Пространственная память - Spatial memory
Эта статья может требовать уборка встретиться с Википедией стандарты качества. (Сентябрь 2011 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |

В когнитивная психология и нейробиология, пространственная память это форма памяти, отвечающая за запись информации об окружающей среде и пространственной ориентации. Например, пространственная память человека необходима, чтобы перемещаться по знакомому городу, точно так же, как пространственная память крысы необходима для того, чтобы узнать местонахождение еды в конце пути. лабиринт. Часто утверждают, что и у людей, и у животных пространственная память резюмируется как когнитивная карта.
Пространственная память имеет представления внутри работы, краткосрочная память и Долгосрочная память. Исследования показывают, что с пространственной памятью связаны определенные области мозга. Многие методы используются для измерения пространственной памяти у детей, взрослых и животных.
Кратковременная пространственная память
Краткосрочная память (STM) можно описать как систему, позволяющую временно хранить и управлять информацией, необходимой для выполнения сложных когнитивных задач.[1] Задачи, использующие кратковременную память, включают: учусь, рассуждение, и понимание.[1] Пространственная память - это когнитивный процесс, который позволяет человеку запоминать разные места, а также пространственные отношения между объектами.[1] Это позволяет запомнить, где находится объект по отношению к другому объекту;[1] например, позволяя кому-то перемещаться через знакомый город. Говорят, что пространственные воспоминания формируются после того, как человек уже собрал и обработал сенсорный информация о ее или его окружении.[1]
Пространственная рабочая память
Рабочая память (WM) можно описать как систему с ограниченными возможностями, позволяющую временно хранить и обрабатывать информацию.[2] Это временное хранилище позволяет выполнять сложные задачи или работать над ними, сохраняя при этом информацию.[2] Например, способность работать над сложной математической задачей использует рабочую память.
Одна очень влиятельная теория WM - это Baddeley и сцепка многокомпонентная модель рабочей памяти.[2][3] Самая последняя версия этой модели предполагает, что есть четыре подкомпонента WM, а именно: фонологическая петля; в визуально-пространственный блокнот; в центральная исполнительная власть; и эпизодический буфер.[2] Считается, что один из компонентов этой модели, визуально-пространственный блокнот, отвечает за временное хранение, обслуживание и манипулирование как визуальной, так и пространственной информацией.[2][3]
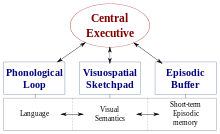
В отличие от многокомпонентной модели, некоторые исследователи считают, что СТМ следует рассматривать как единую конструкцию.[3] В этом отношении считается, что визуальная, пространственная и вербальная информация организована по уровням представления, а не по типу магазина, к которому они принадлежат.[3] В литературе предлагается изучить дальнейшие исследования фракционирования STM и WM.[3][4] Однако большая часть исследований конструкции зрительно-пространственной памяти проводилась в соответствии с парадигмой, предложенной Баддели и Хитчем.[2][3][4][5][6]
Роль центральной исполнительной власти
Исследования точной функции зрительно-пространственного блокнота показали, что как пространственные краткосрочная память и рабочая память зависят от исполнительных ресурсов и не совсем различны.[2] Например, производительность рабочей памяти, но не задачи краткосрочной памяти, зависит от артикуляционное подавление предполагая, что ухудшение пространственной задачи было вызвано одновременным выполнением задачи, которая широко использовала исполнительные ресурсы.[2] Результаты также показали, что производительность была снижена в задачах STM и WM с подавлением руководства.[2] Это показывает, как в визуально-пространственной области и STM, и WM требуют одинаковой полезности центрального исполнительного органа.[2]
Кроме того, во время задачи пространственной визуализации (которая связана с исполнительным функционированием, а не с STM или WM) одновременное подавление исполнительных функций ухудшало производительность, указывая на то, что эффекты были вызваны общими требованиями к центральному исполнительному органу, а не краткосрочным хранилищем.[2] Исследователи пришли к выводу, что центральная исполнительная власть использует когнитивные стратегии позволяя участникам как кодировать, так и поддерживать ментальные представления во время задач краткосрочной памяти.[2]
Хотя исследования показывают, что центральный исполнительный орган принимает непосредственное участие в решении ряда пространственных задач, точный способ их взаимосвязи еще предстоит выяснить.[7]
Долговременная пространственная память
Вызов пространственной памяти построен на иерархическая структура. Это означает, что люди запоминают общую планировку конкретного пространства, а затем «указывают целевые местоположения» в этом пространственном наборе.[8] Эта парадигма включает порядковую шкалу характеристик, на которые должен обратить внимание человек, чтобы сформировать свою когнитивную карту.[9] Воспоминание пространственных деталей - это нисходящая процедура, которая требует от человека вспомнить главные черты когнитивной карты, за которыми следуют ордината и подчиненные черты. Таким образом, при навигации по маршруту выделяются две пространственные особенности: общий план и ориентир (Kahana et al., 2006).
Люди не только способны узнавать о пространственном расположении своего окружения, но также могут соединять новые маршруты и новые пространственные отношения посредством логического вывода. Тем не менее, этой области традиционно препятствовали смешивающие переменные, такие как стоимость и возможность предыдущего воздействия экспериментальной среды. К счастью, технологические скачки открыли для психологов новый, хотя и виртуальный, мир.
Когнитивная карта - это «мысленная модель пространственной конфигурации объектов, позволяющая перемещаться по оптимальному пути между произвольными парами точек».[10] Эта ментальная карта построена на двух фундаментальных основах: планировке, также известной как знание маршрута, и ориентации на ориентир. Макет потенциально является первым методом навигации, который люди учатся использовать; его работа отражает наши самые основные представления о мире.
Хермер и Спелке (1994) определили, что когда малыши начинают ходить, примерно в восемнадцать месяцев, они ориентируются в соответствии со своим чувством планировки мира. Действительно, может показаться, что мир странствующего малыша - это место осевых линий и контрастных границ. Макнамара, Харди и Хиртл определили членство в регионе как главный строительный блок любой когнитивной карты (1989). В частности, принадлежность к региону определяется любым видом границ, будь то физическая, перцептивная или субъективная (McNamara et al., 1989). Границы являются одними из самых основных и эндемичных качеств окружающего нас мира. Эти границы представляют собой не что иное, как осевые линии, к которым люди склонны относиться к пространству; например, одним из определяющих факторов осевой линии является сила тяжести (McNamara & Shelton, 2001; Kim & Penn, 2004). Осевые линии помогают каждому распределить наше восприятие по регионам. Эта идея разделенного мира дополнительно подкрепляется тем фактом, что элементы, которые вспоминаются вместе, с большей вероятностью также будут сгруппированы в одной и той же области более крупной когнитивной карты.[9] Кластеризация показывает, что люди склонны собирать информацию вместе в соответствии с более мелкими схемами на более крупной когнитивной карте.
