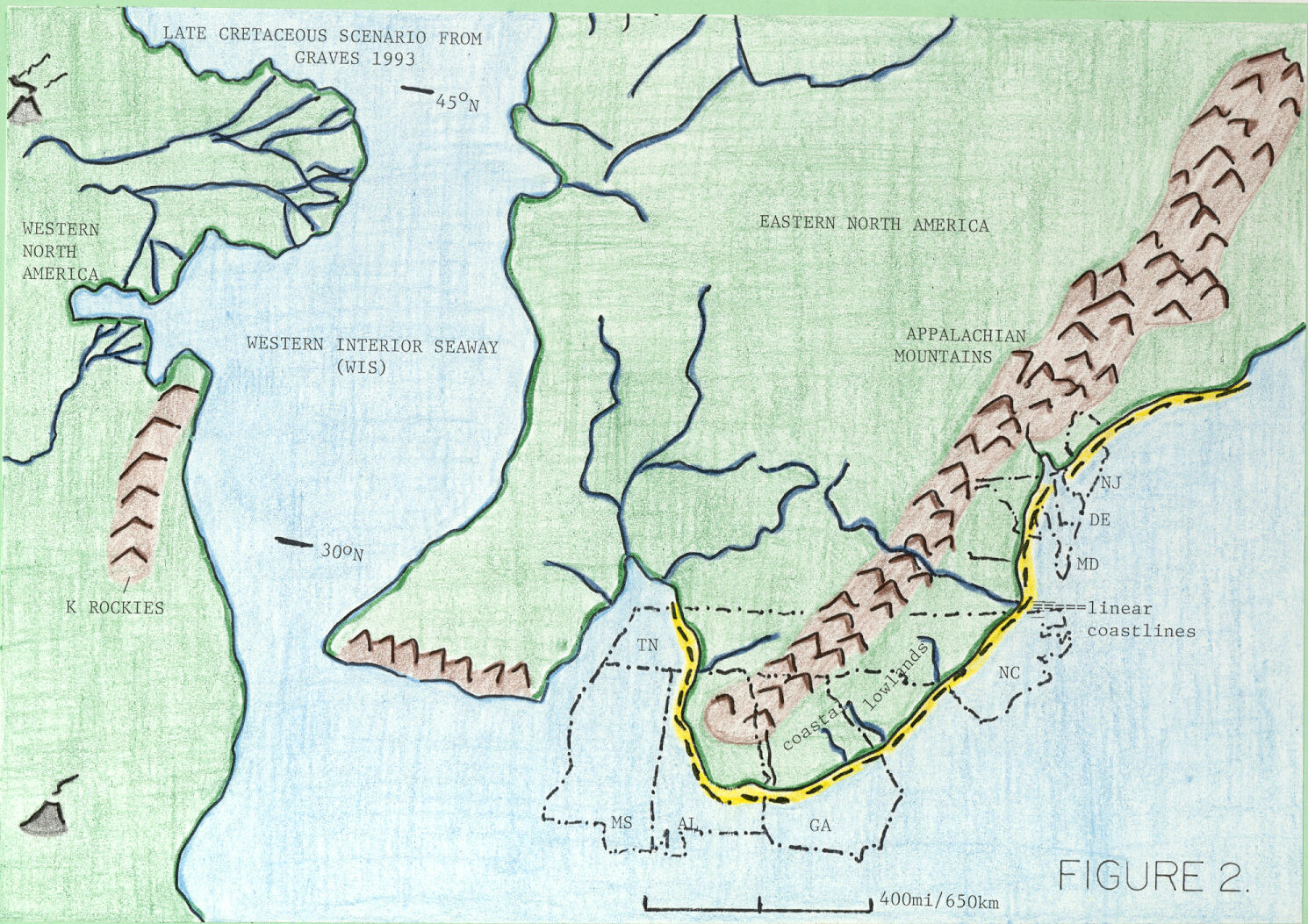
Аппалачи (суша) - Appalachia (landmass)

В течение большей части Поздний мел (100,5–66 миллионов лет назад) восточная половина Северная Америка сформированный Аппалачи (назван в честь Аппалачи ), островной массив суши, отделенный от Ларамидия к западу от Западный внутренний морской путь. Этот морской путь разделил Северную Америку на две огромные суши из-за множества факторов, таких как тектонизм и колебания уровня моря в течение почти 40 миллионов лет.[1][2] Морской путь со временем расширился, разделившись на Дакоты, а к концу мелового периода[3] он отступил к Мексиканский залив и Гудзонов залив. Это привело к тому, что островные массы присоединились к континенту Северной Америки как скалистые горы Роза.[4] От Сеноманский до конца Кампанский возраст Поздний мел, Аппалачи были отделены от остальной части Северной Америки. Как Западный внутренний морской путь отступил в Маастрихтский, Ларамидия и Аппалачи со временем соединились.[5] Из-за этого его фауна был изолирован и развивался совсем иначе, чем тираннозавр, цератопсан, гадрозаврид, пахицефалозавр и анкилозаврид преобладает фауна западной части Северной Америки, известная как "Ларамидия ".[6]
Из-за высокого уровня моря последующая эрозия,[7] и отсутствие орогенный вход отложений в Западный внутренний морской путь, в отличие от восточного побережья Ларамидии, наземных отложений не сохранилось, при этом большая часть останков динозавров произошла от морских туш, которые были перенесены в морскую среду. Такой же недостаток наземных отложений присутствует и на западном побережье Ларамидии. Некоторые отложения в северной части ареала были удалены ледниковая эрозия из Ледяной щит Лаурентиды в течение Четвертичные оледенения,[8] но трудно установить, сколько наносов было удалено и были ли эти отложения более продуктивными, чем те, что остались. Таким образом, относительно мало что известно об Аппалачах по сравнению с Ларамидией, за исключением растений, морских обитателей и насекомых, пойманных в ловушку. янтарь из Нью-Джерси. Кроме того, из-за отсутствия интереса к Аппалачам многие окаменелости, которые были найдены в Аппалачах, не исследованы и остаются в неточных родах, к которым они были отнесены во времена Э. Д. Коуп и О. К. Марш. Лишь несколько окаменелостей земных существ, которые были найдены в этом регионе, дали нам краткое представление о том, какой была жизнь здесь в меловой период. Тем не менее, интерес к этому району возродился благодаря нескольким открытиям, сделанным за последние несколько лет.[9][10][11][12][13] Как упоминалось ранее, об Аппалачах известно не так много, но некоторые ископаемые останки, такие как Формация Вудбайн, Формирование Навесинка,[14] Ископаемые останки Эллисдейла,[15] Мел Мурвилля Формирование, Демополис Мел Формирование, Группа Блэк Крик и Формация Ниобрара,[16] вместе с текущими исследованиями в этой области,[17] дали нам лучший взгляд на этот забытый мир палеонтология.
География
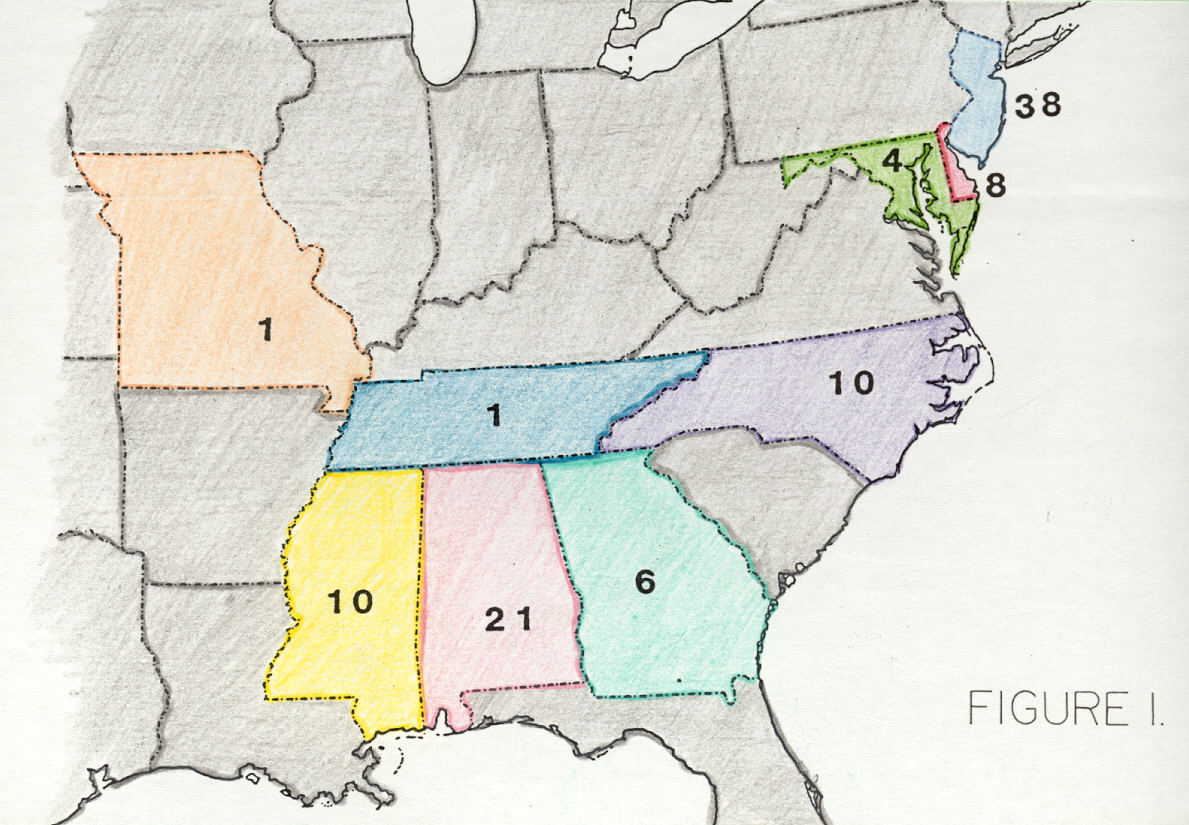
Аппалачи простирались от Квебек и Ньюфаундленд и Лабрадор вплоть до Восточная часть США и на запад к Средний Запад США. Окаменелости, обнаруженные в этих регионах, указывают на то, что территория была покрыта прибрежные равнины и прибрежные низины в меловой период.[18] Некоторые ученые выдвинули идею о том, что архипелаг островов сформировался в то время, когда Западный внутренний морской путь разделял Ларамидию и Аппалачи до ближайшего конца мелового периода. Это позволило бы динозаврам мигрировать на побережье Мексиканского залива и, возможно, объяснить, почему есть некоторые заметные различия с фауной в двух сообществах Аппалачей. Например, в юго-восточном комплексе (который состоит из участков Каролины и побережья Мексиканского залива) есть несколько тираннозавроидов, таких как Аппалачиозавр, некоторые гадрозавроиды, такие как Эотраходон и Lophorhothon, нодозавров, дромеозавров и новых лептоцератопсов, в то время как в северном комплексе (который состоит из Нью-Джерси, Делавэра и Мэриленда) есть несколько тираннозавроидов, таких как Дриптозавр, гадрозавроиды, такие как Гадрозавр, более мелкие тероподы и возможный ламбеозавр в области.[19]
Классифицировать
Окаменелости позвоночных были найдены вдоль Атлантическое побережье а также в других штатах, таких как Алабама, Джорджия, Северная Каролина, Южная Каролина, Миссисипи, Миссури, Кентукки, Теннесси, Канзас, Небраска, Айова и Миннесота. Части Канады, которые были частью Аппалачей в меловой период, включают Манитобу, Онтарио, Квебек, Нунавут, Нью-Брансуик, Ньюфаундленд и Лабрадор, а также Новую Шотландию.[20][21][22]
Фауна
Динозавры
От сеномана до маастрихта Аппалачи населяли различные группы динозавров, в том числе, гадрозавроиды, гадрозавры, нодозавры, лептоцератопсы, неопределенные орнитоподы, тираннозавроиды, дромеозавры, орнитомимиды, и неопределенные манирапторы жил в этом районе. Есть также ископаемые свидетельства возможного хазмозавры, ламбеозавры, зауроподы, аллозавры, ценагнатиды, и троодонтиды которые могли населять этот район.[23][24][6][25][26]
Тираннозавры
В позднемеловой Северной Америке преобладающими хищниками были тираннозавры, огромная хищная тероподы с пропорционально массивными головами, построенными для того, чтобы отрывать плоть от своей добычи. Тираннозавры тоже были доминирующими хищниками в Аппалачах, но, скорее, чем массивные Tyrannosauridae, подобно Горгозавр, Альбертозавр и Lythronax,[27] которые возникли примерно в то же время, когда Западный внутренний морской путь полностью отделил Ларамиду от Аппалачей, меньшего диптозавры были главными хищниками Аппалачей. Вместо того, чтобы развивать огромные головы и массивные тела своих сородичей, у дриптозавров было больше общего с базальными тираннозаврами, такими как Дилонг и Эотираннус, имея длинные руки с тремя пальцами,[28] и были не такими крупными, как самые крупные тираннозавриды, хотя окаменелости из потомакской формации в Нью-Джерси показывают, что некоторые из них действительно развили некоторые из крупнотелых черт, которые можно найти у других тираннозавров.[29] Известно три рода настоящих Аппалачских тираннозавров: Дриптозавр, Аппалачиозавр, и недавно обнаруженный Тейхивенатор в то время как другие неопределенные окаменелости разбросаны по большей части юга Соединенных Штатов, таких как Джорджия, Северная Каролина и Южная Каролина. Ископаемые кости стопы из Аппалачских отложений указывают на еще одного безымянного тираннозавроида размером 9 м (30 футов), что указывает на то, что разнообразие этих базальных тираннозавроидов оставалось высоким в течение последнего мелового периода.