Кинуренин-3-монооксигеназа - Kynurenine 3-monooxygenase
| кинуренин-3-монооксигеназа | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Структура димера кинуренин-3-монооксигеназы, полученного из 4J34.[1] Один мономер изображен в мультяшном формате (голубой), а второй - в виде ленты (зеленый). Гибкие линкерные области (остатки 96-104) окрашены в красный цвет. Флавинадениндинуклеотид (FAD) показан в виде сфер с цветовой кодировкой в соответствии с типом атома. | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 1.14.13.9 | ||||||||
| Количество CAS | 9029-61-2 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
В энзимология, а кинуренин-3-монооксигеназа (ЕС 1.14.13.9 ) является фермент который катализирует в химическая реакция
- L-кинуренин + НАДФН + ЧАС+ + О2 ⇌ 3-гидрокси-L-кинуренин + НАДФ+ + ЧАС2О
Кинуренин-3-монооксигеназа является продуктом экспрессии КМО (ген). В систематическое название этого класса ферментов L-кинуренин, НАДФН: оксидоредуктаза кислорода (3-гидроксилирование). Другие широко используемые имена включают кинуренин-3-гидроксилаза, кинуренингидроксилаза, и L-кинуренин-3-гидроксилаза. Он участвует в метаболизм триптофана сквозь кинурениновый катаболический путь. Этот фермент принадлежит к семейству оксидоредуктазы, а точнее, те, кто действует от парных доноров, с О2 как окислитель. Кинуренин-3-монооксигеназа катализирует внедрение молекулярного кислорода в ароматическое кольцо кинуренина с образованием 3-гидрокси-L-кинуренин.[2] Здесь работает один кофактор, FAD. Кинуренин-3-монооксигеназа служит важной точкой ветвления в пути кинуренина и, как следствие, является привлекательной лекарственной мишенью для иммунологический, нейродегенеративный, и нейровоспалительные заболевания.[3] В настоящее время большинство исследований фермента кинуренин-3-монооксигеназы сосредоточено в основном на крыса модели[4] И в дрожжи,[5] оба из которых имеют высокую последовательность гомология с белком кинуренин-3-монооксигеназа человека. Исследования показали положительные эффекты ингибирования ферментов в этих эукариотических активных центрах кинуренин-3-монооксигеназы, что делает этот фермент привлекательной мишенью для разработки лекарственных препаратов для человека.[3][5]
Структура
Кинуренин-3-монооксигеназа представляет собой димер содержащий асимметричный подразделения[5] и имеет один FAD-связывающий домен в качестве своей простетической группы.[3] Кинуренин-3-монооксигеназа содержит линкерную область, участвующую в связывании субстрата после второй цепи антипараллельной β-лист, шестицепочечный антипараллельный β-листовой домен и α-спираль на карбокси-конце.[5] В гидрофобный С-конец действует как заякоренный домен митохондрий и участвует в ферментативной активности.[6]
Активный сайт
В то время как никакая научная литература не сообщает о кристаллическом изображении комплекса кинуренин-3-монооксигеназы с L-кинуренин, структурные исследования фермента в дрожжах, совместно кристаллизованного с UPF 648, показывают, как кофактор FAD и субстрат связываются в активном центре.[1] Химическое сходство между UPF 648 и L-кинуренин предполагает, что субстрат связывается рядом с Re-лицо флавопротеина. Цикл, содержащий вычеты Pro321–Gln325 Полагают, что это сайт связывания кислорода над обратной стороной простетической группы FAD.[5]
Каждый мономер содержит консервативный гидрофобный карман (остатки Leu221, Встретились230, Иль232, Лей234, Phe246, Pro321, Phe322), расположенный вокруг ароматической бензольной части субстрата.[5] Консервированный Gln325 полярный остаток также участвует в образовании водородных связей на L-кинуренин карбонильная группа, а также водород на атоме FAD N3.[1] Arg83 и Тир97 также образуют полярные контакты с карбоксилатом в аминокислотной части субстрата.[7]
Механизм
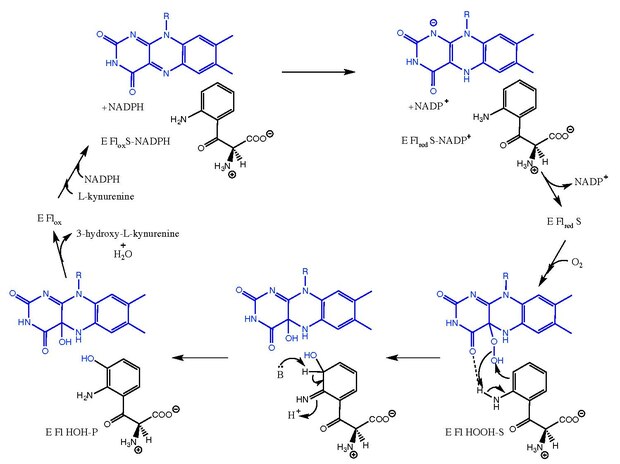
Кинуренин-3-монооксигеназа катализирует гидроксилирование из L-кинуренин до 3-гидрокси-L-кинуренин с сопутствующим взаимным превращением НАДФН в НАДФ+. Механизм реакции не совсем известен, но считается, что он следует за механизмами, связанными с флавин-зависимые монооксигеназы.[8] После L-кинуренин связывает, НАДФН снижает ФАД и уходит в виде НАДФ+. Затем кислород связывается и создает Lпромежуточное соединение -кинуренин-FAD-гидропероксид.[5][9] Этот промежуточный продукт является электрофильным источником для реакции гидроксилирования, давая первичный кетимин форма продукта и C4a-гидрокси-FAD.[10] Таутомеризация дает 3-гидрокси-L-кинуренин в комплексе с ферментом (E Fl HOH-P). Диссоциация 3-гидрокси-L-кинуренин и H2O приводит к свободному ферменту (E Flбык).