Однако границы - не единственные определяющие факторы макета. Кластеризация также демонстрирует еще одно важное свойство нашего отношения к пространственным представлениям. Это то, что пространственное воспоминание - это иерархический процесс. Когда кто-то вспоминает среду или перемещается по местности, этот человек сначала неявно вспоминает общий план. Затем, благодаря «богатой корреляционной структуре» концепции, активируется ряд ассоциаций.[8] В конце концов, результирующий каскад активаций пробудит конкретные детали, соответствующие вызываемой области. Вот как люди кодируют множество сущностей с разных онтологических уровней, таких как расположение степлера; в письменном столе; который находится в офисе .. Увы, есть и свои недостатки. Можно вспомнить только из одного региона за раз (узкое место).
Узкое место в когнитивной навигационной системе человека может иметь катастрофические последствия, например, если возникнет необходимость во внезапном объезде во время долгой поездки. И все же люди по-прежнему способны найти место для проживания функционально. Отсутствие опыта работы в какой-либо местности или просто огромный размер могут дезориентировать человека, особенно в большом и незнакомом месте со многими подавляющими раздражителями. В этих условиях люди все еще могут ориентироваться и даже ориентироваться по ориентирам. Эта способность «расставлять приоритеты для объектов и регионов в сложных сценах для выбора (и) распознавания» была названа Чун и Цзян в 1998 году. Ориентиры дают людям ориентиры, активируя «усвоенные ассоциации между глобальным контекстом и целевыми местоположениями».[8] Mallot и Gillner (2000) показали, что испытуемые усвоили связь между конкретным ориентиром и направлением поворота, тем самым способствуя установлению связи между ассоциациями и ориентирами.[11] Шелтон и Макнамара (2001) кратко резюмировали, почему ориентиры, как маркеры, так полезны: «местоположение ... не может быть описано без ссылки на ориентацию наблюдателя».
Совершенно очевидно, что люди используют как планировку определенного пространства, так и наличие ориентиров для навигации. Тем не менее, психологам еще предстоит объяснить, влияет ли макет на ориентиры или ориентиры определяют границы макета. Таким образом, эта концепция страдает парадоксом курицы и яйца. Фактически, Макнамара обнаружил, что субъекты используют «группы ориентиров как внутренние системы координат», что еще больше запутывает проблему.[10]
Люди воспринимают объекты в своей среде по сравнению с другими объектами в той же среде. Другими словами, ориентиры и расположение - дополнительные системы для пространственного воспоминания. Однако неизвестно, как эти две системы взаимодействуют, когда доступны оба типа информации. Таким образом, мы должны сделать определенные предположения о взаимодействии между этими двумя системами. Например, когнитивные карты не являются «абсолютными», а скорее, как любой может подтвердить, «используются для предоставления значения по умолчанию ... (которое) модулируется в соответствии с ... требованиями задачи».[8] Психологи также считают, что когнитивные карты основаны на конкретных примерах, что объясняет «различительное соответствие с прошлым опытом».[8]
Эти предположения вскоре могут быть подтверждены. Достижения в области технологий виртуальной реальности открыли дверь в эту загадочную область. Теперь экспериментаторы обнаруживают, что создают сценарии, которые невозможно было представить пятнадцать лет назад. Виртуальная реальность дает экспериментаторам роскошь полного контроля над своей тестовой средой. Можно манипулировать любой переменной, включая вещи, которые в реальности были бы невозможны.
Виртуальная реальность
Во время одного из недавних исследований исследователи спроектировали три разных виртуальных города, каждый из которых имел свою «уникальную планировку дорог и уникальный набор из пяти магазинов».[10] Однако общий размер различных карт был одинаковым - «80 кв. Единиц». В этом эксперименте участники должны были принять участие в двух разных наборах испытаний.
Сначала участникам были назначены два из трех виртуальных ландшафтов и поручена роль таксиста. Аватар участника поднял в общей сложности 25 пассажиров и высадил их в случайных местах по всей карте. Исследователи измерили «длину пути доставки» участников в обоих наборах первого испытания. Они обнаружили «явное уменьшение длины пути с увеличением количества доставок в городе». Увеличение длины маршрута участниками коррелировало с увеличением опыта вождения в виртуальных городах. Однако, как и следовало ожидать, знания, которые участники почерпнули из первого города, не передавали и не помогали им в их путешествиях по второму городу. Таким образом, Newman et al. (2006) пришли к выводу, что участники «сформировали обзорное представление каждого города». То есть они объехали достаточно города, чтобы составить общий план остальной его части.
Второй набор испытаний включал ту же задачу для участников, но с некоторыми изменениями в контексте окружающей среды. Исследователи добавили еще две карты и уменьшили их. Второй набор карт в этом испытании был стандартизирован с тем же макетом и ориентирами, что означает, что все участники испытают эту карту во время второго запуска. Более того, три из пяти карт «заменили некоторые ориентиры новыми ориентирами, в то время как расположение целевых местоположений осталось (редактировалось) без изменений». Две другие карты были либо «идентичны» второму пройденному ландшафту второго испытания, либо были изменены как ориентиры, так и их относительное расположение. Опять же, Newman et al. измерили «избыточную длину пути», уделяя особое внимание избыточному расстоянию, пройденному при первой доставке второй карты во втором испытании, где они полагали, что пространственное обучение будет в основном передаваться с первой карты. Они обнаружили, что группа, которая столкнулась с измененными изображениями зданий в тех же местах, имела самый высокий уровень передачи знаний о планировке из первого города.
Результаты первого испытания показали, что люди способны изучать пространственное расположение интерактивной среды. Второе испытание показало, что, несмотря на изменение присутствия достопримечательностей, участники все еще могли «находить новые кратчайшие маршруты». Поскольку участники не путешествовали по ранее запомненным маршрутам и по-прежнему хорошо работали в соответствии со стандартом «избыточной длины пути», Newman et al. пришел к выводу, что эти результаты свидетельствуют о «некотором обзоре окружающей среды более высокого порядка».
В исследовании, проведенном в Университете Мэриленда, сравнивалось влияние разных уровней погружения на вспоминание пространственной памяти.[12] В исследовании 40 участников использовали как традиционный рабочий стол, так и дисплей, закрепленный на голове, для просмотра двух сред: средневекового города и богато украшенного дворца, где они запомнили два набора из 21 лица, представленные в виде трехмерных портретов. После просмотра этих 21 лица в течение 5 минут с последующим коротким периодом отдыха лица в виртуальных средах были заменены числами, и участники вспомнили, какое лицо было в каждом месте. Исследование показало, что в среднем те, кто пользовался головным дисплеем, вспоминали лица на 8,8% точнее и с большей уверенностью. Участники заявляют, что использование их врожденных вестибулярных и проприоцептивных органов чувств с закрепленным на голове дисплеем и отображением аспектов окружающей среды относительно их тела, элементов, которые отсутствуют на рабочем столе, было ключом к их успеху.
