Bacillus subtilis - Bacillus subtilis
| Bacillus subtilis | |
|---|---|
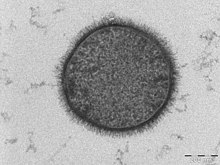 | |
| ТЕМ микрофотография из Б. subtilis ячейка в поперечном сечении (шкала = 200 нм ) | |
| Научная классификация | |
| Домен: | Бактерии |
| Тип: | Фирмикуты |
| Учебный класс: | Бациллы |
| Заказ: | Бациллы |
| Семья: | Bacillaceae |
| Род: | Бациллы |
| Разновидность: | Б. subtilis |
| Биномиальное имя | |
| Bacillus subtilis (Эренберг 1835) Кон 1872 | |
| Синонимы | |
| |
Bacillus subtilis, известный также как сенная палочка или же травяная палочка, это Грамположительный, каталаза -положительный бактерия, содержится в почве и желудочно-кишечный тракт из жвачные животные и люди. Член род Бациллы, Б. subtilis имеет форму стержня и может образовывать прочный, защитный эндоспора, что позволяет ему выдерживать экстремальные условия окружающей среды. Б. subtilis исторически классифицируется как облигатный аэроб, хотя существуют доказательства того, что это факультативный анаэроб. Б. subtilis считается наиболее изученной грамположительной бактерией и модельный организм изучить репликацию бактериальных хромосом и дифференцировку клеток. Это один из бактериальных чемпионов по секретным фермент производятся и используются в промышленных масштабах биотехнологическими компаниями.
Описание
Bacillus subtilis это Грамположительный бактерия стержневидный и каталаза -положительный. Первоначально он назывался Vibrio subtilis к Кристиан Готфрид Эренберг,[3] и переименовал Bacillus subtilis к Фердинанд Кон в 1872 г.[4] (subtilis в переводе с латыни означает «штраф»). Б. subtilis клетки обычно имеют палочковидную форму, имеют длину около 4–10 микрометров (мкм) и диаметр 0,25–1,0 мкм, при этом объем клетки составляет около 4,6 мкл в стационарной фазе.[5] Как и в случае с другими членами род Бациллы, это может сформировать эндоспора, чтобы выжить в экстремальных условиях окружающей среды с температурой и высыханием.[6] Б. subtilis это факультативный анаэроб[7] и считался облигатный аэроб до 1998 года. Б. subtilis сильно бичеванный, что дает ему возможность быстро перемещаться в жидкостях. Б. subtilis оказался очень податливым генетическая манипуляция, и получил широкое распространение как модельный организм для лабораторных исследований, особенно спороношение, который является упрощенным примером клеточная дифференциация. С точки зрения популярности в качестве лабораторной модели организма, Б. subtilis часто рассматривается как Грамположительный эквивалент кишечная палочка, тщательно изученный Грамотрицательный бактерия.[нужна цитата ]
Среда обитания
Этот вид обычно встречается в верхних слоях почвы и Б. subtilis считается нормальный комменсал кишечника в людях. В исследовании 2009 года сравнивалась плотность спор, обнаруженных в почве (около 106 спор на грамм) к количеству в кале человека (около 104 спор на грамм). Количество спор, обнаруженных в кишечнике человека, было слишком большим, чтобы их можно было приписать исключительно употреблению в пищу.[8] Б. subtilis был связан с ростом на возвышенностях и выступал в качестве идентификатора как экологической адаптации, так и здоровья медоносных пчел.[9]
Размножение

Б. subtilis можно разделить симметрично, чтобы получилось два дочерние клетки (бинарное деление), или асимметрично, с образованием одного эндоспора которые могут оставаться жизнеспособными в течение десятилетий и устойчивы к неблагоприятным условиям окружающей среды, таким как засуха, соленость, экстремальный pH, радиация, и растворители. Эндоспоры образуются во время стресса, связанного с питанием, и в результате гидролиза, что позволяет организму существовать в окружающей среде до тех пор, пока условия не станут благоприятными. До процесса споруляции клетки могут стать подвижный производя жгутики, взять ДНК из окружающей среды или произвести антибиотики. Эти ответы рассматриваются как попытки найти питательные вещества путем поиска более благоприятной среды, позволяющей клетке использовать новый полезный генетический материал или просто убивая конкуренцию.[нужна цитата ]
В стрессовых условиях, таких как недостаток питательных веществ, Б. subtilis проходит процесс спороношение. Этот процесс очень хорошо изучен и служит модельным организмом для изучения споруляции.[10]
Хромосомная репликация