Биологическая функция
Кинуренин-3-монооксигеназа катализирует превращение L-кинуренин до 3-гидрокси-L-кинуренин, важный биоактивный метаболит в кинурениновый путь. Кинурениновый путь отвечает за более 95% триптофан окислительная деструкция.[11] L-Кинуренин является важной точкой разветвления этого метаболического пути, преобразуясь в нейротоксин 3-гидрокси-L-кинуренин через кинуренин-3-монооксигеназу, нейропротектор кинуреновая кислота через кинуренин-аминотрансферазы, или антраниловая кислота к кинурениназа.[12]
Кинуренин-3-монооксигеназа регулирует последующее производство хинолиновая кислота, которые могут генерировать реактивные свободные радикалы[13] и активирует NMDA подтип глутаматных рецепторов, вызывающих эксайтотоксические поражения в центральной нервной системе млекопитающих.[14][15] Хинолиновая кислота также является биопредшественником НАД.+.[12]
Ингибирование кинуренин-3-монооксигеназы приводит к увеличению кинурениновой кислоты в кинурениновом пути. Этот метаболит действует как антагонист из α7 никотиновый ацетилхолиновый рецептор и как антагонист на глициновом сайте рецептора NMDA.[16] В результате регуляция фермента кинуренин-3-монооксигеназы определяет нейротоксический и нейрозащитный потенциал кинуренинового пути.
Актуальность болезни
Кинуренин-3-монооксигеназа является привлекательной лекарственной мишенью для нескольких нейродегенеративных и нейровоспалительных заболеваний, особенно Хантингтона, Болезнь Альцгеймера, и болезнь Паркинсона. Введение сильнодействующих ингибиторов ферментов продемонстрировало многообещающие фармакологические результаты.[3][5] В частности, было показано, что генетическое устранение фермента кинуренин-3-монооксигеназы подавляет токсичность Хантингтин белок в дрожжах[17] и Дрозофила[18] модели болезни Хантингтона.
Дефицит кинуренин-3-монооксигеназы, который может быть вызван генетическими полиморфизмы, цитокины, или оба,[19] приводит к накоплению кинуренин и сдвиг в триптофане метаболический путь по отношению к кинуреновой кислоте и антраниловая кислота. Недавние исследования показывают, что гиперфизиологические концентрации кинуренин у пациентов с дефицитом кинуренин-3-монооксигеназы приводит к сдвигу в сторону производства кинуреновой кислоты, что, как полагают, связано с когнитивным дефицитом в предсказательное преследование и зрительно-пространственная рабочая память.[20] Дефицит кинуренин-3-монооксигеназы связан с заболеваниями головного мозга (например, шизофрения, тиковые расстройства ) и печени.[21][22][23][24][25]
Рекомендации
- ^ а б c Амарал, М. (2014). «Кристаллическая структура кинуренин-3-монооксигеназы - усеченная в положении 394 плюс расщепленная метка HIS». Дои:10.2210 / pdb4j34 / pdb. Цитировать журнал требует
| журнал =(помощь) - ^ Филиппини, Грациэлла Аллегри; Коста, Карло В. Л .; Бертаццо, Антонелла; Международное совещание по исследованиям триптофана (1998 г.). «Последние достижения в исследованиях триптофана: пути триптофана и серотонина». Достижения экспериментальной медицины и биологии. 398. Дои:10.1007/978-1-4613-0381-7. ISBN 978-1-4613-8026-9. S2CID 38080353.