Пространственная экспертиза
В литературе есть свидетельства того, что специалисты в определенной области могут выполнять задачи на память в соответствии со своими навыками на исключительном уровне.[6] Также было сказано, что уровень мастерства, продемонстрированный экспертами, превышает пределы нормальных возможностей STM и WM.[6] Считается, что, поскольку эксперты обладают огромным количеством заранее подготовленных и специфических знаний, они могут кодировать информацию более эффективным способом.[6]
Интересное исследование, исследующее такси память водителей на улицы в Хельсинки, Финляндия рассмотрели роль предварительно полученных пространственных знаний.[6] В этом исследовании эксперты сравнивали с контрольной группой, чтобы определить, как эти предварительно полученные знания в их области навыков позволяют им преодолевать ограничения возможностей STM и WM.[6] В исследовании использовалось четыре уровня пространственной случайности:
- Порядок маршрута - пространственно непрерывный маршрут[6]
- Route Random - пространственно непрерывный список, представленный случайным образом[6]
- Порядок на карте - названия улиц образуют прямую линию на карте, но без промежуточных улиц.[6]
- Map Random - улицы на карте представлены в случайном порядке[6]

Результаты этого исследования показывают, что таксисты (эксперты) отзывались об улицах выше как в условиях порядка маршрута, так и в условиях порядка карты, чем в двух случайных условиях.[6] Это указывает на то, что эксперты смогли использовать свои предварительно полученные пространственные знания для организации информации таким образом, чтобы они превзошли ограничения возможностей STM и WM.[6] Стратегия организации, которую использовали водители, известна как дробление.[6] Кроме того, комментарии, сделанные экспертами во время процедуры, указывают на то, что они использовали знание маршрута при выполнении задачи.[6] Чтобы убедиться, что они кодировали именно пространственную информацию, исследователи также представили списки в алфавитном порядке и семантический категории.[6] Однако исследователи обнаружили, что на самом деле эксперты разделили на части пространственную информацию, что позволило им преодолеть ограничения как визуально-пространственной STM, так и WM.[6]
Исследования на животных
В литературе было обнаружено, что некоторые виды Paridae и Corvidae (такой как черношапочный синица и скраб сойка ) могут использовать пространственную память, чтобы запомнить, где, когда и какой тип еды они хранят в кэше.[13] Недавние исследования на крысах и белках также показали, что они могут использовать пространственную память для поиска ранее спрятанной пищи.[13] Эксперименты с использованием радиального лабиринта позволили исследователям контролировать ряд переменных, таких как тип спрятанной еды, места, где она спрятана, интервал удерживания, а также любые запахи, которые могут исказить результаты исследования памяти.[13] В частности, исследования показали, что крысы запоминают, где они спрятали пищу и какую пищу они спрятали.[13] Это проявляется в поведении при поиске, так что крысы избирательны в том, что они чаще подходят к рукавам лабиринта, где они ранее прятали предпочтительную пищу, чем к рукавам с менее предпочтительной пищей или где еда не была спрятана.[13]
Таким образом, данные о пространственной памяти некоторых видов животных, таких как крысы, указывают на то, что они действительно используют пространственную память для поиска и извлечения скрытых запасов еды.[13]
Во время исследования с помощью GPS-слежения, чтобы узнать, где домашние кошки уходят, когда хозяева выпускают их на улицу, показал, что у кошек отличная пространственная память. Некоторые из кошек в исследовании продемонстрировали исключительную долговременную пространственную память. Одна из таких кошек, которая обычно путешествовала не дальше 200 м (660 футов) до 250 м (820 футов) от своего дома, неожиданно покинула свой дом на 1250 м (4100 футов). Первоначально исследователи думали, что это неисправность GPS, но вскоре обнаружили, что владельцы кошки уехали из города в те выходные, и что дом, в который пошла кошка, был старым домом владельца. Хозяева и кошка не жили в этом доме больше года.[14]
Визуально-пространственное различие
Logie (1995) предположил, что визуально-пространственный блокнот разбит на два подкомпонента: визуальный и пространственный.[5] Это визуальный тайник и внутренний писец соответственно.[5] Визуальный кеш - это временное хранилище визуальных данных, включающее такие параметры, как цвет и форма.[5] И наоборот, внутренний писец является механизмом репетиции визуальной информации и отвечает за информацию, касающуюся последовательности движений.[5] Хотя в литературе отмечается общее отсутствие единого мнения относительно этого различия,[4][15][16] появляется все больше свидетельств того, что эти два компонента являются отдельными и выполняют разные функции.
Зрительная память отвечает за сохранение визуальных форм и цветов (т.е. чего), тогда как пространственная память отвечает за информацию о местоположении и движении (т.е. где). Это различие не всегда однозначно, поскольку часть зрительной памяти включает пространственную информацию и наоборот. Например, память форм объекта обычно включает в себя хранение информации о пространственном расположении характеристик, которые определяют рассматриваемый объект.[15]
На практике две системы работают вместе в некоторой степени, но были разработаны разные задачи, чтобы выделить уникальные способности, задействованные в визуальной или пространственной памяти. Например, тест визуальных шаблонов (VPT) измеряет визуальный диапазон, тогда как задача Corsi Blocks измеряет пространственный диапазон. Корреляционные исследования этих двух показателей предполагают разделение между зрительными и пространственными способностями из-за отсутствия корреляции между ними как у здоровых, так и у здоровых людей. мозг поврежден пациенты.[4]
Поддержка разделения компонентов зрительной и пространственной памяти обнаруживается в экспериментах с использованием парадигма двойной задачи. Ряд исследований показал, что сохранение визуальных форм или цветов (т. Е. Визуальной информации) нарушается из-за представления нерелевантных изображений или динамического визуального шума. И наоборот, сохранение местоположения (то есть пространственной информации) нарушается только задачами пространственного отслеживания, задачами пространственного постукивания и движениями глаз.[15][16] Например, участники выполнили как VPT, так и задачу блоков Корси в эксперименте по селективному вмешательству. В течение интервала сохранения VPT субъект просматривал нерелевантные изображения (например, авангард картины). Задача пространственной интерференции требовала, чтобы участники, касаясь стимулов, следовали расположению небольших деревянных колышков, которые были скрыты за экраном. И визуальный, и пространственный интервалы были сокращены соответствующими задачами взаимодействия, подтверждая, что задача блоков Корси относится в первую очередь к пространственной рабочей памяти.[4]
Измерение
Существует множество задач, которые психологи используют для измерения пространственной памяти у взрослых, детей и животных. Эти задачи позволяют профессионалам выявлять когнитивные нарушения у взрослых и детей, а также позволяют исследователям назначать участникам различные типы лекарств и / или поражений и измерять их влияние на пространственную память.
Задача постукивания блока Corsi
Также известный как тест Corsi Span Test, этот психологический тест обычно используется для определения объема зрительно-пространственной памяти и неявных визуально-пространственных обучающих способностей человека.[17][18] Участники садятся с девятью деревянными блоками размером 3x3 см, прикрепленными перед ними на плинтус размером 25 x 30 см в стандартном случайном порядке. Эксперимент использует блоки последовательности, которую участники должны воспроизвести. Блоки пронумерованы со стороны экспериментаторов, чтобы обеспечить эффективную демонстрацию паттернов. Длина последовательности увеличивается с каждым испытанием до тех пор, пока участник больше не сможет правильно воспроизвести образец. Тест может использоваться для измерения как кратковременной, так и долговременной пространственной памяти, в зависимости от промежутка времени между тестом и воспроизведением.
Тест был создан Канадский нейропсихолог Филипп Корси, создавший его по образцу Хебба диапазон цифр задание путем замены числовых заданий на пространственные. В среднем большинство участников набирают пять пунктов в тесте на интервал Корси и семь - в задаче на интервал цифр.