[30] Эти окаменелые зубы, возможно, принадлежат к разновидностям Аппалачиозавр или неописанный вид нового тираннозавра.[31][32] Также существует вероятность появления четвертого тираннозавра, известного из Апплахии, известного как Диплотомодон, но это маловероятно, учитывая, что род считается сомнительным. Окаменелости из Нью-Джерси и Делавэра, особенно в Mt. Формирование Лавра и Формация Мерчантвилл соответственно, показали, что примитивные тираннозавроиды были гораздо более разнообразными, чем ожидалось, и некоторые из них выросли до 8-9 метров в длину, что сделало их примерно такими же, как некоторые из более продвинутых тираннозавров, найденных в Ларамидии.[30][33] Окаменелости в сеноманских отложениях также указывают на то, что тираннозавроиды постоянно присутствовали в Аппалачах со среднего мела.[34]
Дромеозавры
Дриптозавры были не единственными хищными динозаврами в Аппалачах. Неопределенный дромеозавр окаменелости, возможно, принадлежащие Velociraptorinae и Saurornitholestinae, и зубы, наиболее близкие к зубам Saurornitholestes,[35] также были обнаружены в Аппалачах; в основном в южных штатах, таких как Миссури, Северная Каролина, Южная Каролина, Алабама, Миссисипи и Джорджия.[36] Находки из формации Campanian Tar Heel в Северной Каролине указывают на то, что здесь могли быть дромеозавриды значительных размеров; промежуточное звено между такими родами, как Saurornitholestes и Дакотараптор. Хотя это открытие известно только по зубам, это открытие указывает на то, что крупные дромеозавриды были частью фауны Аппалачей.[37] Наряду с останками дромеозаврид, в штате Миссури были обнаружены останки тираннозавроидов и возможные останки орнитомимидов.[38] Следует также отметить, что останки дромеозавров чаще встречаются в южном регионе Аппалачей по сравнению с северным регионом, причем их размеры варьируются от более мелких до гигантских форм.[39] Недавние окаменелости, обнаруженные в Нью-Джерси, показывают, что манирапторы присутствовали и в Аппалачах.[40]
Орнитомимиды
Разные орнитомимид кости, такие как Целозавр, также были зарегистрированы в Аппалачах из Миссури, Миссисипи, Алабамы, Джорджии и далеко на севере в таких штатах, как Мэриленд, Нью-Джерси и Делавэр, но теперь считается, что некоторые из них являются костями молодых дриптозавров, а другие принадлежат различным неописанные виды орнитомимид. По состоянию на 2019 год никаких отдельных видов орнитомимозавров еще не идентифицировано, в основном из-за того, что полный скелет еще не обнаружен. Тем не менее, можно предположить, что большинство из них были примерно того же размера, что и их ларамидийские родственники, хотя есть один экземпляр, который мог достигать больших размеров, подобных Галлимим или же Beishanlong.[41][42][38][43]
Другие тероподы
Окаменелости формации Вудбайн, которая является одним из немногих мест окаменелостей, которые являются одними из наиболее хорошо сохранившихся окаменелостей Аппалачей, в Техасе показывают, что другие тероподы могли бродить по Аппалачам примерно в то время, когда впервые сформировалось Западное внутреннее море, они включают возможные образцы аллозавроиды, троодонтиды, ценагнатиды, дромеозавры и тиранозавры.[23][24]
Гадрозавры
Еще одна распространенная группа, возможно, наиболее распространенная разновидность в этом районе,[44] Аппалачских динозавров были Гадрозавроморфа, Hadrosauroida, а Hadrosauridae [45] которая теперь считается их «прародиной»; в конце концов пробравшись в Ларамидию, Азия,[46][47][48][49][50] Европа,[51][52] Южная Америка[53] и Антарктида[54] где они диверсифицировались в ламбеозаврин и зауролофин динозавры, хотя некоторые из примитивных гадрозавров[55] все еще присутствовали до конца мезозоя.[56][57][58][59] В то время как летопись окаменелостей показывает ошеломляющее разнообразие форм гадрозавров в Ларамидии, останки гадрозавров в Аппалачах демонстрируют меньшее разнообразие из-за относительно необычного количества пластов окаменелостей. Однако в Аппалачах известно приличное количество гадрозавров с Протоадрос, Клаозавр, и Lophorhothon представляющий Гадрозавроморфа, Гипсибема Crassicauda и Hypsibema missouriensis представляющий Hadrosauroida, и Гадрозавр и Эотраходон представляющий Hadrosauridae. Эти гадрозавры из Аппалачей, по-видимому, тесно связаны с бесхребетными гадрозаврами Ларамидии, такими как Грипозавр и Эдмонтозавр, несмотря на то, что они не считаются зауролофины. Клаозавр известен по экземпляру, который плавал во Внутреннем морском пути и был обнаружен в Канзасе, возможно, также из Аппалачей, поскольку он был обнаружен ближе к Аппалачской стороне морского пути и неизвестен из западной части Северной Америки. Останки хадрозавра были найдены даже в Айова, хотя и фрагментарно,[60] Теннесси, в первую очередь из Формация Кун-Крик.[61][62] Hypsibema crassicauda,[63] более пятидесяти футов в длину, был одним из крупнейших восточных гадрозавров, переросший некоторых более производных западных гадрозавров, таких как Ламбеозавр и Зауролоф. Род, вероятно, занял экологическую нишу, занятую крупными зауроподы в других областях, возможно, выросли до колоссальных размеров по сравнению с Magnapaulia[64] и Шантунгозавр.[65] Hypsibema missouriensis, был еще одним крупным видом гадрозавров, но он вырос до 45-49 футов, что было не так много, как Hypsibema crassicauda. Когда он был впервые обнаружен в 1945 году, его ошибочно приняли за разновидность зауроподов.[66] У Hypsibema missouriensis, возможно, даже у всех других гадрозавров, живущих в Аппалачах, были зазубренные зубы, которые позволяли жевать растительность в этом районе.[67] Окаменелости гадрозавра из Формация Кангук на острове Аксель-Хейберг в Нунавуте, Канада, показывают, что гадрозавры были довольно широко распространены в Аппалачах. Окаменелости, обнаруженные в формации Кангук, также выявили диетические предпочтения гадрозавров, которые показали, что их диета хвойные породы, стебли, веточки, и различные лиственные растения.[68]
Ламбеозавры
Неопределенный ламбеозаврины остается, в основном похож на Коритозавр, поступили сообщения из Нью-Джерси Формирование Навесинка, Остров Байлот и Новая Шотландия, Канада. Пока невозможно объяснить, как ламбеозаврины могли достичь Аппалачей, хотя некоторые предполагают, что сухопутный мост должен был образоваться где-то во время Кампанский.[69]