- ^ а б c d Смит, Джейсон Р .; Джейми, Джоанн Ф .; Гийемен, Жиль Ж. (февраль 2016 г.). «Кинуренин-3-монооксигеназа: обзор структуры, механизма и ингибиторов». Открытие наркотиков сегодня. 21 (2): 315–324. Дои:10.1016 / j.drudis.2015.11.001. ISSN 1359-6446. PMID 26589832.
- ^ Horn, U .; Ullrich, V .; Штаудингер, HJ (1971). "Очистка и характеристика L-кинуренин-3-гидроксилаза (EC 1.14.1.2.) из печени крысы ». Hoppe-Seyler's Z. Physiol. Chem. 352 (6): 837–842. Дои:10.1515 / bchm2.1971.352.1.837. PMID 5087636.
- ^ а б c d е ж грамм час я Амарал, М; Леви, К; Привет, диджей; Lafite, P; Отейро, TF; Джорджини, F; Leys, D; Скраттон, Н.С. (2013). «Структурные основы ингибирования кинуренин-3-монооксигеназы». Природа. 496 (7445): 382–385. Bibcode:2013Натура.496..382A. Дои:10.1038 / природа12039. ЧВК 3736096. PMID 23575632.
- ^ Hirai KH и др. (2010). «Двойная роль карбоксиконцевой области печени свиньи L-кинуренин-3-монооксигеназа: сигнал, направленный на митохондрии, и ферментативная активность ». J. Biochem. 148 (6): 639–650. Дои:10.1093 / jb / mvq099. PMID 20802227.
- ^ Mole D и др. (2016). «Ингибирование кинуренин-3-монооксигеназы предотвращает полиорганную недостаточность на моделях острого панкреатита на грызунах». Природа Медицина. 22 (2): 202–209. Дои:10,1038 / нм 4020. ЧВК 4871268. PMID 26752518.
- ^ Бретон, Дж .; и другие. (2000). «Функциональная характеристика и механизм действия рекомбинантной кинуренин-3-гидроксилазы человека». Евро. J. Biochem. 267 (4): 1092–1099. Дои:10.1046 / j.1432-1327.2000.01104.x. PMID 10672018.
- ^ а б Крозье-Рибе, КР; и другие. (2008). "Кинуренин-3-монооксигеназа из Pseudomonas fluorescens: субстратоподобные ингибиторы как стимулируют восстановление флавина, так и стабилизируют промежуточный флавин-пероксо, но при этом приводят к образованию пероксида водорода ». Биохимия. 47 (47): 12420–12433. Дои:10.1021 / bi8010434. PMID 18954092.
- ^ а б Entsch B, et al. (1976). «Флавин-кислородные производные, участвующие в гидроксилировании п-гидроксибензоатгидроксилаза ». J. Biol. Chem. 251 (9): 2550–2563. PMID 816794.
- ^ Thevandavakkam MA, et al. (2010). «Ориентация на кинуренин-3-монооксигеназу (КМО). Значение для терапии при болезни Хантингтона». CNS Neurol. Disord. Цели наркотиков. 9 (6): 791–800. Дои:10.2174/187152710793237430. PMID 20942784.
- ^ а б Giorgini F, Huang SY, Sathyasaikumar KV и др. (2013). «Целенаправленное удаление кинуренин-3-монооксигеназы у мышей: новый инструмент для изучения метаболизма кинуренинового пути в периферии и головном мозге». Журнал биологической химии. 288 (51): 36554–36566. Дои:10.1074 / jbc.M113.503813. ЧВК 3868768. PMID 24189070.
- ^ Rios, C .; Сантамария, А. (1991). «Хинолиновая кислота является сильнодействующим пероксидантом липидов в гомогенатах головного мозга крысы». Neurochem. Res. 16 (10): 1139–1143. Дои:10.1007 / bf00966592. PMID 1686636. S2CID 28669340.
- ^ Stone, T. W .; Перкинс, М. Н. (1981). «Хинолиновая кислота: мощный эндогенный возбудитель аминокислотных рецепторов в ЦНС». Евро. J. Pharmacol. 72 (4): 411–412. Дои:10.1016/0014-2999(81)90587-2. PMID 6268428.
- ^ Schwarcz, R .; Bruno, J. P .; Muchowski, P.J .; Ву, Х.К. (2012). «Кинуренины в мозге млекопитающих. Когда физиология встречается с патологией». Nat. Преподобный Neurosci. 13 (7): 465–477. Дои:10.1038 / номер 3257. ЧВК 3681811. PMID 22678511.