Диапазон визуальных паттернов
Это похоже на тест постукивания по блоку Корси, но рассматривается как более чистый тест на визуальное краткосрочное вспоминание.[19] Участникам предлагается серия шаблонов матриц, половина ячеек которых окрашена, а другая половина - пустая. Матричные шаблоны организованы таким образом, что их трудно кодировать вербально, что заставляет участника полагаться на визуальную пространственную память. Начиная с небольшой матрицы 2 x 2, участники копируют шаблон матрицы из памяти в пустую матрицу. Матричные паттерны увеличиваются в размере и сложности со скоростью две ячейки, пока способность участника воспроизводить их не ломается. В среднем производительность участников снижается в шестнадцати ячейках.
Задача диапазона пути
Это задание предназначено для измерения способностей пространственной памяти у детей.[17] Экспериментатор просит участника визуализировать пустую матрицу с человечком. Посредством серии инструкций, таких как вперед, назад, влево или вправо, экспериментатор направляет маленького человечка участника по пути через матрицу. В конце участника просят указать на реальной матрице, где закончился маленький человечек, которого он или она визуализировал. Длина пути варьируется в зависимости от уровня сложности (1-10), а сами матрицы могут иметь длину от 2 x 2 ячеек до 6 x 6.
Динамические лабиринты
Они предназначены для измерения пространственных способностей у детей. С помощью этого теста экспериментатор представляет участнику рисунок лабиринта с изображением человека в центре.[17] Пока участник наблюдает, экспериментатор использует свой палец, чтобы проследить путь от входа в лабиринт до рисунка человека. Затем ожидается, что участник воспроизведет продемонстрированный путь через лабиринт к рисунку человека. По мере увеличения сложности лабиринты различаются по сложности.
Лабиринт с радиальным рукавом

Впервые применена Олтоном и Самуэльсоном в 1976 году.[20] Лабиринт с радиальным плечом предназначен для проверки способности крыс к пространственной памяти. Лабиринты обычно имеют центральную платформу и различное количество рук.[21] ответвляются с пищей, размещенной на концах. Руки обычно каким-то образом экранированы друг от друга, но не до такой степени, чтобы внешние сигналы нельзя было использовать в качестве ориентира.
В большинстве случаев крысу помещают в центр лабиринта, и ей нужно исследовать каждую руку индивидуально, чтобы достать еду, одновременно запоминая, какие руки она уже преследовала. Лабиринт устроен так, что крыса вынуждена вернуться в центр лабиринта, прежде чем преследовать другую руку. Обычно принимаются меры, чтобы крыса не использовала свой обонятельный чувства к перемещаться например, размещение дополнительной еды на дне лабиринта.
Задача Морриса по навигации по воде - классический тест на изучение пространственного обучения и памяти у крыс.[22] и был впервые разработан в 1981 году Ричардом Г. Моррисом, в честь которого назван тест. Объект помещается в круглый резервуар с прозрачной водой со стенками, которые слишком высоки, чтобы он мог вылезти, и водой, которая слишком глубока, чтобы он мог стоять. Кроме того, стены резервуара украшены визуальными подсказками, которые могут служить ориентиром. точки. Крыса должна плавать вокруг бассейна, пока случайно не обнаружит прямо под поверхностью скрытую платформу, на которую она может забраться.
Как правило, крысы сначала плавают по краю бассейна, а затем, извилистым узором, выходят в центр, прежде чем наткнуться на скрытую платформу. Однако по мере того, как время, проведенное в бассейне, увеличивает опыт, количество времени, необходимое для определения местоположения платформы, уменьшается, и крысы-ветераны подплывают прямо к платформе почти сразу после того, как их поместили в воду.
Физиология
Гиппокамп
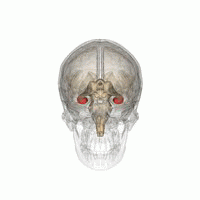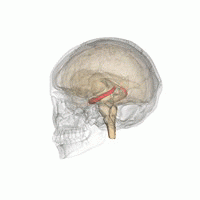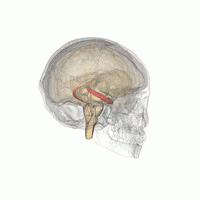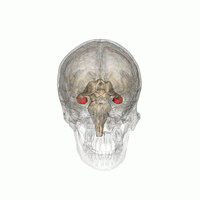
В гиппокамп предоставляет животным пространственную карту их среды обитания.[23] Он хранит информацию, касающуюся неэгоцентрического пространства (эгоцентрические средства в отношении положения тела в пространстве) и, следовательно, поддерживает независимость точки зрения в пространственной памяти.[24] Это означает, что он позволяет управлять точкой обзора из памяти. Однако это важно для долговременной пространственной памяти аллоцентрического пространства (ссылки на внешние сигналы в пространстве).[25] Таким образом, поддержание и извлечение воспоминаний являются относительными или зависит от контекста.[26] Гиппокамп использует справочную и рабочую память и играет важную роль в обработке информации о пространственном местоположении.[27]
Блокировка пластичность в этой области приводит к проблемам с целенаправленной навигацией и ухудшает способность запоминать точное местоположение.[28] Пациенты с амнезией и повреждением гиппокампа не могут изучить или запомнить пространственное расположение, а пациенты, перенесшие удаление гиппокампа, серьезно нарушены в пространственной навигации.[24][29] Обезьяны с поражениями в этой области не могут научиться ассоциациям объект-место, а крысы также демонстрируют пространственный дефицит, не реагируя на пространственные изменения.[24][30] Кроме того, было показано, что крысы с поражениями гиппокампа временно не классифицируются (не зависят от времени). ретроградная амнезия которая устойчива к распознаванию выученной платформенной задачи только тогда, когда поражен весь гиппокамп, но не когда он частично поврежден.[31] Дефицит пространственной памяти также обнаруживается в задачах пространственной дискриминации.[29]

Большие различия в пространственных нарушениях обнаруживаются среди спинной и вентральный гиппокамп. Поражения вентрального гиппокампа не влияют на пространственную память, в то время как дорсальный гиппокамп необходим для восстановления, обработки кратковременной памяти и перевода памяти из краткосрочной в более длительные периоды задержки.[32][33][34] Настой амфетамин в спинной гиппокамп также улучшает память о пространственных положениях, изученных ранее.[35] Эти данные указывают на то, что существует функциональная диссоциация между дорсальным и вентральным гиппокампом.
Также наблюдаются полушарные различия внутри гиппокампа. Исследование на Лондон таксистов просили вспомнить сложные маршруты по городу, а также известные ориентиры для которых водители не знали своего пространственного положения. Это привело к активации правого гиппокампа только во время вызова сложных маршрутов, что указывает на то, что правый гиппокамп используется для навигации в крупномасштабной пространственной среде.[36]
Известно, что гиппокамп содержит две отдельные цепи памяти. Одна схема используется для памяти распознавания мест на основе воспоминаний и включает система энторинал-СА1,[37] в то время как другая система, состоящая из гиппокампа трисинаптическая петля (entohinal-dentate-CA3-CA1) используется для запоминания места[38] и облегчения пластичности энторинально-зубчатого синапса у мышей достаточно для улучшения запоминания места.[39]
Поместите ячейки также находятся в гиппокампе.