Нодозавры
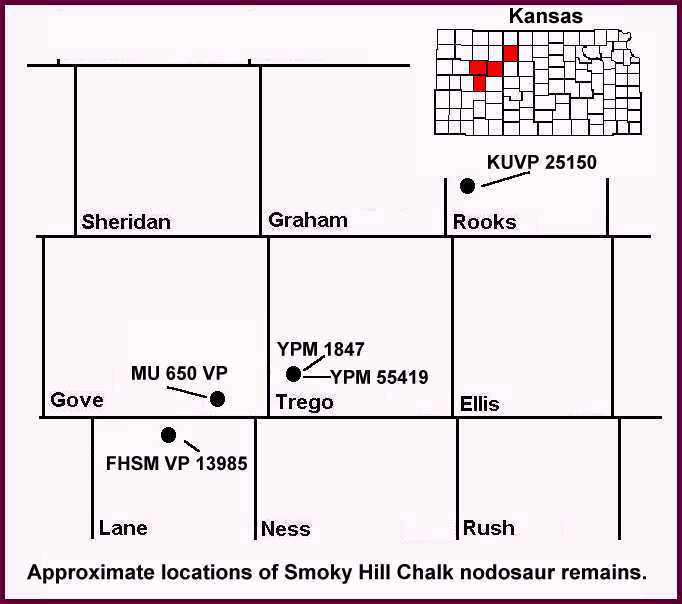
В нодозавриды, группа крупных, травоядных бронированные динозавры напоминающий броненосцы, являются еще одним свидетельством отличия Аппалачии от Ларамидии. В раннем меловом периоде нодозавриды процветали и были одними из самых распространенных динозавров в Северной Америке. Однако к последнему меловому периоду нодозавриды в западной части Северной Америки были редкостью.[70] ограничено формами, такими как Эдмонтония, Денверсавр и Паноплозавр; возможно из-за конкуренции со стороны анкилозавриды; хотя они действительно процветали изолированно, особенно в Аппалачах, как упоминалось ранее и в случае Струтиозавр,[71] Европа тоже. Нодозаврид щитки обычно находили в восточной части Северной Америки, а ископаемые экземпляры очень редки. Часто результаты не являются достаточно диагностическими, чтобы идентифицировать вид, но останки свидетельствуют о большем количестве этих бронированных динозавров в Аппалачах. Несколько экземпляров были обнаружены в Канзас[72] в формации Ниобрара, штат Алабама, в Формация Рипли,[73] Миссисипи, Делавэр, Мэриленд и Нью-Джерси, возможно, принадлежащие множеству различных видов.[74] Пять возможных и наиболее известных примеров нодозаврид Аппалачей, относящихся как к раннему, так и к позднему меловому периоду, включают: Priconodon, Пропаноплозавр, Ниобраразавр,[75][76] Сильвизавр[77] и возможно Иерозавр,[78] хотя его законность оспаривается. Как и Клаозавр образца, возможно, что экземпляры Ниобраразавр, Сильвизавр и Иерозавр поплыли во Внутренний морской путь с востока, так как эти два вида нодозавридов были обнаружены в знаменитом меловые образования[79] Канзаса и не известны ни в одной местности Западной Северной Америки. Канзас также был частью Аппалачей, когда другие части были покрыты океанами, которые были частью Западного внутреннего морского пути.
Лептоцератопсы
В то время как останки продвинутых цератопсов, в первую очередь центрозаврины и хасмозаврины[80] которые были очень распространены в Ларамидии в то время, а не в Аппалачах, лептоцератопсиды каким-то образом удалось заселить это место.[81] А Кампанский -эра лептоцератопсид цератопсан был найден в Образование смоляной пятки, отмечая первое открытие цератопсового динозавра в зоне Аппалачей. У этого экземпляра уникально длинная, тонкая и изогнутая верхняя челюсть, что позволяет предположить, что это было животное со специальной стратегией питания, что является еще одним примером видообразования в окружающей среде острова.[82] Недавно в штате Миссисипи были обнаружены цератопсианские зубы. Формация Сова Крик,[83] возраст которых составляет 67 миллионов лет.[84] Владелец этого единственного зуба, вероятно, был хазмозаврином, поскольку к концу мелового периода центрозаврины полностью исчезли из Северной Америки, хотя они процветали в Азии, как и в случае с Синоцератопс. В то время как останки лептоцератопсидов, те немногие, что были обнаружены в последние годы, были обнаружены в южной части Аппалачей, они, похоже, полностью отсутствуют в северной части Аппалачей, таких как Нью-Джерси, Делавэр и Мэриленд. Предполагает идею, предложенную палеонтологом Дэвидом Р. Швиммером, о возможном провидении во время позднего мелового периода.[85] Не совсем понятно, как лептоцератопсы смогли достичь Аппалачей, однако наиболее распространенной теорией было то, что они остров прыгнул в то время, когда Западный внутренний морской путь разделил североамериканский континент на два разных массива суши таким образом, что некоторые виды лептоцератопсидов, в первую очередь Айкацератопс, смогли добраться до Европы.[86][87] Следует также отметить, что существует явная разница в том, как лептоцератопсы эволюционировали в Аппалачах и Ларамиде. Аппалачский лептоцератопс, обнаруженный в формации Tar Heel, вырос до размеров большой собаки, имел более тонкую челюсть с зубцами, которые загибались вниз и наружу в клюве. Это подразумевает особую стратегию кормления для кормления листвой, которая была родной для Аппалачей во время Кампанский.[88]
Птицы
Несколько останков птиц известны из стоянок Аппалачей, большинство из них морские птицы подобно Гесперорниты, Ихтиорнис, Энантиорниты подобно Халиморнис и Орнитуры подобно Апаторнис и Iaceornis, что, возможно, указывает на то, что в Аппалачах могло быть множество разнообразных птиц, которые были эндемичными для этого региона. Особый интерес возможны литорнитид остается в Нью-Джерси, возможно, одна из лучших записей[89] для меловых птиц[90] поскольку некоторые экземпляры сохранились в зелень[91] в области,[92][93] что представляет собой наглядный пример палеогнат Neornithes в позднем меловом периоде. Однако этот вопрос все еще обсуждается. Примеры птиц, которые были эндемичными для Аппалачей, включают несколько групп, таких как Ржанкообразные, который состоял из Гракулав и Telmatornis, Гусеобразные как представлено Анаталавис, Procellariiformes, с Титтостоникс являясь одним из таких примеров, и Палеотринга и Лаорнис принадлежащий к неизвестной в настоящее время группе птиц.[89] Окаменелости гесперорнитидов также были обнаружены в Арканзасе.[94] Некоторые птицы, обитающие в Канаде, такие как Тингмиаторнис и Canadaga были обнаружены в районах, которые входили в состав Аппалачей.[95][96] Энантиорнитиновые птицы также известны из Аппалачей, как и в случае с Flexomornis из формации Вудбайн из Техаса.[97]
Герпетофауна нединозавров
Амфибии
Сквозь Ископаемые останки Эллисдейла, хорошее изображение нединозаврической фауны Аппалачей присутствует. Среди лиссамфибии, есть доказательства сирениды (в том числе большой Габрозавр ), саламандра батрахозавроидид Parrisia, гилиды, возможные представители Eopelobates и Дискоглосс, демонстрируя близкое сходство с европейскими фаунами, но помимо Габрозавр (который также встречается на Ларамидии) наблюдается высокая степень эндемизма, что предполагает отсутствие взаимообменов с другими массивами суши на протяжении всего позднего мелового периода.[98]
Ящерицы
Существует также высокая степень эндемизма в отношении фауны рептилий: среди чешуекрылых Teiid Прототей - эксклюзив для суши, и местные представители Игуаниды, Helodermatidae, и Некрозавриды.
Черепахи
Среди черепахи, которые довольно часто встречаются в Аппалачах, Adocus, Apalone, и Ботремис хорошо представлены, последние, в частности, более распространены на стоянках Аппалачей, чем Ларамидийские.