Б. subtilis это модельный организм используется для изучения репликации бактериальных хромосом. Тиражирование сингла круговая хромосома инициируется в одном локусе, происхождение (ORIC ). Репликация выполняется двунаправленно, и два вилки репликации продвигаться по хромосоме по часовой стрелке и против часовой стрелки. Репликация хромосомы завершается, когда вилки достигают конечной области, которая расположена напротив начала координат на карта хромосом. Конечная область содержит несколько коротких последовательностей ДНК (Тер сайты), которые способствуют задержке репликации. Определенные белки опосредуют все этапы репликации ДНК. Сравнение белков, участвующих в репликации хромосомной ДНК в Б. subtilis И в кишечная палочка выявляет сходства и различия. Хотя основные компоненты, способствующие инициации, удлинению и прекращению репликации, хорошоконсервированный могут быть обнаружены некоторые важные различия (например, у одной бактерии отсутствуют белки, необходимые для другой). Эти различия подчеркивают разнообразие механизмов и стратегий, которые различные виды бактерий приняли для выполнения дупликации своих геномов.[11]
Геном
Б. subtilis имеет около 4100 генов. Из них только 192 оказались незаменимыми; еще 79 были предсказаны как необходимые. Подавляющее большинство основных генов было отнесено к относительно небольшому количеству областей клеточного метаболизма, при этом около половины участвовало в обработке информации, одна пятая участвовала в синтезе клеточной оболочки и определении формы и деления клетки, а одна десятая была связана с клеткой. энергетика.[12]
Полная последовательность генома Б. subtilis субштамм QB928 имеет 4 146 839 пар оснований ДНК и 4292 гена. Штамм QB928 широко используется в генетических исследованиях из-за наличия различных маркеров [aroI (aroK) 906 purE1 dal (alrA) 1 trpC2].[13]
Несколько некодирующих РНК были охарактеризованы в Б. subtilis генома в 2009 г., в том числе Bsr РНК.[14]Сравнительный геномный анализ на основе микрочипов показал, что Б. subtilis члены демонстрируют значительное геномное разнообразие.[15]
FsrA - это малая РНК нашел в Bacillus subtilis. Это эффектор реакции сохранения железа и подавляет железосодержащие белки в периоды низкой биодоступности железа.[16][17]
Трансформация
Естественная бактериальная трансформация включает передачу ДНК от одной бактерии к другой через окружающую среду. В Б. subtilis длина перенесенной ДНК превышает 1271 килобайт (более 1 миллиона оснований).[18] Перенесенная ДНК, вероятно, представляет собой двухцепочечную ДНК и часто составляет более трети общей длины хромосомы, составляющей 4215 т.п.н.[19] Похоже, что около 7–9% реципиентных клеток занимают целую хромосому.[20]
Чтобы бактерия-реципиент могла связываться, брать экзогенную ДНК другой бактерии того же вида и рекомбинировать ее в свою хромосому, она должна войти в особое физиологическое состояние, называемое компетентность.Компетентность в Б. subtilis индуцируется к концу логарифмического роста, особенно в условиях ограничения аминокислот.[21] В этих стрессовых условиях полубогащения клетки обычно имеют только одну копию своей хромосомы и, вероятно, имеют повышенное повреждение ДНК. Чтобы проверить, является ли преобразование адаптивной функцией для Б. subtilis Чтобы восстановить повреждение ДНК, были проведены эксперименты с использованием ультрафиолетового света в качестве повреждающего агента.[22][23][24] Эти эксперименты привели к выводу, что компетентность с поглощением ДНК специфически индуцируется условиями повреждения ДНК, и что трансформация функционирует как процесс рекомбинационной репарации повреждений ДНК.[25]
В то время как естественное компетентное состояние является обычным явлением в лаборатории Б. subtilis и полевые изоляты, некоторые промышленно значимые штаммы, например Б. subtilis (натто) не склонны к поглощению ДНК из-за наличия систем рестрикционной модификации, которые разрушают экзогенную ДНК. Б. subtilis (натто) мутанты, которые являются дефектными в эндонуклеазе системы рестрикционной модификации типа I, способны действовать как реципиенты конъюгативных плазмид в экспериментах по спариванию, открывая путь для дальнейшей генной инженерии этого конкретного Б. subtilis напряжение.[26]
Использует
1900-е годы

Культуры Б. subtilis были популярны во всем мире до появления антибиотики как иммуностимулирующее средство для лечения желудочно-кишечный и мочеиспускательный канал болезни. Он использовался на протяжении 1950-х годов как Альтернативная медицина, которые при пищеварении значительно стимулируют иммунную активность широкого спектра, включая активацию секреции специфических антитела IgM, IgG и IgA[27] и высвобождение динуклеотидов CpG, индуцирующих активность продуцирования INF A / Y лейкоциты и цитокины важно в развитии цитотоксичность к опухолевые клетки.[28] Он продавался по всей Америке и Европе с 1946 года в качестве иммуностимулирующего средства при лечении заболеваний кишечника и мочевыводящих путей, таких как Ротавирус и Шигеллез. Сообщалось, что в 1966 году армия США сбросила bacillus subtilis на решетку станций метро Нью-Йорка в течение четырех дней, чтобы наблюдать за реакцией людей, покрытых странной пылью, поскольку считается, что из-за своей способности выжить, он все еще присутствует там.[29]