Задняя теменная кора
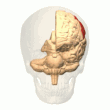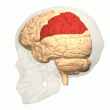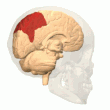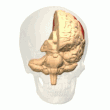
В теменная кора кодирует пространственную информацию, используя эгоцентрическую систему координат. Следовательно, он участвует в преобразовании координат сенсорной информации в координаты действия или эффектора путем обновления пространственного представления тела в окружающей среде.[40] В результате поражения теменной коры вызывают дефицит в овладении и удержании эгоцентрических задач, тогда как среди аллоцентрических задач наблюдаются незначительные нарушения.[41]
Крысы с поражением передний регион задняя теменная кора повторно исследовать перемещенные объекты, в то время как крысы с повреждениями задний область задней теменной коры не реагировала на пространственные изменения.[30]
Известно также, что поражения теменной коры вызывают временно неклассифицированные ретроградная амнезия.[42]
Энторинальная кора

Дорсалькаудальный медиальный энторинальная кора (dMEC) содержит топографически организованную карту пространственной среды, состоящую из ячейки сетки.[43] Таким образом, эта область мозга преобразует сенсорный ввод из окружающей среды и сохраняет его в виде прочного аллоцентрического представления в мозге, которое будет использоваться для интеграция пути.[44]
Энторинальная кора способствует обработке и интеграции геометрических свойств и информации в окружающей среде.[45] Поражения в этой области затрудняют использование дистальный но нет проксимальный ориентиры во время навигации и вызывает зависящий от задержки дефицит пространственной памяти, который пропорционален длине задержки.[46][47] Известно также, что поражения в этой области создают дефицит удержания при выполнении задач, которым вы научились за 4 недели, но не за 6 недель до повреждений.[42]
Консолидация памяти в энторинальной коре достигается за счет регулируемых внеклеточными сигналами киназа Мероприятия.[48]
Префронтальная кора

Медиальный префронтальная кора обрабатывает эгоцентрическую пространственную информацию. Он участвует в обработке кратковременной пространственной памяти, используемой для управления запланированным поисковым поведением, и, как полагают, объединяет пространственную информацию с ее мотивационный значимость.[34][49] Идентификация нейронов, ожидающих ожидаемого награды в пространственной задаче поддерживают эту гипотезу. Медиальная префронтальная кора также участвует во временной организации информации.[50]
Специализация полушария находится в этой области мозга. Левая префронтальная кора преимущественно обрабатывает категориальную пространственную память, включая исходную память (ссылку на пространственные отношения между местом или событием), тогда как правая префронтальная кора преимущественно обрабатывает координатную пространственную память, включая память элемента (ссылка на пространственные отношения между характеристиками элемента).[51]
Поражения медиальной префронтальной коры ухудшают производительность крыс в ранее обученном лабиринте с лучевыми руками, однако крысы могут постепенно улучшаться до уровня контроля в зависимости от опыта.[52] Поражения в этой области также вызывают дефицит отложенных несоответствий по позициям и нарушения в усвоении задач пространственной памяти во время тренировочных проб.[53][54]
Ретроспленальная кора
В ретроспленальная кора участвует в обработке аллоцентрической памяти и геометрические свойства в окружающей среде.[45] Инактивация этой области приводит к нарушению навигации в темноте и, таким образом, подразумевает участие в процессе интеграция пути.[55]
Поражения ретроспленальной коры постоянно ухудшают тесты аллоцентрической памяти, сохраняя при этом эгоцентрическую память.[56] Животные с поражениями каудальной ретроспленальной коры демонстрируют снижение производительности в лабиринте с лучевым плечом только тогда, когда лабиринт поворачивают, чтобы не полагаться на сигналы внутри лабиринта.[57]

У людей повреждение ретроспленальной коры приводит к топографической дезориентации. Большинство случаев связано с повреждением правой ретроспленальной коры и включает область 30 Бродмана. Пациенты часто теряют способность изучать новые маршруты и перемещаться по знакомой среде.[58] Однако большинство пациентов обычно выздоравливают в течение 8 недель.
Ретроспленальная кора преимущественно обрабатывает пространственную информацию в правом полушарии.[58]
Периринальная кора
В периринальная кора связан как с пространственной привязкой, так и с пространственной рабочей памятью.[27] Он обрабатывает относительную информацию об экологических подсказках и местах.
Поражения в перигинальной коре головного мозга вызывают дефицит справочной памяти и рабочей памяти и увеличивают скорость забывая информации во время тренировочных испытаний водного лабиринта Морриса.[59] Это объясняет обесценение при первоначальном приобретении задачи. Поражения также вызывают нарушение задачи определения местоположения объекта и снижают привыкание к новой среде.[27]
Нейропластичность
Пространственные воспоминания формируются после того, как животное собирает и обрабатывает сенсорную информацию о своем окружении (особенно зрение и проприоцепция ). В общем, млекопитающим требуется функционирующий гиппокамп (особенно область CA1) для формирования и обработки воспоминаний о космосе. Есть некоторые свидетельства того, что пространственная память человека сильно связана с правым полушарием мозга.[60][61][62]
Пространственное обучение требует обоих NMDA и AMPA рецепторов, консолидация требует рецепторов NMDA, а восстановление пространственной памяти требует рецепторов AMPA.[63] Было показано, что у грызунов пространственная память зависит от размера части гиппокампа. моховое волокно проекция.[64]
Функция рецепторов NMDA варьируется в зависимости от субрегиона гиппокампа. Рецепторы NMDA необходимы в CA3 гиппокампа, когда необходимо реорганизовать пространственную информацию, в то время как рецепторы NMDA в CA1 требуются для получения и извлечения памяти после задержки, а также для формирования полей мест CA1.[65] Блокада рецепторов NMDA предотвращает индукцию долгосрочное потенцирование и ухудшает пространственное обучение.[66]
CA3 гиппокампа играет особенно важную роль в кодировании и восстановлении пространственных воспоминаний. CA3 иннервируется двумя афферентными путями, известными как перфорантный путь (PPCA3) и зубчатые извилины (DG) -опосредованные мшистые волокна (MF). Первый путь рассматривается как путь индекса поиска, а второй связан с кодированием.[67]
Расстройства / дефициты
Топографическая дезориентация
Топографическая дезориентация - это когнитивное расстройство, в результате которого человек не может ориентироваться в реальной или виртуальной среде. Пациенты также борются с задачами, зависящими от пространственной информации. Эти проблемы могут быть результатом нарушения способности доступа к своей когнитивной карте, мысленного представления окружающей среды или неспособности судить о местоположении объектов по отношению к самому себе.[68]
Топографическая дезориентация развития (DTD) диагностируется, когда пациенты показали неспособность перемещаться даже в знакомом с рождения окружении и не демонстрируют очевидных неврологических причин этого дефицита, таких как повреждение или повреждение мозга. DTD - относительно новое заболевание, которое может иметь разную степень тяжести.