Крокодилы
Крокодилы были довольно многочисленны в Аппалачах: девять местных крокодил родов, принадлежащих к нескольким подтвержденным семействам, с вероятностью появления гораздо большего количества неоткрытых крокодилов, ожидающих своего открытия. Goniopholididae представлен Woodbinesuchus,[99] Alligatoridae представлен Боттозавр,[100] Neosuchia представлен Сколомастакс и Deltasuchus,[101] Alligatoroidea представлены Дейнозух и Leidyosuchus, Гавиалоидея представлены Торакозавр,[102] Эоторакозавр,[103] и Крокодилы представлен Бореалозух,[104] также хорошо зарекомендовали себя в Ларамидии, что, вероятно, свидетельствует об их способности пересекать океан. Дейнозух,[105] будучи одним из крупнейших крокодилов в летописи окаменелостей,[106] был хищник вершины что охотились на динозавров[107] в этом районе то же самое относится и к Ларамидии,[108][109] несмотря на то, что большую часть его рациона составляли черепахи[110] и морские черепахи.[111] Однако крокодилы по-прежнему охотились на эндемичных динозавров всякий раз, когда у них была возможность сделать это; есть свидетельства следов укусов крокодила на бедренной кости большого орнитомимозавра, что указывает на хищное поведение местных крокодилов.[112] Окаменелости, обнаруженные в Южной Каролине и Нью-Джерси, показывают, что некоторые из крокодилов, эндемичных для Аппалачей, пережили вымирание динозавров и даже сохранились до кайнозоя.[113][114]
Dyrosauridae
Dyrosauridae, в первую очередь Дирозавр и Гипозавр окаменелости также известны из Аппалачей, особенно в Нью-Джерси, Алабаме и Южной Каролине.[115]
Pholidosauridae
Только один вид Pholidosauridae известно, что жил в Аппалачах; Терминонарис останки которого были обнаружены в Техасе и Канзасе.[116][117]
Птерозавры
Птерозавр окаменелости, в основном похожие на Птеранодон и Никтозавр, раскопаны в Грузии,[118] Алабама[119] и Делавэр.[120] Аналогичным образом аждрахид остается, которые принадлежат Arambourgiania,[121] были обнаружены в Теннесси.[122] Было обнаружено несколько образцов птерозавров, обнаруженных в районах, которые были частью Аппалачей в то время, когда Западный внутренний морской путь разделил Северную Америку на две суши. Окаменелости Аппалачских птерозавров также были обнаружены в Канзас и Техас.[123][124][125][126] Pteranodontidae кажутся наиболее распространенными группами птерозавров в Аппалачах и представлены пятью видами: Птеранодон, Никтозавр, Геостернбергия, Dawndraco, и Аламодактилюс.[127] Члены Орнитохейриды и Anhangueridae представлены Aetodactylus и Цимолиоптер соответственно.[128]
Нет окаменелых останков змеи были обнаружены в Аппалачах в меловой период, но были найдены только в Ларамидии.[129]
Млекопитающие
Несколько видов млекопитающие[130] также присутствуют на Эллисдейл и в обоих Каролины.[131] Наиболее распространены птилодонтоид многотуберкулезные, Такие как Мезодма, Цимолодон и крупный вид. Абсолютное разнообразие видов на суше, а также более раннее появление по сравнению с другими местами позднего мелового периода предполагает, что ptilodontoideans эволюционировали в Аппалачах.[132][133] Метатерии также известны, в том числе альфадонтид,[133] а стагодонтид,[134] и герпетотериид.[135] В отличие от ptilodontoideans, метатерии демонстрируют меньшую степень эндемизма, что подразумевает некоторую степень взаимообмена с Laramidia и Европой. Исследования в этой области показали, чтоTaeniolabidoidea млекопитающие могут проследить свое происхождение здесь и что было несколько видов мультитуберкулезных, эндемичных для Аппалачей.[136] Евтерия окаменелости, в первую очередь коренные зубы, также были обнаружены в Миссисипи. Не исключено, что они принадлежат существу, скорее напоминающему Protungulatum.[137] Род Alphadon а также другие члены Аллотерия останки семьи были раскопаны в Нью-Джерси.[138]
морская жизнь
Хотя до недавнего времени о наземной фауне Аппалачей было мало что известно, морская жизнь в этом районе, а также жизнь, обитавшая в близлежащем Западном внутреннем морском пути, были хорошо изучены палеонтологами в течение многих лет. Такие примеры окаменелостей, которые сохранили остатки морской флоры и фауны того периода, включают формацию Ниобрара, формацию мела Демополис и формацию Мурвилль, - это лишь несколько примеров мест окаменелостей, которые успешно сохранили остатки скелетов различных морских обитателей. существа из мелового периода.[139][140][141][142][143][144][145][146] Примеры морской фауны, обитавшей недалеко от Аппалачей, включают: Chondrichthyes, Osteichthyes, Chelonioidea, Плезиозавры, и Мозазавры, которые в то время были главными хищниками в своей среде обитания.[147][148]
Останки мозазавра были обнаружены даже в штате Миссури.[149]
Окаменелости рыб довольно распространены по всей Аппалачии, особенно в местах, где много мергель, сланец, и известняк.[150][151] Окаменелости рыб, а также большая часть морской фауны меловой эпохи довольно многочисленны в таких регионах, как формация Ниобрара в Канзасе, которая состоит из сланца, песчаника и известняка, а также Формация Вудбери в Нью-Джерси.[152][153]
Членистоногие
Многие виды членистоногих известны с туронского возраста. Нью-Джерси янтарь, расположенный на атлантическом побережье Аппалачей. Членистоногие известны также с сеноманского возраста. Формация Редмонд Лабрадора, Канада.
Флора
В то время как окаменелости в южной части Аппалачей, таких как Алабама и Каролина, содержат очень скудное количество окаменелостей меловых растений, за исключением Джорджии, в северных частях Аппалачей, таких как Нью-Джерси, Мэриленд и Делавэр, есть намного лучше с точки зрения обнаруженных здесь видов растений, особенно с такими ископаемыми местами, как Ископаемые останки Эллисдейла, и дали нам возможность лучше заглянуть в забытый мир. На месте окаменелостей Эллисде раскопки показали, что растения, такие как Picea, Метасеквойя, Лириодендрон, и возможно Ризофора населял регион в поздний меловой период, подразумевая, что окружающей средой в этот период времени был прибрежный лес с несколькими типами морской среды, включая эстуарий, лагунный, морские и наземные. Окаменелости растений, найденные в соседних штатах, таких как Делавэр и Мэриленд, показали, что папоротники, голосеменные, и покрытосеменные действительно обитал в этом районе.[154] Окаменелости покрытосеменных растений были обнаружены в формации Дакота в Небраске.[155] Как упоминалось ранее, в Джорджии имеется богатая летопись окаменелостей растений, восходящих к меловому периоду. Некоторые из примеров флоры, которая присутствовала в этой области, включают Salicaceae, Lauraceae, Секвойные, Moraceae, Pinophyta, Мальпигиалес, Однодольные, Вересковые, Корица, Ранункулалес, Salicaceae, Торрейя, Cupressaceae, Магнолиевые и Рамновые.[156] Окаменелости растений Миннесоты показали, что саговники, вечнозеленые растения, Equisetum, лавры, папоротники, ивы, секвойи, тополя, тюльпановые деревья, и гранаты присутствовали в этом районе во время творения.[157] Также существует огромная концентрация Нормаполь был обнаружен на юго-востоке Соединенных Штатов, что позволяет предположить, что в меловом периоде в этом районе существовала отличная фитогеография.[158]
Окаменелости, обнаруженные недалеко от Формация Редмонд в Лабрадоре показали, что многие современные покрытосеменные впервые появились в Аппалачах.[159] Окаменелости растений найдены в Crawford County, Грузия демонстрирует большое разнообразие растений, таких как спорофиты, гаметофиты, Детрусандра, Hamamelidaceae, Актинидиевые, и в этом регионе было обнаружено 63 вида растений.[160] Покрытосеменные растения были обнаружены в формации Вудбайн.[161] Pinaceae и Lauraceae окаменелости были обнаружены в Миссисипи и Северной Каролине соответственно.[162][163]
Окаменелости растений, найденные в Массачусетсе и Род-Айленде, указывают на то, что климат здесь был субгумидным и паратропическим, что указывает на то, что некоторые из мест обитания Аппалачей в основном состояли из прибрежные равнины и лиственные леса.[164] Растения Pinaceae, Taxodioideae, Араукариевые, Taxaceae, Cycas и Таллофит были найдены в Джорджии и Южной Каролине.[165]
Смотрите также
- Список динозавров Аппалачей - Статья со списком Википедии
Рекомендации
- ^ Нильсен, Карстен Шёдт; Schröder-Adams, Claudia J .; Leckie, Dale A .; Хаггарт, Джеймс У .; Эльбердак, Халифа (декабрь 2008 г.). «Палеоэкологические изменения от турона до сантона в западной части внутреннего моря мелового периода: образования Карлиле и Ниобрара в южной Альберте и юго-западном Саскачеване, Канада». Палеогеография, палеоклиматология, палеоэкология. 270 (1–2): 64–91. Bibcode:2008ППП ... 270 ... 64Н. Дои:10.1016 / j.palaeo.2008.08.018.
- ^ Бертог, Джанет (2010). «Стратиграфия сланцев нижнего Пьера (кампан): последствия для тектонического и эвстатического контроля за распределением фаций». Журнал геологических исследований. 2010: 1–15. Дои:10.1155/2010/910243.
- ^ Эриксон, Дж. Марк (декабрь 1999 г.). "Дакотский перешеек - закрытие западного внутреннего морского пути позднего мела". Слушания Академии наук Северной Дакоты. 53: 124–129. Получено 15 июн 2019.
- ^ Weishampel, David B .; Янг, Лютер (1996). Динозавры восточного побережья. Балтимор, Мэриленд: Издательство Университета Джона Хопкинса.
- ^ Стэнли, Стивен М. (1999). История системы Земля. Нью-Йорк: W.H. Фримен и компания. С. 487–489. ISBN 978-0-7167-2882-5.
- ^ а б Браунштейн, Чейз Д. (февраль 2018 г.). «Биогеография и экология нептичьих динозавров мелового периода Аппалачей». Palaeontologia Electronica. 21 (1.5A): 1–56. Дои:10.26879/801.
- ^ Лю, Лицзюнь (15.06.2014). «Омоложение топографии Аппалачей, вызванное дифференциальной эрозией, вызванной проседанием». Природа Геонауки. 7 (7): 518–523. Bibcode:2014NatGe ... 7..518L. Дои:10.1038 / ngeo2187. ISSN 1752-0894.
- ^ Браун, Дуэйн Д. (сентябрь 1989 г.). «Ледниковая и перигляциальная эрозия Аппалачей». Геоморфология. 2 (1–3): 233–256. Bibcode:1989 Геомо ... 2..233B. Дои:10.1016 / 0169-555X (89) 90014-7.
- ^ Урен, Адам. «Динозавры в Миннесоте: ископаемые когти, найденные в Железном хребте, взволновали ученых». Получено 10 октября 2015.
- ^ Сойер, Лиз. «Ископаемые остатки подтверждают наличие динозавров в Миннесоте». Звездная трибуна. Получено 8 октября 2015.
- ^ «Ископаемые находки за торговым центром Нью-Джерси вызывают ажиотаж». CBS Evening News. Получено 16 ноября 2014.
- ^ Анонимный. "Редкая окаменелость рогатого динозавра, найденная на" затерянном континенте "'". Новости Университета Бата. Университет Бата. Получено 30 ноября 2015.
- ^ Андерсон, Натали. "Eotrachodon orientalis: обнаружены новые виды утконосых динозавров". Science News.com. Новости науки. Получено 26 января 2016.
- ^ Кеннеди, Уильям Дж .; Landman, Neil H .; Коббан, Уильям Обри; Джонсон, Р.О. (13 декабря 2000 г.). «Дополнения к фауне аммонитов верхнемеловой формации Навесинк в Нью-Джерси. Американский музей Novitates». Американский музей Novitates: 31. HDL:2246/2008.
- ^ Галлахер, У. (1997). Когда динозавры бродили по Нью-Джерси. Нью-Брансуик, Нью-Джерси: Издательство Университета Рутгерса.
- ^ "Океаны Канзаса".
- ^ Браунштейн, Чейз (17 января 2018 г.). «Биогеография и экология нептичьих динозавров мелового периода Аппалачей». Palaeontologia Electronica. 21 (1.5a): 1–56. Дои:10.26879/801.
- ^ Браунштейн, Чейз Д. (февраль 2018 г.). «Биогеография и экология нептичьих динозавров мелового периода Аппалачей». Palaeontologia Electronica. 21 (1): 1–56. Дои:10.26879/801.
