства гена гистон-деметилазы GASC1 при раке груди». Онкоген. 28 (50): 4491–500. Дои:10.1038 / onc.2009.297. ЧВК 2795798. PMID 19784073.
- ^ Northcott PA, Nakahara Y, Wu X, Feuk L, Ellison DW, Croul S, Mack S, Kongkham PN, Peacock J, Dubuc A, Ra YS, Zilberberg K, McLeod J, Scherer SW, Sunil Rao J, Eberhart CG, Grajkowska У, Гиллеспи И., Лак Б., Гранди Р., Поллак И.Ф., Гамильтон Р.Л., Ван Метер Т., Карлотти К.Г., Буп Ф., Бигнер Д., Гилбертсон Р.Дж., Рутка Д.Т., Тейлор М.Д. (апрель 2009 г.). «Множественные повторяющиеся генетические события сходятся на контроле метилирования гистонового лизина в медуллобластоме». Природа Генетика. 41 (4): 465–72. Дои:10,1038 / нг.336. ЧВК 4454371. PMID 19270706.
- ^ Ян З. К., Имото И., Фукуда Й, Пимхаокхам А., Шимада Й, Имамура М., Сугано С., Накамура Ю., Инадзава Дж. (Сентябрь 2000 г.). «Идентификация нового гена GASC1 в ампликоне 9p23-24, часто обнаруживаемом в клеточных линиях рака пищевода». Исследования рака. 60 (17): 4735–9. PMID 10987278.
- ^ Cloos PA, Christensen J, Agger K, Maiolica A, Rappsilber J, Antal T, Hansen KH, Helin K (июль 2006 г.). «Предполагаемый онкоген GASC1 деметилирует три- и диметилированный лизин 9 на гистоне H3». Природа. 442 (7100): 307–11. Дои:10.1038 / природа04837. PMID 16732293.
- ^ Christensen J, Agger K, Cloos PA, Pasini D, Rose S, Sennels L, Rappsilber J, Hansen KH, Salcini AE, Helin K (март 2007 г.). «RBP2 принадлежит к семейству деметилаз, специфичных для три- и диметилированного лизина 4 на гистоне 3». Клетка. 128 (6): 1063–76. Дои:10.1016 / j.cell.2007.02.003. PMID 17320161.
- ^ Gildea JJ, Lopez R, Shearn A (октябрь 2000 г.). «Скрининг новых генов группы триторакса выявил маленькие имагинальные диски, гомолог Drosophila melanogaster связывающего белка 2 ретинобластомы человека». Генетика. 156 (2): 645–63. ЧВК 1461290. PMID 11014813.
- ^ Ли Н., Эрдджумент-Бромаж Х, Темпст П., Джонс Р.С., Чжан Й. (март 2009 г.). «Крышка деметилазы H3K4 связывает и ингибирует гистондеацетилазу Rpd3». Молекулярная и клеточная биология. 29 (6): 1401–10. Дои:10.1128 / MCB.01643-08. ЧВК 2648242. PMID 19114561.
- ^ Беневоленская Е.В., Мюррей Х.Л., Брантон П., Янг Р.А., Каелин WG (июнь 2005 г.). «Связывание pRB с белком PHD RBP2 способствует клеточной дифференцировке». Молекулярная клетка. 18 (6): 623–35. Дои:10.1016 / j.molcel.2005.05.012. PMID 15949438.
- ^ Лопес-Бигас Н., Кисиэль Т.А., Деваал Д.К., Холмс К.Б., Фолькерт Т.Л., Гупта С., Лав Дж., Мюррей Х.Л., Янг Р.А., Беневоленская Е.В. (август 2008 г.). «Полногеномный анализ гистонодеметилазы H3K4 RBP2 выявляет транскрипционную программу, контролирующую дифференцировку». Молекулярная клетка. 31 (4): 520–30. Дои:10.1016 / j.molcel.2008.08.004. ЧВК 3003864. PMID 18722178.
- ^ Pasini D, Hansen KH, Christensen J, Agger K, Cloos PA, Helin K (май 2008 г.). «Скоординированная регуляция репрессии транскрипции деметилазой RBP2 H3K4 и Polycomb-Repressive Complex 2». Гены и развитие. 22 (10): 1345–55. Дои:10.1101 / gad.470008. ЧВК 2377189. PMID 18483221.
- ^ ван Овелен С., Ван Дж., Асп П., Ян К., Келин В. Г., Клюгер Ю., Dynlacht BD (ноябрь 2008 г.). «Роль Sin3 млекопитающих в постоянном молчании генов». Молекулярная клетка. 32 (3): 359–70. Дои:10.1016 / j.molcel.2008.10.015. ЧВК 3100182. PMID 18995834.
- ^ Цзэн Дж., Гэ З., Ван Л., Ли Кью, Ван Н., Бьоркхольм М., Цзя Дж., Сюй Д. (март 2010 г.). «Гистоновая деметилаза RBP2 сверхэкспрессируется при раке желудка, и ее ингибирование вызывает старение раковых клеток». Гастроэнтерология. 138 (3): 981–92. Дои:10.1053 / j.gastro.2009.10.004. PMID 19850045.