Топографическая дезориентация при умеренном когнитивном нарушении: морфометрическое исследование на основе вокселей было проведено, чтобы выяснить, влияет ли топографическая дезориентация на людей с легкими когнитивными нарушениями. В исследование были включены сорок один пациент с диагнозом MCI и 24 здоровых человека из контрольной группы. Для этого эксперимента были установлены следующие стандарты:
- Субъективная когнитивная жалоба пациента или его / ее опекуна.
- Нормальная общая когнитивная функция выше 16-го процентиля по корейской версии Краткого экзамена на психическое состояние (K-MMSE).
- Нормальная повседневная деятельность (ADL) оценивалась как клинически, так и по стандартной шкале (как описано ниже).
- Объективное снижение когнитивных функций ниже 16-го процентиля по нейропсихологическим тестам.
- Исключение деменции.
(TD) оценивалась клинически у всех участников. Неврологические и нейропсихологические оценки проводились с помощью сканирования магнитной томографии, которое проводилось для каждого участника. Морфометрия на основе вокселей использовалась для сравнения паттернов атрофии серого вещества между пациентами с ТД и без нее, а также группой нормального контроля. Результатом эксперимента было то, что они обнаружили TD у 17 из 41 пациента с MCI (41,4%). Функциональные возможности были значительно нарушены у пациентов с MCI с TD по сравнению с пациентами с MCI без TD, и что наличие TD у пациентов с MCI связано с потерей серого вещества в медиальных височных областях, включая гиппокамп.[69]
Повреждение гиппокампа и шизофрения
Исследования на крысах показывают, что на пространственную память могут отрицательно влиять неонатальный повреждение гиппокампа способом, который очень напоминает шизофрения. Считается, что шизофрения происходит от неврологический проблемы вскоре после рождения.[70]
Крысы обычно используются в качестве моделей пациентов с шизофренией. Экспериментаторы создают поражения в вентральной области гиппокампа вскоре после рождения, эта процедура известна как неонатальное вентральное поражение гиппокампа (NVHL). Взрослые крысы с NVHL демонстрируют типичные признаки шизофрении, такие как гиперчувствительность к психостимуляторы, снижение социальных взаимодействий и нарушение предымпульсное торможение, рабочая память и набор-смещение.[71][72][73][74][75] Подобно шизофрении, крысы с ограниченными возможностями не могут использовать контекст окружающей среды в задачах пространственного обучения, таких как демонстрация сложности прохождения лабиринта с радиальными рукавами и водного лабиринта Мориса.[76][77][78]
GPS

Недавние исследования пространственной памяти и поиск пути в статье Исикавы и другие. в 2008[79] показал, что использование GPS Движущееся устройство карты снижает возможности навигации человека по сравнению с другими участниками, которые использовали карты или имели предыдущий опыт на маршруте с гидом. Устройства с движущейся картой GPS часто настраиваются так, чтобы пользователь мог видеть только небольшой подробный крупный план определенного сегмента карты, который постоянно обновляется. Для сравнения, карты обычно позволяют пользователю видеть один и тот же вид всего маршрута от отправления до прибытия. Другое исследование показало, что люди, использующие GPS, в целом путешествуют медленнее, чем пользователи карт, которые путешествуют быстрее. Пользователи GPS останавливаются чаще и на более длительный период времени, тогда как пользователи карт и отдельные лица, использующие прошлый опыт в качестве ориентира, путешествуют по более прямым маршрутам для достижения своей цели.
NEIL1
Эндонуклеаза VIII-подобная 1 (NEIL1 ) это Ремонт ДНК фермент, который широко экспрессируется в мозг. NEIL1 - это ДНК гликозилаза что инициирует первый шаг в базовая эксцизионная пластика путем расщепления оснований, поврежденных активными формами кислорода, а затем введения разрыва цепи ДНК через связанный лиазе реакция. Этот фермент распознает и удаляет окисленные основания ДНК включая формамидопиримидин, тимингликоль, 5-гидроксиурацил и 5-гидроксицитозин. NEIL1 способствует кратковременному сохранению пространственной памяти.[80] У мышей, лишенных NEIL1, в тесте в водном лабиринте нарушалась кратковременная пространственная память.[80]
Проблемы в изучении
Невербальная неспособность к обучению характеризуется нормальными речевыми способностями, но нарушенными зрительно-пространственными способностями. Проблемными областями для детей с невербальной обучаемостью являются арифметика, геометрия и естественные науки. Нарушения пространственной памяти связаны с нарушением невербального обучения и другими трудностями в обучении.[81]
Арифметика текстовые задачи включают письменный текст, содержащий набор данных, за которым следует один или несколько вопросов, и требуют использования четырех основных арифметических операций (сложение, вычитание, умножение или деление).[16] Исследователи предполагают, что успешное решение арифметических задач со словами требует пространственного рабочая память (участвует в построении схематических представлений), который облегчает создание пространственных отношений между объектами. Создание пространственных отношений между объектами - важная часть решения текстовых задач, поскольку требуются мысленные операции и преобразования.[16]
Например, рассмотрим следующий вопрос: «Ребенок строит три башни из блоков красного и белого цвета одинакового размера. В самой нижней башне 14 блоков, в самой высокой - на 7 блоков больше. В промежуточной башне на три блока меньше, чем в самой высокой. . Сколько блоков в каждой из трех башен? "[16] Чтобы решить вопрос, необходимо поддерживать поступающую информацию (например, текст) и интегрировать ее с предыдущей информацией (например, знаниями для арифметических операций). Человек также должен выбирать релевантную (то есть пространственные отношения между блоками) и запрещать нерелевантную информацию (то есть цвета и текстуры блоков) и одновременно создавать мысленное представление о проблеме.[16]
Исследователи изучали роль пространственной и зрительной памяти в способности решать арифметические задачи со словами. Дети в исследовании выполнили задание блока Корси (прямая и обратная серия) и задание пространственной матрицы, а также задание на визуальную память, называемое тестом на распознавание дома. Бедные решающие проблемы были нарушены в задачах блока Корси и задаче пространственной матрицы, но выполнялись нормально в тесте на распознавание дома по сравнению с нормально достигающими детьми. Эксперимент показал, что плохое решение проблемы связано именно с недостаточной обработкой пространственной информации.[16]
Спать
Спать было обнаружено, что улучшает пространственную память за счет усиления гиппокампа. консолидация памяти.[82] Области гиппокампа, активированные при обучении маршрутам, реактивируются во время последующего сна (NREM сон особенно). В конкретном исследовании было продемонстрировано, что фактическая степень реактивации во время сна коррелировала с улучшением поиска маршрута и, следовательно, улучшения памяти на следующий день.[83] Таким образом, исследование установило идею о том, что сон усиливает процесс консолидации на системном уровне, который, следовательно, улучшает / улучшает поведенческие характеристики. Кроме того, период бодрствования не влияет на стабилизацию следов памяти по сравнению с периодом сна. Сон после первой ночи после тренировки, то есть во вторую ночь, не способствует дальнейшей консолидации пространственной памяти. Следовательно, сон в первую ночь после тренировки, например после изучения маршрута самое важное.[82]
Связь лишения сна и сна также была исследована. Лишение сна фактически препятствует повышению производительности памяти из-за активного нарушения консолидации пространственной памяти.[82] Таким образом, пространственная память улучшается за счет периода сна.