Антибиотик бацитрацин был впервые выделен из группы licheniformis Bacillus subtilis вар Трейси в 1945 году, а бацитрацин до сих пор коммерчески производится путем выращивания бактерий. Bacillus subtilis var Tracy I в емкости с жидкостью среда роста. Со временем бактерии синтезируют бацитрацин и выделяют антибиотик в среду. Затем бацитрацин извлекается из среды с помощью химических процессов.[30]
С 1960-х гг. Б. subtilis имел историю как испытательный вид в экспериментах в космических полетах. Его эндоспоры может прожить до 6 лет в космосе, если покрыт частицами пыли, защищающими его от солнечных ультрафиолетовых лучей.[31] Он использовался как экстремофил показатель выживаемости в космическое пространство Такие как Радиационная ассамблея экзобиологии,[32][33] EXOSTACK,[34][35] и РАЗОБЛАЧАТЬ орбитальные миссии.[36][37][38]
Природные изоляты дикого типа Б. subtilis труднее работать по сравнению с лабораторными штаммами, которые прошли процессы одомашнивания мутагенез и выбор. Эти штаммы часто обладают улучшенными возможностями трансформации (поглощения и интеграции ДНК окружающей среды), роста и потери способностей, необходимых «в дикой природе». И хотя существуют десятки различных штаммов, соответствующих этому описанию, штамм, обозначенный как «168», является наиболее широко используемым. Штамм 168 представляет собой ауксотроф триптофана, выделенный после мутагенеза рентгеновских лучей Б. subtilis Марбургский штамм и он широко используется в исследованиях благодаря своей высокой эффективности трансформации. [39]

B. globigii, тесно связанный, но филогенетически отдельные виды, теперь известные как Bacillus atrophaeus[40][41] использовался в качестве биологической войны имитатор в течение Проект SHAD (он же Проект 112).[42] Последующий геномный анализ показал, что штаммы, использованные в этих исследованиях, были продуктами преднамеренного обогащения штаммов, которые демонстрировали аномально высокие показатели спороношение.[43]
Напряжение Б. subtilis ранее известный как Bacillus natto используется в коммерческом производстве японской еды натто, а также аналогичная корейская кухня Cheonggukjang.
2000-е
- Как модельный организм, Б. subtilis обычно используется в лабораторных исследованиях, направленных на обнаружение основных свойств и характеристик грамположительных спорообразующих бактерий.[15] В частности, основные принципы и механизмы, лежащие в основе формирования прочной эндоспоры, были выведены из исследований образования спор в Б. subtilis.
- Его свойства связывания с поверхностью играют роль в безопасных радионуклидных отходах [например, торий (IV) и плутоний (IV)].[нужна цитата ]
- Благодаря отличным ферментационным свойствам, с высоким выходом продукта (от 20 до 25 грамм на литр) он используется для производства различных ферментов, таких как амилаза и протеазы.[44]
- Б. subtilis используется как почвенный модификатор в садоводство и сельское хозяйство.[45][требуется полная цитата ][46][требуется полная цитата ][47][требуется полная цитата ]
- Это может принести пользу шафран производителей за счет ускорения роста клубнелуковиц и увеличения урожайности биомассы стигмы.[48]
- Он используется как «индикаторный организм» во время процедур газовой стерилизации, чтобы гарантировать успешное завершение цикла стерилизации.[49][требуется полная цитата ][50][требуется полная цитата ] Это связано с трудностями стерилизации эндоспор.
- Б. subtilis было обнаружено, что он действует как полезный фунгицид биопродукта, который предотвращает рост Monilinia Vacinii-corymbosi, он же мумие-ягодный гриб, не влияющий на опыление или качество плодов.[51]
- Как метаболически активный, так и неметаболически активный Б. subtilis было показано, что клетки восстанавливают золото (III) до золота (I) и золота (0), когда присутствует кислород. Это биотическое восстановление играет роль в круговороте золота в геологических системах и потенциально может быть использовано для извлечения твердого золота из указанных систем.
Новые и искусственные грунты
- Новые штаммы Б. subtilis которые могли использовать 4-фтортриптофан (4FTrp), но не канонический триптофан (Trp) для размножения, были изолированы. Поскольку Trp кодируется только одним кодоном, есть свидетельства того, что Trp может быть замещен 4FTrp в генетическом коде. Эксперименты показали, что канонический генетический код может быть изменчивым.[52]
- Рекомбинантные штаммы pBE2C1 и pBE2C1AB использовали для получения полигидроксиалканоаты (PHA), а солодовые отходы могут использоваться в качестве источника углерода для производства более дешевого PHA.[нужна цитата ]
- Он используется для производства гиалуроновая кислота, который используется в секторе совместного ухода в здравоохранении[53][требуется полная цитата ] и косметика.