- ^ Швиммер, Дэвид Р. (1 апреля 2016 г.). «Была ли юго-восточная провинция динозавров в позднем меловом периоде?». Рефераты Геологического общества Америки с программами. 48 (3): 22–3. Дои:10.1130 / абс / 2016SE-271634.
- ^ Кинг-младший, Дэвид Т. «Позднемеловые динозавры юго-востока США». auburn.edu. Обернский университет. Получено 9 января 2019.
- ^ http://webhome.auburn.edu/~kingdat/dino_fig_1.jpg. Отсутствует или пусто
| название =(помощь) - ^ http://webhome.auburn.edu/~kingdat/dino_fig_2.jpg. Отсутствует или пусто
| название =(помощь) - ^ а б Адамс, Томас (13 марта 2017 г.). «Новые данные об экосистемах среднего мела и разнообразии фауны в Аппалачах: выводы из формации Вудбайн (сеноман) в Северном Техасе». Рефераты Геологического общества Америки с программами. 49 (1). Дои:10.1130 / абс / 2017SC-289203.
- ^ а б Драмхеллер, Стефани К. (13 апреля 2018 г.). «Реализация затерянного мира: разнообразие среднего мелового периода в Аппалачах с помощью сведений, полученных с территории Арлингтонского архозавра». Рефераты Геологического общества Америки с программами. 50 (3). Дои:10.1130 / абс / 2018SE-312397.
- ^ Браунштейн, Чейз Д. (ноябрь 2017 г.). "Тираннозавроид плюсны из формации Мерчантвилл в Нью-Джерси увеличивает разнообразие не тираннозавридных тираннозавроидов в Аппалачах". PeerJ. 5 (e4123): e4123. Дои:10.7717 / peerj.4123. ЧВК 5712462. PMID 29204326.
- ^ Кинг, Джеймс Л. (18 октября 2009 г.). «ДИНОЗАВРИАНСКАЯ ФАУНА ЮГО-ВОСТОКА США». Рефераты Геологического общества Америки с программами. 41 (7): 106. Архивировано с оригинал 23 апреля 2020 г.. Получено 16 июн 2019.
- ^ Loewen, Mark A .; Irmis, Randall B .; Setrich, Joseph J. W .; Дж. Карри, Филип; Д. Сэмпсон, Скотт (6 ноября 2013 г.). «Эволюция динозавров-тиранов отслеживает подъем и падение океанов позднего мелового периода». PLOS ONE. 8 (11): 14. Bibcode:2013PLoSO ... 879420L. Дои:10.1371 / journal.pone.0079420. ЧВК 3819173. PMID 24223179.
- ^ Свитек, Брайан (2011). "Удивительные руки дриптозавра". Американский музей Novitates. 3717 (3717): 1–53. Дои:10.1206/3717.2. S2CID 53318615. Получено 11 июля 2011.
- ^ Браунштейн, Чейз Д. (17 апреля 2018 г.). «Тираннозавроид из нижнего сеномана штата Нью-Джерси и его эволюционные и биогеографические последствия». Бюллетень музея естественной истории Пибоди. 59 (1): 95–106. Дои:10.3374/014.058.0210. S2CID 90633156.
- ^ а б Браунштейн, Чейз (декабрь 2018 г.). «БОЛЬШИЕ БАЗАЛЬНЫЕ ТИРАННОЗАВРОИДЫ ИЗ МААСТРИХТАНСКОГО И НАЗЕМНОГО РАЗНООБРАЗНЫХ ПОЗВОНОЧНЫХ В ТЕНИ ИСКЛЮЧЕНИЯ K-PG». Мозазавр. Икс: 105–110. Получено 7 января 2019.
- ^ Чан Гю, Юн (2017). "Teihivenator gen. Nov., Новое родовое название тираннозавроидного динозавра" Laelaps "macropus (Cope, 1868; озабочено Кохом, 1836)". Журнал зоологических и биологических исследований. 4. Архивировано из оригинал на 2017-07-29. Получено 2017-07-23.
- ^ Карр, Томас Д; Уильямсон, Томас Э; Швиммер, Дэвид Р. (2005). «Новый род и вид тираннозавроидов из позднемеловой (средний кампан) формации Демополис в Алабаме». Журнал палеонтологии позвоночных. 25: 119–43. Дои:10.1671 / 0272-4634 (2005) 025 [0119: ANGASO] 2.0.CO; 2.
- ^ Браунштейн, Чейз (30 ноября 2017 г.). "Тираннозавроидная плюсневая кость из формации Мерчантвилл в Делавэре увеличивает разнообразие тиранозавроидов, не являющихся тираннозаврами, в Аппалачах". PeerJ. 5 (e4123): e4123. Дои:10.7717 / peerj.4123. ЧВК 5712462. PMID 29204326.
- ^ Браунштейн, Чейз Доран (2018). «Тираннозавроид из нижнего сеномана штата Нью-Джерси и его эволюционные и биогеографические последствия». Бюллетень музея естественной истории Пибоди. 59: 95–105. Дои:10.3374/014.058.0210. S2CID 90633156.
- ^ Kiernan, Caitlin R .; Швиммери, Дэвид Р. (январь 2004 г.). «Первое обнаружение тероподов-велоцирапторин (Tetanurae, Dromaeosauridae) из восточного побережья Мексиканского залива в США». Мозазавр. 7: 89–93. Получено 7 мая 2004.
- ^ Западный Край, Аундреа. «Дромеозавры». Энциклопедия Алабамы. Энциклопедия Алабамы. Получено 18 мая 2016.
- ^ Браунштейн, Чейз (2018). «Гигантский дромеозаврид из Северной Каролины». Меловые исследования. 92: 1–7. Дои:10.1016 / j.cretres.2018.07.006.
- ^ а б Исправить, Майкл Ф. «Динозаврия и связанная с ней фауна позвоночных в районе хронистов позднего мела на юго-востоке Миссури». Геологическое общество Америки. Получено 1 апреля 2004.
- ^ Браунштейн, Чейз Д. (5 апреля 2018 г.). «Разнообразие динозавров-хищников на юго-востоке Северной Америки выявлено в первой достоверной записи из Северной Каролины». Препринты PeerJ: 1–19. Дои:10.7287 / peerj.preprints.26829v1. Получено 14 июн 2019.
- ^ Браунштейн, Чейз Доран (1 ноября 2019 г.). «Новые сведения о тероподах из последнего мела Нью-Джерси и фауны Маастрихтских Аппалачей». Королевское общество открытой науки. 6 (11): 191206. Bibcode:2019RSOS .... 691206B. Дои:10.1098 / rsos.191206. ЧВК 6894593. PMID 31827856.
- ^ Baird, D .; Хорнер, Дж. (1979). «Меловые динозавры Северной Каролины». Бримлеяна. 2: 1–28.
- ^ Браунштейн, К. (24.07.2017). «Образцы теропод из формации Навесинк и их значение для разнообразия и биогеографии орнитомимозавров и тираннозавроидов в Аппалачах». Препринты PeerJ. 5: e3105v1. Дои:10.7287 / peerj.preprints.3105v1.
- ^ Браунштейн, Чейз Доран (24 июля 2017 г.). «Образцы теропод из формации Навесинк и их значение для разнообразия и биогеографии орнитомимозавров и тираннозавроидов в Аппалачах». Препринты PeerJ. e3105v1. Получено 23 декабря 2019.
- ^ Кинг-младший, Дэвид Т. «Позднемеловые динозавры на юго-востоке США». aubrun.edu. Auburn University Press. Получено 14 сентября 2016.
- ^ Колберт, Эдвин Х. (1948). «Динозавр Hadrosaurian из Нью-Джерси». Труды Академии естественных наук Филадельфии. 100: 23–37. JSTOR 4064414.
- ^ Godefroit, P .; Болотский, Ю.Л .; Лаутерс, П. (2012). Джогер, Ульрих (ред.). "Новый динозавр-зауролофин из позднего мела Дальнего Востока России". PLOS ONE. 7 (5): e36849. Bibcode:2012PLoSO ... 736849G. Дои:10.1371 / journal.pone.0036849. ЧВК 3364265. PMID 22666331.
- ^ Годфруа, Паскаль; Болотский, Юрий; Алифанов, Владимир (2003). «Замечательный гадрозавр с полым гребнем из России: азиатское происхождение ламбеозавров». Comptes Rendus Palevol. 2 (2): 143–151. Дои:10.1016 / S1631-0683 (03) 00017-4.
- ^ Годфруа, Паскаль; Шуцин Зан; Лиюн Цзинь (2000). "Charonosaurus jiayinensis п. г., п. sp., динозавр-ламбеозавр из позднего маастрихта на северо-востоке Китая ". Comptes Rendus de l'Académie des Sciences, Série IIA. 330 (12): 875–882. Bibcode:2000CRASE. 330..875 г. Дои:10.1016 / S1251-8050 (00) 00214-7.
- ^ Болотский, Ю. & Курзанов, С. 1991. [Гадрозавры Амурской области.] В: [Геология границы Тихого океана]. Благовещенск: Амурский КНИИ. 94-103. [На русском]
- ^ Godefroit, P .; Болотский, Ю.Л .; Ван Иттербек, Дж. (2004). «Динозавр-ламбеозавр Amurosaurus riabinini из маастрихта Дальнего Востока России». Acta Palaeontologica Polonica. 49 (4): 585–618.
- ^ Казановас, М.Л; Переда-Субербиола, X .; Santafé, J.V .; Weishampel, D.B. (1999). «Первый гадрозаврид ламбеозавров из Европы: палеобиогеографические последствия». Геологический журнал. 136 (2): 205–211. Bibcode:1999ГеоМ..136..205С. Дои:10,1017 / с0016756899002319.
- ^ Переда-Субербиола, Ксабье; Хосе Игнасио Канудо; Пенелопа Крусадо-Кабальеро; Хосе Луис Барко; Ньевес Лопес-Мартинес; Ориол Омс; Хосе Игнасио Руис-Оменьяка (2009). «Последние динозавры-гадрозавриды в Европе: новый ламбеозаврин из самых верхних меловых периодов Арена (Уэска, Испания)» (PDF). Comptes Rendus Palevol. 8 (6): 559–572. Дои:10.1016 / j.crpv.2009.05.002.