Смотрите также
- Познавательная карта
- Диссоциация (нейропсихология)
- Метод локусов
- Пространственная способность
- Картирование космоса
- Зрительная память
Рекомендации
- ^ а б c d е Johnson, E .; Адамо-Виллани, Н. (2010). «Исследование влияния погружения на кратковременную пространственную память». Инженерия и технологии. 71: 582–587.
- ^ а б c d е ж грамм час я j k л Ang, S. Y .; Ли, К. (2008). «Центральное исполнительное участие в пространственной памяти детей». объем памяти. 16 (8): 918–933. Дои:10.1080/09658210802365347. PMID 18802804. S2CID 42610047.
- ^ а б c d е ж Jones, D .; Farrand, P .; Стюарт, G .; Morris, N .; и другие. (1995). «Функциональная эквивалентность вербальной и пространственной информации в серийной кратковременной памяти». Журнал экспериментальной психологии: обучение, память и познание. 21 (4): 1008–1018. Дои:10.1037/0278-7393.21.4.1008. PMID 7673864.
- ^ а б c d е Della Sala, S .; Серый, C .; Baddeley, A .; Allamano, N .; Wilson, L .; и другие. (1999). «Размах паттернов: инструмент для раскрытия зрительно-пространственной памяти». Нейропсихология. 37 (10): 1189–1199. Дои:10.1016 / S0028-3932 (98) 00159-6. PMID 10509840. S2CID 1894333.
- ^ а б c d е Mammarella, I.C .; Pazzaglia, F .; Cornoldi, C .; и другие. (2008). «Доказательства различных компонентов зрительно-пространственной рабочей памяти детей». Британский журнал психологии развития. 26 (3): 337–355. Дои:10.1348 / 026151007X236061.
- ^ а б c d е ж грамм час я j k л м п о п Калакоски, В .; Саарилуома, П. (2001). «Исключительная память водителей такси о названиях улиц». Память и познание. 29 (4): 634–638. Дои:10.3758 / BF03200464. PMID 11504011.
- ^ Fisk, J. E .; Sharp, C.A .; и другие. (2003). «Роль исполнительной системы в функционировании зрительно-пространственной памяти». Мозг и познание. 52 (3): 364–381. Дои:10.1016 / S0278-2626 (03) 00183-0. PMID 12907181. S2CID 37589101.
- ^ а б c d е Чун, М .; Цзян, Ю. (1998). «Контекстная подсказка: неявное обучение и память визуального контекста направляют пространственное внимание». Когнитивная психология. 36 (1): 28–71. CiteSeerX 10.1.1.25.5066. Дои:10.1006 / cogp.1998.0681. PMID 9679076. S2CID 1955059.
- ^ а б McNamara, T .; Харди, Дж .; Hirtle, S .; и другие. (1989). «Субъективные иерархии в пространственной памяти». Журнал экспериментальной психологии. 15 (2): 211–227. Дои:10.1037/0278-7393.15.2.211. PMID 2522511.
- ^ а б c Newman, E.L .; Caplan, J.B .; Киршен, М.П .; Королев И.О .; Секулер, Р .; Kahana, M.J .; и другие. (2007). «Узнай свой путь по городу: как водители виртуальных такси учатся использовать и макет, и информацию о достопримечательностях» (PDF). Познание. 104 (2): 231–253. CiteSeerX 10.1.1.69.5387. Дои:10.1016 / j.cognition.2006.05.013. PMID 16879816. S2CID 267034.
- ^ Гиллнер С, Маллот Х (2000). «Роль глобальных и локальных ориентиров в навигации по виртуальной среде». Восприятие. 9 (1): 69–83. CiteSeerX 10.1.1.138.8266. Дои:10.1162/105474600566628. S2CID 13062306.
- ^ Крокос, Эрик; Плезан, Екатерина; Варшней, Амитабх (16 мая 2018 г.). "Дворцы виртуальной памяти: воспоминания о помощи при погружении". Виртуальная реальность. 23: 1–15. Дои:10.1007 / s10055-018-0346-3.
- ^ а б c d е ж Bird, L.R .; Робертс, В. А .; Abroms, B .; Kit, K. A .; Крапи, К. (2003). «Пространственная память на пищу, скрытую крысами (Раттус норвегикус) по радиальному лабиринту: исследования памяти о том, где, что и когда ». Журнал сравнительной психологии. 117 (2): 176–187. Дои:10.1037/0735-7036.117.2.176. PMID 12856788.
- ^ Дженнифер С. Холланд (8 августа 2014 г.). "Смотрите: как далеко бродят ваши кошки?". Национальная география. Получено 23 августа, 2016.
- ^ а б c Klauer, K.C .; Zhao, Z .; и другие. (2004). «Двойные диссоциации в зрительной и пространственной кратковременной памяти». Журнал экспериментальной психологии: Общие. 133 (3): 355–381. Дои:10.1037/0096-3445.133.3.355. PMID 15355144. S2CID 7224613.
- ^ а б c d е ж грамм Passolunghi, M.C .; Mammarella, I.C .; и другие. (2010). «Способность к пространственной и зрительной рабочей памяти у детей с трудностями в решении арифметических задач». Европейский журнал когнитивной психологии. 22 (6): 944–963. Дои:10.1080/09541440903091127. S2CID 143882629.
- ^ а б c Mammarella, I.C .; Pazzaglia, F .; Корнольди, К. (2008). «Доказательства различных компонентов зрительно-пространственной рабочей памяти детей». Британский журнал психологии развития. 26 (3): 337–355. Дои:10.1348 / 026151007X236061.
- ^ Корси, П. М. (1972). «Память человека и медиальная височная область мозга». Международные тезисы диссертаций. 34 (2): 891.
- ^ Делла Сала, С., Грей, К., Баддели, А., и Уилсон, Л. (1997). Тест на визуальные шаблоны: новый тест на кратковременную визуальную память. Фелтхэм, Саффолк: испытательная компания в долине Темзы.
- ^ Olton, D.S .; Самуэльсон, Р.Дж. (1976). «Воспоминание о местах прошлого: пространственная память у крыс». Журнал экспериментальной психологии: процессы поведения животных. 2 (2): 97–116. CiteSeerX 10.1.1.456.3110. Дои:10.1037/0097-7403.2.2.97.
- ^ Cole, M.R .; Чаппелл-Стивенсон, Робин (2003). «Исследование пределов пространственной памяти с помощью очень больших лабиринтов». Обучение и поведение. 31 (4): 349–368. Дои:10.3758 / BF03195996. PMID 14733483.
- ^ Моррис, Р. Г. (1981). «Пространственная локализация не требует наличия локальных ориентиров». Обучение и мотивация. 12 (2): 239–260. Дои:10.1016/0023-9690(81)90020-5.
- ^ О'Киф, Дж .; Достровский, Дж. (1971). «Гиппокамп как пространственная карта. Предварительные данные о деятельности единиц у свободно перемещающейся крысы». Исследование мозга. 34 (1): 171–175. Дои:10.1016/0006-8993(71)90358-1. PMID 5124915.
- ^ а б c Сквайр, Л. Р. (1992). «Память и гиппокамп: синтез результатов, полученных на крысах, обезьянах и людях». Психологический обзор. 99 (2): 195–231. Дои:10.1037 / 0033-295X.99.2.195. PMID 1594723. S2CID 14104324.