- Monsanto выделил ген из Б. subtilis который экспрессирует белок B холодового шока и встраивает его в свой засухоустойчивый гибрид кукурузы MON 87460, который был одобрен для продажи в США в ноябре 2011 года.[54][55]
- Был изменен новый штамм для производства медовый[56]
Безопасность
У других животных
Б. subtilis был рассмотрен FDA США Центр ветеринарной медицины и было установлено, что не представляет проблем безопасности при использовании в микробных продуктах прямого кормления, поэтому Ассоциация американских чиновников по контролю кормов внесла его в список одобренных для использования в качестве корма для животных ингредиент согласно Разделу 36.14 «Микроорганизмы прямого кормления».[нужна цитата ]В Канадское агентство по инспекции пищевых продуктов Секция здоровья животных и производственных кормов классифицировала Бациллы обезвоженные культуры, одобренные кормовые ингредиенты в качестве добавки к силосу в соответствии с Приложением IV-Часть 2-Класс 8.6 и которому присвоен международный номер кормового ингредиента IFN 8-19-119.[нужна цитата ]С другой стороны, несколько кормовых добавок, содержащих жизнеспособные споры Б. subtilis положительно оценены Европейское агентство по безопасности пищевых продуктов относительно их безопасного использования для набора веса в животноводстве.
В людях
Б. subtilis споры могут пережить сильную жару во время приготовления. Немного Б. subtilis штаммы ответственны за появление вязкости - липкой, вязкой консистенции, вызванной бактериальным образованием длинноцепочечных полисахариды - в испорченном хлебном тесте. Долгое время туповатость хлеба ассоциировалась однозначно с Б. subtilis виды по биохимическим тестам. Молекулярные анализы (анализ ПЦР с произвольной амплификацией полиморфной ДНК, анализ денатурирующего градиентного гель-электрофореза и секвенирование области V3 рибосомальной ДНК 16S) выявили большее Бациллы разнообразие видов хлеба из панировки, которые, по-видимому, обладают положительной активностью амилазы и высокой термостойкостью.[57] Б. subtilis CU1 (2 × 109 спор в день) оценивали в 16-недельном исследовании (10-дневное введение пробиотика, затем 18-дневный период вымывания каждый месяц; повторение той же процедуры в течение всего 4 месяцев) на здоровых субъектах. Б. subtilis Было обнаружено, что CU1 безопасен и хорошо переносится субъектами без каких-либо побочных эффектов.[58]
Б. subtilis и полученные из него вещества были оценены различными авторитетными органами на предмет их безопасного и полезного использования в пищевых продуктах. В Соединенных Штатах в письме-мнении, опубликованном в начале 1960-х гг. Управление по контролю за продуктами и лекарствами (FDA) признало некоторые вещества, полученные из микроорганизмов, как общепризнанно безопасным (GRAS), включая ферменты карбогидразы и протеазы из Б. subtilis. Мнения основывались на использовании непатогенных и нетоксикогенных штаммов соответствующих организмов и на использовании современных передовых методов производства.[59] FDA заявило, что ферменты, полученные из Б. subtilis до 1 января 1958 г. широко использовались в пищевых продуктах, а нетоксигенные и непатогенные штаммы Б. subtilis широко доступны и безопасно используются в различных пищевых продуктах. Это включает потребление японских ферментированных соевых бобов в виде Натто, который обычно употребляется в Японии и содержит до 108 жизнеспособных клеток на грамм. Ферментированные бобы признаны за их вклад в здоровую микрофлору кишечника и витамин К2 прием; за эту долгую историю широкого использования, натто не был связан с нежелательными явлениями, потенциально связанными с наличием Б. subtilis.[нужна цитата ] Продукт натто и Б. subtilis Натто в качестве его основного компонента - это FOSHU (продукты питания по назначению), одобренные Японией. Министерство здравоохранения, труда и социального обеспечения так же эффективен для сохранения здоровья.[60]
Б. subtilis получил статус «Квалифицированная презумпция безопасности» Европейское агентство по безопасности пищевых продуктов.[61]
Идентификация
Ниже приведена таблица, которая помогает определить Б. subtilis с помощью определенных идентификационных тестов.
| Тест | Результат[62][63] |
|---|---|
| Кислота из глюкозы | +. |
| Кислота из лактозы | - |
| Кислота из сахарозы | + |
| Кислота из маннита | + |
| Каталаза | + |
| Цитрат (Симмонс) | + |
| Эндоспора | + |
| Газ из глюкозы | - |
| Гидролиз желатина | + |
| Окраска по Граму | + |
| Производство сероводорода | - |
| Индол Производство | - |
| Подвижность | + |
| Снижение нитратов | + |
| Оксидаза | + |
| Фенилаланин дезаминаза | - |
| Форма | стержень |
| Гидролиз мочевины | - |
| Voges-Proskaeur-Proskaeur | + |
Смотрите также
Рекомендации
- ^ Euzéby JP (2008). «Бациллы». Список названий прокариот, стоящих в номенклатуре. Получено 2008-11-18.
- ^ Амброзиано Н. (30.06.1999). «Лабораторные биодетекторные тесты должны быть безопасными, общественность должна быть хорошо информированной». пресс-релиз. Национальные лаборатории Лос-Аламоса. Архивировано из оригинал 21 сентября 2008 г.. Получено 2008-11-18.
- ^ Эренберг К.Г. (1835). Physikalische Abhandlungen der Koeniglichen Akademie der Wissenschaften zu Berlin aus den Jahren 1833–1835. С. 145–336.