- ^ Рубен Д. Хуарес Вальери; Хосе А. Аро; Лукас Э. Фиорелли; Хорхе О. Кальво (2010). «Новый гадрозавроид (Dinosauria: Ornithopoda) из формации Аллен (поздний мел) в Патагонии, Аргентина» (PDF). Revista del Museo Argentino de Ciencias Naturales N.s. 11 (2): 217–231. Архивировано из оригинал (PDF) на 2011-09-03. Получено 2016-09-13.
- ^ Дело, Джадд А .; Мартин, Джеймс Э .; Чейни, Дэн С .; Регуреро, Марсело; Marenssi, Sergio A .; Сантильяна, Серджио М .; Вудберн, Майкл О. (25 сентября 2000 г.). «Первый динозавр с утконосом (семейство Hadrosauridae) из Антарктиды». Журнал палеонтологии позвоночных. 20 (3): 612–614. Дои:10.1671 / 0272-4634 (2000) 020 [0612: tfdbdf] 2.0.co; 2. JSTOR 4524132.
- ^ Далла Веккья, Ф. М. (2009). "Tethyshadros insularis, новый гадрозавроидный динозавр (Ornithischia) из верхнего мела Италии ". Журнал палеонтологии позвоночных. 29 (4): 1100–1116. Дои:10.1671/039.029.0428.
- ^ Лунд, Эрик К .; Гейтс, Терри А. (январь 2006 г.). «Историко-биогеографическое исследование динозавров гадрозавров». Цитировать журнал требует
| журнал =(помощь) - ^ Кэй, Джон М .; Рассел, Дейл. (1973). «Самая старая запись гадрозавров о динозаврах в Северной Америке». Журнал палеонтологии: 91–93.
- ^ «Исследовательская группа определила редкого динозавра из Аппалачей». 21 января 2016 г.. Получено 11 сентября 2016. Цитировать журнал требует
| журнал =(помощь) - ^ Затишье, Ричард С .; Райт, Нельда Э. (1942). «Гадрозавровые динозавры Северной Америки». Специальные статьи Геологического общества Америки. 40. Геологическое общество Америки. С. 1–272. Дои:10.1130 / SPE40-p1. ISBN 9780813720401.
- ^ Витцке, Брейн Дж. (12 августа 2014 г.). «Динозавры в Айове». Геологическое общество Айовы. Департамент природных ресурсов штата Айова, Университет Айовы. Получено 12 августа 2014.
- ^ Байран, Джонатан Р .; Frederick, Daniel L .; Швиммер, Дэвид Р .; Сиссер, Уильям Г. (июль 1991 г.). «Первая запись о динозавре из Теннесси: кампанский гадрозавр». Журнал палеонтологии. 65 (4): 696–697. Дои:10.1017 / S0022336000030808. Получено 11 сентября 2016.
- ^ Маркин, Вальтер Л .; Гибсон, Майкл А. (3 ноября 2010 г.). «Открытие второго хадрозавра из позднемеловой формации Coon Creek, Западный Теннесси». Рефераты Геологического общества Америки с программами. 42 (5): 631.
- ^ Коуп, Э. (1869). "Замечания по Eschrichtius polyporus, Hypsibema crassicauda, Hadrosaurus tripos, и Полидектес битургидус". Труды Академии естественных наук Филадельфии. 21: 191–192.
- ^ Альбер, Прието-Маркес; Луис, М. Чиаппе; Шантану, Х. Джоши (12 июня 2012 г.). "Динозавр-ламбеозавр Magnapaulia laticaudus из позднего мела Нижней Калифорнии, Северо-Западная Мексика". PLOS ONE. 7 (6): 29. Bibcode:2012PLoSO ... 738207P. Дои:10.1371 / journal.pone.0038207. ЧВК 3373519. PMID 22719869.
- ^ Чейз (22.06.2015). "Допотопные звери Востока: Hypsibema crassicauda". thetetanuraeguy.wordpress.com. Получено 22 июн 2015.
- ^ Гилмор, Чарльз У .; Стюарт, Дэн Р. (январь 1945 г.). «Новый динозавр зауроподов из верхнего мела штата Миссури». Журнал палеонтологии. 19 (1): 23–29. JSTOR 1299165.
- ^ "Добро пожаловать в лабораторию подготовки ископаемых!". Музей естественной истории округа Боллинджер. Архивировано из оригинал на 2016-06-23. Получено 2016-09-13.
- ^ ВАВРЕК, МЭТЬЮ Дж .; HILLS, LEN V .; КЮРРИ, ФИЛИПП Дж. (2014). «Гадрозаврид (Dinosauria: Ornithischia) из позднемеловой (кампанской) формации Кангук острова Аксель-Хейберг, Нунавут, Канада, и его экологические и географические последствия». Арктический. 67 (1): 1–9. Дои:10.14430 / арктика4362. JSTOR 24363716. S2CID 55810951.
- ^ Чейз (30.07.2015). "Ответ на подкаст № 45 The Tetrapod Zoology: Почему ламбеозаврины действительно сохранились в Маастрихте". Одиссея времени. Анонимный. Получено 30 июля 2015.
- ^ Баккер, Роберт Т. (1988). «Обзор нодозавроидов позднего мела: Denversaurus schlessmani, новый бронированный динозавр из позднего мела в Южной Дакоте, последний выживший из нодозавров, с комментариями об отношениях стегозавров и нодозавров». Hunteria. 1 (3): 1–23.
- ^ Garcia, G .; Переда-Субербиола, X. (2003). "Новый вид Струтиозавр (Dinosauria: Ankylosauria) из верхнего мела Виллеверака (юг Франции) ". Журнал палеонтологии позвоночных. 23 (1): 156–165. Дои:10.1671 / 0272-4634 (2003) 23 [156: ansosd] 2.0.co; 2.
- ^ «Примерное местонахождение останков нодозавра Смоки Хилл Мел». Получено 13 сентября 2016.
- ^ Брунс, Майкл Э. «Новый Аппалачский бронированный материал динозавров (Nodosauridae, Ankylosauria) из маастрихтской формации Рипли в Алабаме». Геологическое общество Америки. Геологическое общество Америки. Получено 1 апреля 2016.
- ^ Эберсол, июн. «Нодозавр». Энциклопедия Алабамы. Энциклопедия Алабамы. Получено 18 мая 2016.
- ^ Эверхарт, Майкл Дж .; Хамм, Шон А. (январь 2005 г.). «Новый образец нодозавра (Dinosauria: Nodosauridae) из мела Смоки-Хилл (верхний мел) в западном Канзасе». Труды Канзасской академии наук. 108 (1&2): 15–21. Дои:10.1660 / 0022-8443 (2005) 108 [0015: ANNSDN] 2.0.CO; 2.
- ^ Карпентер, Кеннет; Эверхарт, Майкл Дж. (Апрель 2007 г.). "Skull of the ankylosaur Niobrarasaurus coleu (Ankylosauria: Nodosauridae) from the Smoky Hill Chalk (Coniacian) of western Kansas". Труды Канзасской академии наук. 110 (1 & 2): 1–9. Дои:10.1660/0022-8443(2007)110[1:SOTANC]2.0.CO;2.
- ^ Eaton, T.H.; Jr (1960). "A new armored dinosaur from the Cretaceous of Kansas". The University of Kansas Paleontological Contributions: Vertebrata. 8: 1–24. HDL:1808/3799.
- ^ Wieland, G. R (1909). "A new armored saurian from the Niobrara". Американский журнал науки. 27 (159): 250–2. Bibcode:1909AmJS...27..250W. Дои:10.2475/ajs.s4-27.159.250.
- ^ Carpenter, Kenneth; Dilkes, David; Weishampel, Dave (June 1995). "The Dinosaurs of the Niobrara Chalk Formation (Upper Cretaceous, Kansas)". Журнал палеонтологии позвоночных. 15 (2): 275–297. Дои:10.1080/02724634.1995.10011230.
- ^ "Amazing horned dinosaurs unearthed on 'lost continent'; New discoveries include bizarre beast with 15 horns". ScienceDaily. Университет Юты. Получено 22 сентября 2010.
- ^ Anonymous (2015-09-14). "A new Leptoceratopsid Ceratopsian From Campanian Cretaceous Appalachia". The Dragon's Tales. Получено 14 сентября 2015.
- ^ Longrich, Nicholas R. (2016). "A ceratopsian dinosaur from the Late Cretaceous of eastern North America, and implications for dinosaur biogeography". Меловые исследования. 57: 199–207. Дои:10.1016/j.cretres.2015.08.004.
- ^ Brantley, Mary Grace. "Paleontologists make big dinosaur discovery in Mississippi". MSNewsNow. Архивировано из оригинал 31 августа 2016 г.. Получено 14 сентября 2016.
- ^ Fleet, Micah (2016-07-29). "Rare dinosaur tooth found in Mississippi". wapt.com. 16 Новости WAPT. Получено 14 сентября 2016.
- ^ David R., Schwimmer. "Was There a Southeastern Dinosaur Province in the Late Cretaceous?". 1 апреля 2016 г.. Геологическое общество Америки. Получено 12 сентября 2016.
- ^ Osi, Attila; Батлер, Ричард Дж .; Weishampel, David B. (27 May 2010). «Поздний меловой динозавр цератопса из Европы с азиатским родством». Природа. 465 (7297): 466–468. Bibcode:2010Натура.465..466O. Дои:10.1038 / природа09019. PMID 20505726. S2CID 205220451.
- ^ LINDGREN, JOHAN; CURRIE, PHILIP J.; SIVERSON, MIKAEL; REESS, JAN; LINDGREN, FILIP (2007). "The First Neoceratopsian Dinosaur Remains from Europe". Палеонтология. 50 (4): 929–937. Дои:10.1111/j.1475-4983.2007.00690.x.
- ^ Longrich, Nicholas (January 2016). "A ceratopsian dinosaur from the Late Cretaceous of eastern North America, and implications for dinosaur biogeography". Меловые исследования. 57: 199–207. Дои:10.1016/j.cretres.2015.08.004.
- ^ а б Olson, Storrs L .; Parris, David C. (1987). "The Cretaceous Birds of New Jersey" (PDF). Вклад Смитсоновского института в палеобиологию. 63 (63): 1–25. Дои:10.5479/si.00810266.63.1.