- ^ Дженсен Л.Р., Аменде М., Гурок У., Мозер Б., Гиммель В., Цшах А., Янеке А.Р., Таривердиан Дж., Челли Дж., Фринс Дж. П., Ван Эш Х., Клифстра Т., Хамель Б., Морейн С., Гец Дж., Тернер Дж., Рейнхардт Р., Кальшойер В.М., Роперс Х.Х., Лензнер С. (февраль 2005 г.). «Мутации в гене JARID1C, который участвует в регуляции транскрипции и ремоделировании хроматина, вызывают Х-сцепленную умственную отсталость». Американский журнал генетики человека. 76 (2): 227–36. Дои:10.1086/427563. ЧВК 1196368. PMID 15586325.
- ^ Ивасе С., Лан Ф., Бейлисс П., де ла Торре-Убьета Л., Уарте М., Ци Х. Х., Ветстайн Дж. Р., Бонни А., Робертс TM, Ши Й. (март 2007 г.). «Х-сцепленный ген умственной отсталости SMCX / JARID1C определяет семейство лизин-4 деметилаз гистона H3». Клетка. 128 (6): 1077–88. Дои:10.1016 / j.cell.2007.02.017. PMID 17320160.
- ^ Agger K, Cloos PA, Rudkjaer L, Williams K, Andersen G, Christensen J, Helin K (май 2009 г.). «Деметилаза H3K27me3 JMJD3 способствует активации локуса INK4A-ARF в ответ на старение, вызванное онкогеном и стрессом». Гены и развитие. 23 (10): 1171–6. Дои:10.1101 / gad.510809. ЧВК 2685535. PMID 19451217.
- ^ Ледерер Д., Гризарт Б., Дигилио М.С., Бенуа В., Креспин М., Гарриани С.К., Майштадт I, Даллапиккола Б., Вереллен-Дюмулен С. (январь 2012 г.). «Делеция KDM6A, гистоновой деметилазы, взаимодействующей с MLL2, у трех пациентов с синдромом Кабуки». Американский журнал генетики человека. 90 (1): 119–24. Дои:10.1016 / j.ajhg.2011.11.021. ЧВК 3257878. PMID 22197486.
- ^ Мияке Н., Мидзуно С., Окамото Н., Охаши Н., Шиина М., Огата К., Цурусаки Ю., Накашима М., Сайцу Н., Ниикава Н., Мацумото Н. (январь 2013 г.). «Точечные мутации KDM6A вызывают синдром Кабуки». Человеческая мутация. 34 (1): 108–10. Дои:10.1002 / humu.22229. PMID 23076834.
- ^ а б Аггер К., Клоос П.А., Кристенсен Дж., Пазини Д., Роуз С., Раппсилбер Дж., Исаева И., Канаани Е., Сальчини А. Е., Хелин К. (октябрь 2007 г.). «UTX и JMJD3 представляют собой деметилазы гистона H3K27, участвующие в регуляции и развитии гена HOX». Природа. 449 (7163): 731–4. Дои:10.1038 / природа06145. PMID 17713478.
- ^ De Santa F, Totaro MG, Prosperini E, Notarbartolo S, Testa G, Natoli G (сентябрь 2007 г.). «Гистон H3 лизин-27 деметилаза Jmjd3 связывает воспаление с ингибированием опосредованного поликомбусом сайленсинга генов». Клетка. 130 (6): 1083–94. Дои:10.1016 / j.cell.2007.08.019. PMID 17825402.
- ^ Burgold T, Spreafico F, De Santa F, Totaro MG, Prosperini E, Natoli G, Testa G (2008). «Гистон H3 лизин 27-специфическая деметилаза Jmjd3 необходима для нейронной связи». PLOS ONE. 3 (8): e3034. Дои:10.1371 / journal.pone.0003034. ЧВК 2515638. PMID 18716661.
- ^ Lan F, Bayliss PE, Rinn JL, Whetstine JR, Wang JK, Chen S, Iwase S, Alpatov R, Issaeva I, Canaani E, Roberts TM, Chang HY, Shi Y (октябрь 2007 г.). «Гистон H3 лизин 27 деметилаза регулирует заднее развитие животных». Природа. 449 (7163): 689–94. Дои:10.1038 / природа06192. PMID 17851529.
- ^ Ван Дж. К., Цай М. С., Пулин Дж., Адлер А. С., Чен С., Лю Х., Ши Ю., Чанг ХЙ (Февраль 2010 г.). «Гистоновая деметилаза UTX обеспечивает RB-зависимый контроль судьбы клеток». Гены и развитие. 24 (4): 327–32. Дои:10.1101 / gad.1882610. ЧВК 2816731. PMID 20123895.
- ^ а б Владимиров Н., Левдок Л., Лебедз Д., Суржик В. (декабрь 2008 г.). «Зависимость бактериального хемотаксиса от формы градиента и скорости адаптации». PLoS вычислительная биология. 4 (12): e1000242. Дои:10.1371 / journal.pcbi.1000242. ЧВК 2588534. PMID 19096502.
- ^ Park SY, Borbat PP, Gonzalez-Bonet G, Bhatnagar J, Pollard AM, Freed JH, Bilwes AM, Crane BR (май 2006 г.). «Реконструкция сборки рецептор-киназа хемотаксиса». Структурная и молекулярная биология природы. 13 (5): 400–7. Дои:10.1038 / nsmb1085. PMID 16622408.
- ^ West AH, Martinez-Hackert E, Stock AM (июль 1995 г.). «Кристаллическая структура каталитического домена метилэстеразы рецептора хемотаксиса, CheB». Журнал молекулярной биологии. 250 (2): 276–90. Дои:10.1006 / jmbi.1995.0376. PMID 7608974.