- ^ Hilmas, C .; Pereira, E. F .; Алкондон, М .; Rassoulpour, A .; Schwarcz, R .; Альбукерке, Э. X. (2001). «Метаболит мозга кинуреновая кислота ингибирует активность никотинового рецептора α7 и увеличивает экспрессию никотинового рецептора, отличного от α7: физиопатологические последствия». J. Neurosci. 21 (19): 7463–7473. Дои:10.1523 / JNEUROSCI.21-19-07463.2001. ЧВК 6762893. PMID 11567036.
- ^ Giorgini, F .; Guidetti, P .; Nguyen, Q .; Bennett, S.C .; Муховски, П. Дж. (2005). «Геномный скрининг дрожжей показывает, что кинуренин-3-монооксигеназа является терапевтической мишенью для лечения болезни Хантингтона». Nat. Genet. 37 (5): 526–531. Дои:10,1038 / ng1542. ЧВК 1449881. PMID 15806102.
- ^ Campesan, S .; Green, E.W .; Breda, C .; Sathyasaikumar, K. V .; Muchowski, P.J .; Schwarcz, R .; Kyriacou, C.P .; Джорджини, Ф (2011). "Путь кинуренина модулирует нейродегенерацию в Дрозофила модель болезни Хантингтона ". Curr. Биол. 21 (11): 961–966. Дои:10.1016 / j.cub.2011.04.028. ЧВК 3929356. PMID 21636279.
- ^ Müller, N; Myint, AM; Шварц, MJ (2010). «Воспалительные биомаркеры и депрессия». Нейротокс. Res. 19 (2): 308–318. Дои:10.1007 / s12640-010-9210-2. PMID 20658274. S2CID 3225744.
- ^ Воноди И., Колин-Стайн О., Сатьясайкумар К.В. и др. (2011). «Снижение экспрессии гена кинуренин-3-монооксигеназы и ферментативной активности при шизофрении и генетической ассоциации с эндофенотипами шизофрении». Arch Gen Psychiatry. 68 (7): 665–674. Дои:10.1001 / archgenpsychiatry.2011.71. ЧВК 3855543. PMID 21727251.
- ^ Хольце М., Сэтре П., Энгберг Г. и др. (2012). «Полиморфизмы кинуренин-3-монооксигеназы: значение для синтеза кинуреновой кислоты у пациентов с шизофренией и здоровых людей». J Psychiatry Neurosci. 37 (1): 53–57. Дои:10.1503 / jpn.100175. ЧВК 3244499. PMID 21693093.
- ^ Кэмпбелл, Брайан М .; Чарыч, Эрик; Ли, Анна В .; Мёллер, Томас (2014). «Кинуренины при заболевании ЦНС: регуляция воспалительными цитокинами». Границы неврологии. 8 (12): 12. Дои:10.3389 / fnins.2014.00012. ЧВК 3915289. PMID 24567701.
- ^ Hoekstra, PJ; Андерсон, GM; Troost, PW (2007). «Плазменный кинуренин и связанные с ним меры у пациентов с тиковым расстройством». Eur Детская подростковая психиатрия. 16 (Приложение 1): 71–77. Дои:10.1007 / s00787-007-1009-1. PMID 17665285. S2CID 39150343.
- ^ Бунесс А., Рот А., Херрманн А., Шмитц О., Камп Н. и др. (2014). «Идентификация метаболитов, маркеров клинической химии и транскриптов, связанных с гепатотоксичностью». PLOS ONE. 9 (5): e97249. Bibcode:2014PLoSO ... 997249B. Дои:10.1371 / journal.pone.0097249. ЧВК 4023975. PMID 24836604.
- ^ Юкико, Хирата; Такаши, Кавачи; Такаши, Сугимура (2 октября 1967 г.). "Жирная печень, вызванная инъекцией L-триптофан ». Biochimica et Biophysica Acta (BBA) - липиды и липидный метаболизм. 144 (2): 233–241. Дои:10.1016/0005-2760(67)90153-1. PMID 4168935.
- Фушер А.Л., Макинтош А., Дус Дж., Вастлинг Дж., Тейт А., Тернер С.М. (2006). «Протеомный анализ лекарственной устойчивости к мышьяку у Trypanosoma brucei». Протеомика. 6 (9): 2726–2732. Дои:10.1002 / pmic.200500419. PMID 16526094. S2CID 24074942.
- Окамото Х., Хаяиси О (1967). «Потребность в флавинадениндинуклеотиде для кинуренингидроксилазы митохондрий печени крысы». Biochem. Биофиз. Res. Сообщество. 29 (3): 394–399. Дои:10.1016 / 0006-291X (67) 90469-X. PMID 6076241.
- Сайто Ю., Хаяиши О., Ротберг С. (1957). "Исследования оксигеназ; ферментативное образование 3-гидрокси-L-кинуренин из L-кинуренин ". J. Biol. Chem. 229 (2): 921–934. PMID 13502353.