- ^ Wetmore, Alexander (April 1930). "The Age of the Supposed Cretaceous Birds from New Jersey". Аук. 47 (2): 186–188. Дои:10.2307/4075921. JSTOR 4075921. S2CID 87807904.
- ^ Baird, Donald (April 1967). "Age of Fossil Birds from the Greensands of New Jersey". Аук. 84 (2): 260–262. Дои:10.2307/4083191. JSTOR 4083191.
- ^ Palaeogene Fossil Birds
- ^ A lithornithid (Aves: Palaeognathae) from the Paleocene (Tiffanian) of southern California
- ^ Bell, Alyssa; Irwin, Kelly J.; Davis, Leo Carson (September 2015). "Hesperornithiform Birds from the Late Cretaceous (Campanian) of Arkansas, USA". Труды Канзасской академии наук. 118 (3 & 4): 219–229. Дои:10.1660/062.118.0305. S2CID 83921936. Получено 13 июн 2019.
- ^ Bono, R.K.; Clarke, J .; Tarduno, J.A.; Brinkman, Donald (2016). "A Large Ornithurine Bird (Tingmiatornis arctica) from the Turonian High Arctic: Climatic and Evolutionary Implications". Научные отчеты. 6: 38876. Bibcode:2016NatSR...638876B. Дои:10.1038/srep38876. ЧВК 5171645. PMID 27991515.
- ^ Hou (1999). "New hesperornithid (Aves) from the Canadian Arctic". Позвоночные PalAsiatica. 37 (7): 228–233.
- ^ TYKOSKI, RONALD S.; FIORILLO, ANTHONY R. (January 2010). "AN ENANTIORNITHINE BIRD FROM THE LOWER MIDDLE CENOMANIAN OF TEXAS". 30. 1: 288–292. Получено 16 ноября 2020.
- ^ Le Loeuff, J (1991). "The Campano-Maastrichtian vertebrate faunas of southern Europe and their relationship with other faunas in the world; paleobiogeographic implications". Cretaceous Res. 12 (2): 93–114. Дои:10.1016/s0195-6671(05)80019-9.
- ^ Lee, Yuong–Nam (1997). "The Archosauria from the Woodbine Formation (Cenomanian) in Texas". Журнал палеонтологии. 71 (6): 1147–1156. Дои:10.1017/S0022336000036088.
- ^ Cossette, Adam P.; Brochu, Christopher A. (2018). "A new specimen of the alligatoroid Bottosaurus harlani and the early history of character evolution in alligatorids". Журнал палеонтологии позвоночных. 38 (4): (1)-(22). Дои:10.1080/02724634.2018.1486321. S2CID 92801257.
- ^ Adams, T.L.; Noto, C.R.; Drumheller, S. (2017). "A large neosuchian crocodyliform from the Upper Cretaceous (Cenomanian) Woodbine Formation of North Texas". Журнал палеонтологии позвоночных. 37 (4): e1349776. Дои:10.1080/02724634.2017.1349776. S2CID 133647239.
- ^ Troxell, Edward L. (September 1925). "Thoracosaurus, A Cretaceous Crocodile". Американский журнал науки. 5 (10): 219–233. Bibcode:1925AmJS...10..219T. Дои:10.2475/ajs.s5-10.57.219.
- ^ Brochu, Christopher A. (5 January 2004). "A new Late Cretaceous gavialoid crocodylian from eastern North America and the phylogenetic relationships of thoracosaurs". Журнал палеонтологии позвоночных. 24 (3): 610–633. Дои:10.1671/0272-4634(2004)024[0610:ANLCGC]2.0.CO;2.
- ^ Brochu, Christopher A .; Parris, David C.; Grandstaff, Barbara Smith; Denton Jr., Robert K.; Gallagher, William B. (12 January 2012). "A new species of Borealosuchus (Crocodyliformes, Eusuchia) from the Late Cretaceous–early Paleogene of New Jersey". Журнал палеонтологии позвоночных. 32 (1): 105–116. Дои:10.1080/02724634.2012.633585. S2CID 83931184.
- ^ Schwimmer, David R. (12 June 2002). King of the Crocodylians: The Paleobiology of Deinosuchus. Bloomington, IN: Indiana University Press. п. 240.
- ^ Erickson, Gregory M.; Brochu, Christopher A. (18 June 1999). "How the 'terror crocodile' grew so big". Природа. 398 (6724): 205–206. Bibcode:1999Natur.398..205E. Дои:10.1038/18343. S2CID 4402210.
- ^ Handwerk, Brain (2010-03-25). "Feces, Bite Marks Flesh Out Giant Dino-Eating Crocs". National Geographic News. Получено 25 марта 2010.
- ^ RIVERA-SYLVA, Héctor E.; FREY, Eberhard; GUZMÁN-GUTIÉRREZ, José Rubén (2009). "Evidence of predation on the vertebra of a hadrosaurid dinosaur from the Upper Cretaceous (Campanian) of Coahuila, Mexico". Carnets de Géologie: 1–7.
- ^ Rivera-Sylva, Hector E.; МЫ. Hone, David; Dodson, Peter (2012). "Bite marks of a large theropod on an hadrosaur limb bone from Coahuila, Mexico" (PDF). 64 (1): 157–161. Цитировать журнал требует
| журнал =(помощь) - ^ Milan, J; Лукас, Спенсер Дж .; Lockley, M G; Schwimmer, David R. (January 2010). "Bite Marks of the Giant Crocodylian Deinosuchus on Late Cretaceous (Campanian) Bones". New Mexico Museum of Natural History and Science Bulletin. 51: 183–190.
- ^ Harrell, Samantha D.; Schwimmer, David R. (2010). "Coprolites of Deinosuchus and other crocodylians from the Upper Cretaceous of western Georgia, USA": 1–7. Цитировать журнал требует
| журнал =(помощь) - ^ Brownstein, Chase D. (11 June 2018). "Trace fossils on dinosaur bones reveal ecosystem dynamics along the coast of eastern North America during the latest Cretaceous". PeerJ. 6: 23. Дои:10.7717/peerj.4973. ЧВК 6001717. PMID 29910985.
- ^ Erickson, Bruce (1998). "Crocodilians of the Black Mingo Group (Paleocene) of the South Carolina Coastal Plain". Труды Американского философского общества. 88 (4): 196–214. Дои:10.2307/1006674. JSTOR 1006674.
- ^ Cossette, Adam; Christopher, Christopher (3 April 2019). "A new specimen of the alligatoroid Bottosaurus harlani and the early history of character evolution in alligatorids". Журнал палеонтологии позвоночных. 38 (4): (1)-(22). Дои:10.1080/02724634.2018.1486321. S2CID 92801257.
- ^ Callahan, Wayne; Pellegrini, Rodrigo; Schein, Jason; Parris, David C. (January 2015). "A Nearly Complete Specimen of Hyposaurus Rogersii (Crocodylomorpha, Dyrosauridae) from the Late Cretaceous-Early Paleogene of New Jersey". Общество палеонтологии позвоночных. Дои:10.13140/RG.2.1.2253.2724. Получено 4 января 2019. Цитировать журнал требует
| журнал =(помощь) - ^ Adams, Thomas L .; Polcyn, Майкл Дж .; Матеус, Октавио; Winkler, Dale A .; Jacobs, Louis L. (May 2011). "First occurrence of the long-snouted crocodyliform Terminonaris (Pholidosauridae) from the Woodbine Formation (Cenomanian) of Texas". Журнал палеонтологии позвоночных. 31 (3): 712–716. Дои:10.1080/02724634.2011.572938. S2CID 86061260.
- ^ Шимада, Кеншу; Parris, David C. (Spring 2007). "A Long-Snouted Late Cretaceous Crocodyliform, Terminonaris cf. T. browni, from the Carlile Shale (Turonian) of Kansas". Труды Канзасской академии наук. 110 (1/2): 107–115. Дои:10.1660/0022-8443(2007)110[107:ALLCCT]2.0.CO;2. JSTOR 20476300.
- ^ Schwimmer, David R.; Падиан, Кевин; Woodhead, Alfred B. (1985). "First Pterosaur Records from Georgia: Open Marine Facies, Eutaw Formation (Santonian)". Журнал палеонтологии. 59 (3): 674–676. JSTOR 1304987.
- ^ Westfall, Aundrea. "Pterosaurs". Encyclopedia of Alabama. Получено 26 августа 2016.
- ^ Bairid, Donald; Galton, Peter M. (1981). "Pterosaur Bones from the Upper Cretaceous of Delaware". Журнал палеонтологии позвоночных. 1 (1): 67–71. Дои:10.1080/02724634.1981.10011880. JSTOR 4522837.
- ^ Harrell Jr., T. Lynn; Gibson, Michael A.; Langston Jr., Wann (1 December 2016). "A cervical vertebra of Arambourgiania philadelphiae (Pterosauria, Azhdarchidae) from the late Campanian micaceous facies of the Coon Creek Formation in McNairy County, Tennessee, USA". Бык. Alabama Mus. Nat. Hist. 33: 94–103.
- ^ Gibson, Michael A. "Review of Vertebrate Diversity n the Coon Creek Formation Lagerstätte (Late Cretaceous) of Western Tennessee". Геологическое общество Америки. Получено 31 марта 2008.
- ^ S. Bennett, Christopher (February 1992). "Sexual dimorphism of Pteranodon and other pterosaurs, with comments on cranial crests". Журнал палеонтологии позвоночных. 12 (4): 422–434. Дои:10.1080/02724634.1992.10011472.
- ^ Carpenter, Kenneth (2008). "Vertebrate Biostratigraphy of the Smoky Hill Chalk (Niobrara Formation) and the Sharon Springs Member (Pierre Shale)". High-Resolution Approaches in Stratigraphic Paleontology. Разделы геобиологии. 21. pp. 421–437. Дои:10.1007/978-1-4020-9053-0_11. ISBN 978-1-4020-1443-7.
- ^ Андрес, Брайан; Myers, Timothy S. (23 September 2013). «Птерозавры одинокой звезды». Труды по науке о Земле и окружающей среде Королевского общества Эдинбурга. 103 (3–4): 383–398. Дои:10.1017 / S1755691013000303.
- ^ S. Meyers, Timothy (November 2010). "Earliest occurrence of the Pteranodontidae (Archosauria: Pterosauria) in North America: new material from the Austin Group of Texas". Журнал палеонтологии. 84 (6): 1071–1081. Дои:10.1666/09-082.1. S2CID 129049242.