- ^ Кон Ф (1872). "Untersuchungen über Bacterien". Beiträge zur Biologie der Pflanzen. 1. С. 127–224.
- ^ Yu AC, Loo JF, Yu S, Kong SK, Chan TF (январь 2014 г.). «Мониторинг роста бактерий с помощью настраиваемого резистивного импульсного датчика с использованием поровой техники». Прикладная микробиология и биотехнология. 98 (2): 855–62. Дои:10.1007 / s00253-013-5377-9. PMID 24287933.
- ^ Мэдиган М., Мартинко Дж., Ред. (2005). Биология микроорганизмов Брока (11-е изд.). Прентис Холл. ISBN 978-0-13-144329-7.[страница нужна ]
- ^ Накано М.М., Зубер П. (1998). «Анаэробный рост« строгого аэроба »(Bacillus subtilis)". Ежегодный обзор микробиологии. 52 (1): 165–90. Дои:10.1146 / annurev.micro.52.1.165. PMID 9891797.
- ^ Хонг Х.А., Ханеджа Р., Там Н.М., Каззато А., Тан С., Урдачи М., Бриссон А., Гасбаррини А., Барнс И., Каттинг С.М. (март 2009 г.). "Bacillus subtilis выделен из желудочно-кишечного тракта человека ». Исследования в области микробиологии. 160 (2): 134–43. Дои:10.1016 / j.resmic.2008.11.002. PMID 19068230.
- ^ Судхагар, S; Редди, Рами (апрель 2017 г.). «Влияние подъема на структуру кишечных бактериальных сообществ Apis cerana Fab» (PDF). Журнал энтомологии и зоологии. 5 (3): 2, 7 - через entomoljournal.com.
- ^ Маккенни, Питер Т .; Дрикс, Адам; Эйхенбергер, Патрик (2012). «Эндоспора Bacillus subtilis: сборка и функции многослойной оболочки». Обзоры природы Микробиология. 11 (1): 33–44. Дои:10.1038 / nrmicro2921. PMID 23202530.
- ^ Нуаро П. (2007). "Репликация Bacillus subtilis хромосома ". В Graumann P (ред.). Бациллы: Клеточная и молекулярная биология. Caister Academic Press. ISBN 978-1-904455-12-7.[страница нужна ]
- ^ Кобаяши К., Эрлих С.Д., Альбертини А., Амати Г., Андерсен К.К., Арно М. и др. (Апрель 2003 г.). «Основные гены Bacillus subtilis». Труды Национальной академии наук Соединенных Штатов Америки. 100 (8): 4678–83. Bibcode:2003ПНАС..100.4678К. Дои:10.1073 / pnas.0730515100. JSTOR 3144001. ЧВК 153615. PMID 12682299.
- ^ Ю. С., Йим К. Ю., Цуй С. К., Чан Т.Ф. (ноябрь 2012 г.). «Полная последовательность генома штамма Bacillus subtilis QB928, штамма, широко используемого в генетических исследованиях B. subtilis». Журнал бактериологии. 194 (22): 6308–9. Дои:10.1128 / JB.01533-12. ЧВК 3486399. PMID 23105055.
- ^ Сайто С., Какешита Х, Накамура К. (январь 2009 г.). «Новые гены, кодирующие малую РНК, в межгенных областях Bacillus subtilis». Ген. 428 (1–2): 2–8. Дои:10.1016 / j.gene.2008.09.024. PMID 18948176.
- ^ а б Эрл А.М., Лосик Р., Колтер Р. (июнь 2008 г.). «Экология и геномика Bacillus subtilis». Тенденции в микробиологии. 16 (6): 269–75. Дои:10.1016 / j.tim.2008.03.004. ЧВК 2819312. PMID 18467096.
- ^ Gaballa, A .; Antelmann, H .; Aguilar, C .; Khakh, S.K .; Песня, К. -Б .; Smaldone, G.T .; Хельманн, Дж. Д. (2008). «Железосберегающий ответ Bacillus subtilis опосредуется регулируемой Fur малой РНК и тремя небольшими основными белками». Труды Национальной академии наук. 105 (33): 11927–11932. Дои:10.1073 / pnas.0711752105. ЧВК 2575260. PMID 18697947.
- ^ Smaldone, G.T .; Antelmann, H .; Gaballa, A .; Хельманн, Дж. Д. (2012). «FsrA sRNA и FbpB Protein опосредуют железозависимую индукцию железосеросодержащих оксидаз Bacillus subtilis LutABC». Журнал бактериологии. 194 (10): 2586–2593. Дои:10.1128 / JB.05567-11. ЧВК 3347220. PMID 22427629.
- ^ Сайто Й, Тагучи Х, Акамацу Т (март 2006 г.). «Судьба трансформации бактериального генома после включения в компетентные клетки Bacillus subtilis: непрерывная длина встроенной ДНК». Журнал биологии и биоинженерии. 101 (3): 257–62. Дои:10.1263 / jbb.101.257. PMID 16716928.