- ^ Myers, T.S. (2010). "Earliest occurrence of the Pteranodontidae (Archosauria: Pterosauria) in North America: New material from the Austin Group of Texas". Журнал палеонтологии. 84 (6): 1071–1081. Дои:10.1666/09-082.1. S2CID 129049242.
- ^ Майерс, Тимоти С. (2010). "Новый птерозавр орнитохейрид из верхнего мела (сеноман-турон) группы Игл Форд в Техасе" (PDF). Журнал палеонтологии позвоночных. 30 (1): 280–287. Дои:10.1080/02724630903413099. S2CID 130367018.
- ^ Holman, J. Alan (22 May 2000). Fossil Snakes of North America: Origin, Evolution, Distribution, Paleoecology. Блумингтон, Индиана: Издательство Индианского университета. п. 376.
- ^ Baird, D.; Krause, D.W. (1 May 1979). "Late Cretaceous mammals east of the North American Western Interior Seaway". Журнал палеонтологии. 53 (3). Получено 13 сентября 2016.
- ^ Denton Jr., Robert K. "Late Cretaceous Mammals of the Carolinas". gsa.confex.com. Геологическое общество Америки. Получено 1 апреля 2016.
- ^ Late Cretaceous Multituberculates of the Carolinas: My...What Big Teeth You Have!
- ^ а б Grandstaff, B. S.; Parris, D. C.; Denton Jr, R. K.; Gallagher, W. B. (1992). "Alphadon (Marsupialia) and Multituberculata (Allotheria) in the Cretaceous of eastern North America". Журнал палеонтологии позвоночных. 12 (2): 217–222. Дои:10.1080/02724634.1992.10011450.
- ^ Denton, R. K. Jr., & O’Neill, R. C., 2010, A New Stagodontid Metatherian from the Campanian of New Jersey and its implications for a lack of east-west dispersal routes in the Late Cretaceous of North America. Jour. Vert. Paleo. 30(3) supp.
- ^ Martin, JE; Case, JA; Jagt, JWM; Schulp, AS; Mulder, EWA (2005). "A new European marsupial indicates a Late Cretaceous high latitude dispersal route". Mammal. Evol. 12 (3–4): 495–511. Дои:10.1007 / s10914-005-7330-х. S2CID 39202343.
- ^ Краузе, Дэвид В .; Baird, Donald (May 1979). "Late Cretaceous Mammals East of the North American Western Interior Seaway". Журнал палеонтологии. 53 (3): 562–565. JSTOR 1303997.
- ^ Emry, Robert J.; Арчибальд, Дж. Дэвид; Smith, Charles C. (September 1982). "A Mammalian Molar from the Late Cretaceous of Northern Mississippi". Журнал палеонтологии. 55 (5): 953–956. JSTOR 1304518.
- ^ Grandstaff, Barbara S.; Parris, David C.; Denton Jr., Robert K.; Gallagher, William B. (10 June 1992). "Alphadon (Marsupialia) and Multituberculata (Allotheria) in the Cretaceous of Eastern North America". Журнал палеонтологии позвоночных. 12 (2): 217–222. Дои:10.1080/02724634.1992.10011450. JSTOR 4523441.
- ^ "OCEANS OF KANSAS PALEONTOLOGY Fossils from the Late Cretaceous Western Interior Sea". Получено 12 июн 2019.
- ^ Everhart, Mike. "A Field Guide to Fossils of the Smoky Hill Chalk Part 2: Sharks and Bony Fish". Oceans of Kansas. Получено 12 июн 2019.
- ^ Everhart, Mike. "A Field Guide to the Smoky Hill Chalk Part 1: Invertebrates". Oceans of Kansas. Получено 12 июн 2019.
- ^ Everhart, Mike. "A Field Guide to Fossils of the Smoky Hill Chalk Part 3: Marine Reptiles". Oceans of Kansas. Получено 12 июн 2019.
- ^ Everhart, Mike. "A Field Guide to Fossils of the Smoky Hill Chalk Part 4: Pteranodons, Birds, and Dinosaurs". Oceans of Kansas. Получено 12 июн 2019.
- ^ Everhart, Mike. "A Field Guide to Fossils of the Smoky Hill Chalk Part 5: Coprolites, pearls, fossilized wood and other remains". Oceans of Kansas. Получено 12 июн 2019.
- ^ Everhart, Mike. "M. J. EVERHART'S MARINE REPTILE REFERENCES: MOSASAURS, PLESIOSAURS, TURTLES AND OTHER VERTEBRATE FAUNA". Oceans of Kansas. Получено 12 июн 2019.
- ^ Everhart, Mike. "Other references about fossils from the Late Cretaceous Western Interior Sea, including: Invertebrates, sharks and fish". Oceans of Kansas. Получено 12 июн 2019.
- ^ Kiernan, Caitlin R. Kiernan (2002). "Stratigraphic distribution and habitat segregation of mosasaurs in the Upper Cretaceous of western and central Alabama, with an historical review of alabama mosasaur discoveries". Журнал палеонтологии позвоночных. 22 (1): 91–103. Дои:10.1671 / 0272-4634 (2002) 022 [0091: SDAHSO] 2.0.CO; 2. ISSN 0272-4634.
- ^ KIERNAN, CAITLIN R. (2002). "Stratigraphic distribution and habitat segregation of mosasaurs in the Upper Cretaceous of western and central Alabama, with an historical review of alabama mosasaur discoveries". Журнал палеонтологии позвоночных. 22 (1): 91–103. Дои:10.1671 / 0272-4634 (2002) 022 [0091: SDAHSO] 2.0.CO; 2.
- ^ Gallagher, William B.; Campbell, Carl E.; Jagt, John W. M .; Mulder, Eric W. A. (2005). "Mosasaur (Reptilia, Squamata) Material from the Cretaceous-Tertiary Boundary Interval in Missouri". Журнал палеонтологии позвоночных. 25 (2): 473–475. Дои:10.1671/0272-4634(2005)025[0473:MRSMFT]2.0.CO;2. JSTOR 4524462.
- ^ Rapp, William F. (1946). "Check List of the Fossil Fishes of New Jersey". Журнал палеонтологии. 20 (5): 510–513. JSTOR 1299280.
- ^ Case, Gerald R.; Schwimmer, David R. (March 1988). "Late Cretaceous fish from the Blufftown Formation (Campanian) in western Georgia". Журнал палеонтологии. 62 (2): 290–301. Дои:10.1017/S0022336000029942. S2CID 131268461.
- ^ Cockerell, T. D. A.; Stanton, T. W. (1915). "SOME AMERICAN CRETACEOUS FISH SCALES, WITH NOTES ON THE CLASSIFICATION AND DISTRIBUTION OF -CRETACEOUS FISHES" (PDF). Бюллетень геологической службы США. 603: 34–57. Получено 20 декабря 2019.
- ^ Stringer, Gary L.; Oman, Luke D.; Badger, Robert F. (28 November 2016). "Woodbury Formation (Campanian) in New Jersey yields largest known Cretaceous otolith assemblage of teleostean fishes in North America". Труды Академии естественных наук Филадельфии. 165 (1): 15–36. Дои:10.1635/053.165.0101. S2CID 132425261.
- ^ Lauginiger, Edward M. (September 1988). "Cretaceous Fossils from the Chesapeake and Delaware Canal" (PDF). Геологическая служба Делавэра: 61.
- ^ Lidgard, Scott; Crane, Peter R. (Winter 1990). "Angiosperm Diversification and Cretaceous Floristic Trends: A Comparison of Palynofloras and Leaf Macrofloras". Палеобиология. 16 (1): 77–93. Дои:10.1017/S009483730000974X. JSTOR 2400934.
- ^ "22A; Veatch & Stephenson 1911 Cretaceous Material". Georgias Fossils. Получено 11 июн 2019.
- ^ Murray (1974). Миннесота. С. 162–163.
- ^ Sims, Hallie J.; Херендин, Патрик С .; Lupia, Richard; Christopher, Raymond A.; Crane, Peter R. (August 1999). "Fossil flowers with Normapolles pollen from the Upper Cretaceous of southeastern North America". Обзор палеоботаники и палинологии. 106 (3–4): 131–151. Дои:10.1016/s0034-6667(99)00008-1.
- ^ V. Demers‐Potvin, Alexandre; C. E. Larsson, Hans (2 August 2019). «Палеоклиматическая реконструкция сеноманской флоры покрытосеменных растений около Шеффервилля, Лабрадор». Палеонтология. 62 (6): 1027–1048. Дои:10.1111/pala.12444.
- ^ Херендин, Патрик С .; Magallon-Puebla, Susana; Lupia, Richard; R. Crane, Peter; Kobylinska, Jolanta (1999). "A Preliminary Conspectus of the Allon Flora from the Late Cretaceous (Late Santonian) of Central Georgia, U.S.A.". Annals of the Missouri Botanical Garden. 86 (2): 407–471. Дои:10.2307/2666182. JSTOR 2666182.
- ^ Крейн, Питер Р .; Dilcher, David L. (1984). "Lesqueria: An Early Angiosperm Fruiting Axis From the Mid-Cretaceous". Annals of the Missouri Botanical Garden. 71 (2): 384–402. Дои:10.2307/2399031. JSTOR 2399031.
- ^ Blackwell, W. H. (January 1984). "Fossil Ponderosa-like Pine Wood from the Upper Cretaceous of North-east Mississippi". Анналы ботаники. 53 (1): 133–136. Дои:10.1093/oxfordjournals.aob.a086664. JSTOR 42756870.
- ^ Mickell, James E. (Spring 1996). "Grexlupus carolinensis, A NEW PROBABLE LAURACEOUS FRUIT FROM THE LATE CRETACEOUS OF NORTH CAROLINA". Журнал Научного общества Элиши Митчелла. 112 (1): 1–6. JSTOR 24335383.
- ^ Frederiksen, Norman O. (January 1989). "Late Cretaceous and Tertiary Floras, Vegetation, and Paleoclimates of New England". Родора. 91 (865): 25–48. JSTOR 23312460.
- ^ Berry, Edward Wilbur (1914). "The Upper Cretaceous and Eocene floras of South Carolina and Georgia". Типография правительства США. 84.
{kind=link}
{kind=link}
{kind=link}