- ^ Сайто Й, Тагучи Х, Акамацу Т (апрель 2006 г.). «ДНК, введенная в компетентные клетки Bacillus subtilis путем трансформации лизированного протопласта, является не оцДНК, а дцДНК». Журнал биологии и биоинженерии. 101 (4): 334–39. Дои:10.1263 / jbb.101.334. PMID 16716942.
- ^ Акамацу Т., Тагучи Х (апрель 2001 г.). «Включение всей хромосомной ДНК в лизаты протопластов в компетентные клетки Bacillus subtilis». Биология, биотехнология и биохимия. 65 (4): 823–29. Дои:10.1271 / bbb.65.823. PMID 11388459.
- ^ Анагностопулос С., Спизизен Дж. (Май 1961 г.). «Требования к трансформации у Bacillus Subtilis». Журнал бактериологии. 81 (5): 741–46. Дои:10.1128 / JB.81.5.741-746.1961. ЧВК 279084. PMID 16561900.
- ^ Hoelzer MA, Michod RE (июнь 1991 г.). «Ремонт ДНК и эволюция трансформации в Bacillus subtilis. III. Секс с поврежденной ДНК». Генетика. 128 (2): 215–23. ЧВК 1204460. PMID 1906416.
- ^ Michod RE, Wojciechowski MF, Hoelzer MA (январь 1988 г.). «Ремонт ДНК и эволюция трансформации бактерии Bacillus subtilis». Генетика. 118 (1): 31–39. ЧВК 1203263. PMID 8608929.
- ^ Войцеховски MF, Hoelzer MA, Michod RE (март 1989 г.). «Ремонт ДНК и эволюция трансформации в Bacillus subtilis. II. Роль индуцибельной репарации». Генетика. 121 (3): 411–22. ЧВК 1203629. PMID 2497048.
- ^ Мичод Р.Э., Бернштейн Х., Недельку А.М. (май 2008 г.). «Адаптивное значение секса у микробных патогенов». Инфекция, генетика и эволюция. 8 (3): 267–85. Дои:10.1016 / j.meegid.2008.01.002. PMID 18295550.
- ^ Сато, Мицуру; Томита, Масару; Канеко, Шинья; Ёсикава, Хирофуми; Shiwa, Yuh; Отани, Наото; Симада, Томоэ; Нагасаку, Маюми; Итая, Мицухиро (2019). «Стабильная и эффективная доставка ДНК к Bacillus subtilis (натто) с использованием плазмид конъюгационного переноса pLS20». Письма о микробиологии FEMS. 366 (4). Дои:10.1093 / femsle / fnz032. PMID 30726909.
- ^ Ciprandi G, Scordamaglia A, Venuti D, Caria M, Canonica GW (декабрь 1986). «Влияние Bacillus subtilis на иммунный ответ in vitro». Химиотерапия. 5 (6): 404–07. PMID 3100070.
- ^ Шилаховенко В.А. (июнь 2003 г.). «Противораковые и иммуностимулирующие эффекты нуклеопротеиновой фракции Bacillus subtilis».'". Экспериментальная онкология. 25: 119–23.
- ^ Стивен Розофф, Генри Понтелл, Роберт Тиллман (2020). Прибыль без чести: преступление белых воротничков и разграбление Америки. Пирсон. п. 352-3. ISBN 9780134871486.CS1 maint: несколько имен: список авторов (связь)
- ^ Джонсон Б.А., Анкер Х., Мелени Флорида (октябрь 1945 г.). «Бацитрацин: новый антибиотик, производимый членом группы B. subtilis». Наука. 102 (2650): 376–7. Bibcode:1945Sci ... 102..376J. Дои:10.1126 / science.102.2650.376. PMID 17770204.
- ^ Хорнек Г., Клаус Д.М., Манчинелли Р.Л. (март 2010 г.). «Космическая микробиология». Обзоры микробиологии и молекулярной биологии. 74 (1): 121–56. Bibcode:2010MMBR ... 74..121H. Дои:10.1128 / mmbr.00016-09. ЧВК 2832349. PMID 20197502.
- ^ Доза К, Бигер-Доза А, Диллманн Р., Гилл М., Керц О, Кляйн А., Мейнерт Х., Наврот Т., Ризи С., Стридде С. (1995). "ЭРА-эксперимент" Космическая биохимия."". Успехи в космических исследованиях. 16 (8): 119–29. Bibcode:1995AdSpR..16..119D. Дои:10.1016 / 0273-1177 (95) 00280-Р. PMID 11542696.
- ^ Вайсберг О., Федоров А., Дунюшкин Ф, Кожуховский А., Смирнов В., Аванов Л., Рассел С., Луманн Дж. (1995). «Ионные популяции в хвосте Венеры». Успехи в космических исследованиях. 16 (4): 105–18. Bibcode:1995AdSpR..16..105V. Дои:10.1016/0273-1177(95)00217-3.
- ^ Клэнси, Пол (23 июня 2005 г.). В поисках жизни, в поисках солнечной системы. Издательство Кембриджского университета.[страница нужна ]
- ^ Хорнек Г., Клаус Д.М., Манчинелли Р.Л. (март 2010 г.). «Космическая микробиология». Обзоры микробиологии и молекулярной биологии. 74 (1): 121–56. Bibcode:2010MMBR ... 74..121H. Дои:10.1128 / MMBR.00016-09. ЧВК 2832349. PMID 20197502.
- ^ Фахардо-Кавасос П., Линк Л., Мелош Х. Дж., Николсон В. Л. (декабрь 2005 г.). «Споры Bacillus subtilis на искусственных метеоритах выживают при входе в атмосферу на сверхвысокой скорости: последствия для литопанспермии». Астробиология. 5 (6): 726–36. Bibcode:2005AsBio ... 5..726F. Дои:10.1089 / ast.2005.5.726. PMID 16379527.
- ^ Brandstätter F, Brack A, Baglioni P, Cockell CS, Demets R, Edwards HG и др. (2008). «Минералогическое изменение искусственных метеоритов при входе в атмосферу. Эксперимент STONE-5». Планетарная и космическая наука. 56 (7): 976–84. Bibcode:2008P & SS ... 56..976B. CiteSeerX 10.1.1.549.4307. Дои:10.1016 / j.pss.2007.12.014.
- ^ Wassmann M, Moeller R, Rabbow E, Panitz C, Horneck G, Reitz G, Douki T, Cadet J, Stan-Lotter H, Cockell CS, Rettberg P (май 2012 г.). «Выживание спор устойчивого к УФ-излучению штамма Bacillus subtilis MW01 после воздействия на низкую околоземную орбиту и в смоделированных марсианских условиях: данные космического эксперимента ADAPT на EXPOSE-E». Астробиология. 12 (5): 498–507. Bibcode:2012AsBio..12..498W. Дои:10.1089 / ast.2011.0772. PMID 22680695.
- ^ Zeigler, D. R .; Pragai, Z .; Rodriguez, S .; Chevreux, B .; Глушитель, А .; Альберт, Т .; Bai, R .; Wyss, M .; Перкинс, Дж. Б. (22 августа 2008 г.). «Происхождение 168, W23 и других традиционных штаммов Bacillus subtilis». Журнал бактериологии. 190 (21): 6983–95. Дои:10.1128 / JB.00722-08. ЧВК 2580678. PMID 18723616.
- ^ Накамура Л.К. (1989). "Таксономическая взаимосвязь штаммов Bacillus subtilis с черным пигментом и предложение для Bacillus atrophaeus sp. ноя ". Международный журнал систематической бактериологии. 39 (3): 295–300. Дои:10.1099/00207713-39-3-295.
- ^ Берк С.А., Райт Дж. Д., Робинсон М.К., Бронк Б.В., Уоррен Р.Л. (май 2004 г.). «Обнаружение молекулярного разнообразия в Bacillus atrophaeus с помощью анализа полиморфизма длин амплифицированных фрагментов». Прикладная и экологическая микробиология. 70 (5): 2786–90. Дои:10.1128 / AEM.70.5.2786-2790.2004. ЧВК 404429. PMID 15128533.
- ^ «Проект 112 / ШАД - Корабельная опасность и защита». Департамент США по делам ветеранов. Архивировано из оригинал 21 февраля 2015 г.. Получено 25 февраля 2015.
- ^ Гиббонс Х.С., Брумолл С.М., МакНью Л.А., Далиго Х., Чепмен С., Брюс Д., Каравис М., Креппс М., МакГрегор ПА, Хонг С., Парк К. Х., Акмал А., Фельдман А., Лин Дж. С., Чанг В.Е., Хиггс Б.В., Демирев П. , Линдквист Дж, Лием А., Фохлер Э., Рид Т.Д., Тапиа Р., Джонсон С., Бишоп-Лилли К.А., Деттер С., Хан С., Сожаманнан С., Розенцвейг К.Н., Сковронски Е.В. (март 2011 г.). «Геномные сигнатуры отбора и улучшения штаммов в Bacillus atrophaeus var. Globigii, историческом симуляторе биологической войны». PLOS ONE. 6 (3): e17836. Bibcode:2011PLoSO ... 617836G. Дои:10.1371 / journal.pone.0017836. ЧВК 3064580. PMID 21464989.
- ^ ван Дейл Дж. М., Хеккер М. (январь 2013 г.). «Bacillus subtilis: от почвенной бактерии до фабрики суперсекретирующих клеток». Фабрики микробных клеток. 12 (3): 3. Дои:10.1186/1475-2859-12-3. ЧВК 3564730. PMID 23311580.
- ^ http://www.eppo.int/QUARANTINE/fungi/Monilinia_fructicola/MONIFC_ds.pdf
- ^ Суэйн MR, Рэй RC (2009). «Биоконтроль и другие полезные действия Bacillus subtilis, выделенные из микрофлоры коровьего навоза». Микробиологические исследования. 164 (2): 121–30. Дои:10.1016 / j.micres.2006.10.009. PMID 17320363.
- ^ Янес-Мендисабаль V (2011). «Биологический контроль персиковой коричневой гнили (Monilinia spp.) С помощью Bacillus subtilis CPA-8 основан на производстве фенгицин-подобных липопептидов». Европейский журнал патологии растений. 132 (4): 609–19. Дои:10.1007 / s10658-011-9905-0.
- ^ Sharaf-Eldin M, Elkholy S, Fernández JA, Junge H, Cheetham R, Guardiola J, Weathers P (август 2008 г.). «Bacillus subtilis FZB24 влияет на количество цветков и качество шафрана (Crocus sativus)». Planta Medica. 74 (10): 1316–20. Дои:10.1055 / с-2008-1081293. ЧВК 3947403. PMID 18622904.
- ^ «Международная фармакопея - четвертое приложение: Методы анализа: 5. Фармацевтические технические процедуры: 5.8 Методы стерилизации».
- ^ «Продукты Andersen: биологический индикатор AN-2203 для ЭО (25 шт. В коробке)».
- ^ Нгуги Х.К., Дедей С., Делаплан К.С., Савелле А.Т., Шерм Х. (01.04.2005). «Влияние наносимого на цветы биофунгицида Serenade (Bacillus subtilis) на переменные, связанные с опылением голубики кроличьего глаза». Биологический контроль. 33 (1): 32–38. Дои:10.1016 / j.biocontrol.2005.01.002. ISSN 1049-9644.
- ^ Yu AC, Yim AK, Mat WK, Tong AH, Lok S, Xue H, Tsui SK, Wong JT, Chan TF (март 2014 г.). «Мутации, позволяющие заменить триптофан на 4-фтортриптофан как каноническую аминокислоту генетического кода». Геномная биология и эволюция. 6 (3): 629–41. Дои:10.1093 / gbe / evu044. ЧВК 3971595. PMID 24572018.
- ^ «Архивная копия». Архивировано из оригинал на 2013-08-28. Получено 2013-08-13.CS1 maint: заархивированная копия как заголовок (связь)
- ^ Харриган Г.Г., Ридли В.П., Миллер К.Д., Сорбет Р., Риордан С.Г., Немет М.А., Ривз В., Пестер Т.А. и др. (Октябрь 2009 г.). «Корм и зерно MON 87460, засухоустойчивого гибрида кукурузы, по составу эквивалентны таковым для обычной кукурузы». Журнал сельскохозяйственной и пищевой химии. 57 (20): 9754–63. Дои:10.1021 / jf9021515. PMID 19778059.
- ^ USDA: Определение статуса нерегулируемого для кукурузы MON 87460 (Zea Mays L)
- ^ Блюм, Брайан (17.11.2019). «Израильские студенты получают награду за изготовление меда без пчел». Израиль21c. Получено 2019-11-24.
- ^ Пепе О, Блайотта Дж., Москетти Дж., Греко Т., Виллани Ф. (апрель 2003 г.). «Штаммы-продуценты Bacillus spp. Из пшеничного хлеба и стратегия борьбы с ними с помощью молочнокислых бактерий». Прикладная и экологическая микробиология. 69 (4): 2321–29. Дои:10.1128 / AEM.69.4.2321-2329.2003. ЧВК 154770. PMID 12676716.
- ^ Лефевр, Мари; Racedo, Silvia M .; Denayrolles, Мюриэль; Риперт, Габриель; Дефужер, Томас; Лобач, Александра Р .; Саймон, Райан; Пелерин, Фанни; Юстен, Питер; Урдачи, Мария К. (1 февраля 2017 г.). «Оценка безопасности Bacillus subtilis CU1 для использования в качестве пробиотика для людей». Нормативная токсикология и фармакология. 83: 54–65. Дои:10.1016 / j.yrtph.2016.11.010. PMID 27825987.
- ^ «Неполный список микроорганизмов FDA». Управление по контролю за продуктами и лекарствами. 2002 г.
- ^ Гибсон Г.Р., 2005. Функциональные продукты питания: перспективы пищевых продуктов для конкретных медицинских целей (FOSHU), Колетт Шорт, Yakult UK. Том 1, стр. 7-1.
- ^ Панель EFSA по биологическим опасностям (BIOHAZ) (2010 г.). «Научное заключение о ведении списка микроорганизмов QPS, намеренно добавленных в пищу или корм (обновление 2010 г.)». Журнал EFSA. 8 (12): 1944. Дои:10.2903 / j.efsa.2010.1944.CS1 maint: использует параметр авторов (связь)
- ^ Prokaryotae, Regnum. "Энциклопедия ABIS". www.tgw1916.net. Получено 2016-12-03.
- ^ Leifert C, Epton HA, Sigee DC (30 сентября 1993 г.), Биологический контроль послеуборочных болезней, получено 2016-12-03
внешняя ссылка
- SubtiВики «актуальная информация по всем генам Bacillus subtilis"
- Окончательная оценка риска Bacillus subtilis на EPA.gov. Архивировано 9 сентября 2015 года..
- Bacillus subtilis браузер генома
- Типовой штамм Bacillus subtilis в BacНырять - База метаданных по бактериальному разнообразию