- ^ Рамос, Дж. М. Дж. (2000). «Долговременная пространственная память у крыс с поражением гиппокампа». Европейский журнал нейробиологии. 12 (9): 3375–3384. Дои:10.1046 / j.1460-9568.2000.00206.x. PMID 10998120. S2CID 18121369.
- ^ Winocur, G .; Москович, М .; Каруана, Д. А .; Биннс, М.А. (2005). «Ретроградная амнезия у крыс с поражением гиппокампа при тестировании пространственной памяти». Нейропсихология. 43 (11): 1580–1590. Дои:10.1016 / j.neuropsychologia.2005.01.013. PMID 16009240. S2CID 9160452.
- ^ а б c Liu, P .; Билки, Д. К. (2001). «Эффект эксайтотоксических поражений, сосредоточенных на гиппокампе или периринальной коре в задачах распознавания объектов и пространственной памяти». Поведенческая неврология. 115 (1): 94–111. Дои:10.1037/0735-7044.115.1.94. PMID 11256456.
- ^ Hebert, A.E .; Даш, П. К. (2004). «Неизбыточные роли гиппокампа и энторинальной корковой пластичности в хранении пространственной памяти». Фармакология, биохимия и поведение. 79 (1): 143–153. Дои:10.1016 / j.pbb.2004.06.016. PMID 15388294. S2CID 25385758.
- ^ а б Сохранить, E .; Poucet, B .; Бригадир, Н .; Бухот, М. (1992). «Исследование объекта и реакции на пространственные и непространственные изменения у крыс с капюшонами после повреждения теменной коры или образования гиппокампа». Поведенческая неврология. 106 (3): 447–456. Дои:10.1037/0735-7044.106.3.447. PMID 1616611.
- ^ Мартин, С. Дж .; de Hozl, L .; Моррис, Р. Г. М. (2005). «Ретроградная амнезия: ни частичное, ни полное поражение гиппокампа у крыс не приводит к преимущественному сохранению отдаленной пространственной памяти даже после напоминания». Нейропсихология. 43 (4): 609–624. Дои:10.1016 / j.neuropsychologia.2004.07.007. PMID 15716151. S2CID 24894665.
- ^ Баннерман, Д. М .; Дикон, Р. М. Дж .; Offen, S .; Friswell, J .; Grubb, M .; Роулинз, Дж. Н. П. (2002). «Двойная диссоциация функций в гиппокампе: пространственная память и гипонеофагия». Поведенческая неврология. 116 (5): 884–901. Дои:10.1037/0735-7044.116.5.884. PMID 12369808.
- ^ Moser, M .; Мозер, Э. И. (1998). «Распределенное кодирование и извлечение пространственной памяти в гиппокампе». Журнал неврологии. 18 (18): 7535–7542. Дои:10.1523 / JNEUROSCI.18-18-07535.1998. ЧВК 6793256. PMID 9736671.
- ^ а б Lee, I .; Кеснер, Р. П. (2003). «Зависящие от времени отношения между дорсальным гиппокампом и префронтальной корой в пространственной памяти». Журнал неврологии. 23 (4): 1517–1523. Дои:10.1523 / JNEUROSCI.23-04-01517.2003. ЧВК 6742248. PMID 12598640.
- ^ Макгоу, Дж. Л. (2000). «Память - век консолидации». Наука. 287 (5451): 248–251. Bibcode:2000Sci ... 287..248M. Дои:10.1126 / science.287.5451.248. PMID 10634773. S2CID 40693856.
- ^ Maguire, E. A .; Frackowiak, R. S. J .; Фрит, К. Д. (1997). «Вспоминая маршруты по Лондону: активация правого гиппокампа у таксистов». Журнал неврологии. 17 (18): 7103–7110. Дои:10.1523 / JNEUROSCI.17-18-07103.1997. ЧВК 6573257. PMID 9278544.
- ^ Brun, V.H .; Otnaess, M. K .; Molden, S .; Steffenach, H .; Witter, M. P .; Moser, M .; Мозер, Э. И. (2002). «Размещение клеток и распознавание мест поддерживается прямой энторинально-гиппокампальной схемой». Наука. 296 (5576): 2243–2246. Bibcode:2002Sci ... 296.2243B. Дои:10.1126 / science.1071089. PMID 12077421. S2CID 8458253.
- ^ Goodrich-Hunsaker NJ, Hunsaker MR, Kesner RP (2008). «Взаимодействие и диссоциация субрегионов дорсального гиппокампа: как зубчатая извилина, CA3 и CA1 обрабатывают пространственную информацию». Behav. Неврологи. 122 (1): 16–26. Дои:10.1037/0735-7044.122.1.16. PMID 18298245.
- ^ Сааб Б.Дж., Георгиу Дж., Нат А., Ли Ф.Дж., Ван М., Михалон А., Лю Ф., Мансуй И.М., Родер Дж.С. (2009). «NCS-1 в зубчатой извилине способствует исследованию, синаптической пластичности и быстрому приобретению пространственной памяти». Нейрон. 63 (5): 643–56. Дои:10.1016 / j.neuron.2009.08.014. PMID 19755107. S2CID 5321020.
- ^ Colby, C.L .; Гольдберг, М. Э. (1999). «Пространство и внимание в теменной коре». Ежегодный обзор нейробиологии. 22: 319–349. Дои:10.1146 / annurev.neuro.22.1.319. PMID 10202542. S2CID 14379470.
- ^ Сохранить, E .; Могхаддам, М. (1996). «Влияние поражений ассоциативной теменной коры на приобретение и использование пространственной памяти в эгоцентрических и аллоцентрических навигационных задачах у крыс». Поведенческая неврология. 110 (1): 74–85. Дои:10.1037/0735-7044.110.1.74. PMID 8652075.
- ^ а б Cho, Y.H .; Кеснер, Р. П. (1996). «Участие энторинальной коры или теменной коры в долговременной пространственной дискриминирующей памяти у крыс: ретроградная амнезия». Поведенческая неврология. 110 (3): 436–442. Дои:10.1037/0735-7044.110.3.436. PMID 8888988.
- ^ Хафтинг, Т .; Fyhn, M .; Molden, S .; Moser, M .; Мозер, Э. И. (2005). «Микроструктура пространственной карты энторинальной коры». Природа. 436 (7052): 801–806. Bibcode:2005Натура 436..801H. Дои:10.1038 / природа03721. PMID 15965463. S2CID 4405184.
- ^ Fyhn, M .; Molden, S .; Witter, M. P .; Moser, E. I .; Мозер, М. (2004). «Пространственное представление в энторинальной коре». Наука. 305 (5688): 1258–1264. Bibcode:2004Наука ... 305.1258F. Дои:10.1126 / science.1099901. PMID 15333832.
- ^ а б Parron, C .; Сохранить, Э. (2004). «Сравнение эффектов энторинальных и ретроспленальных корковых поражений на привыкание, реакцию на пространственные и непространственные изменения во время исследования объекта у крысы». Нейробиология обучения и памяти. 82 (1): 1–11. Дои:10.1016 / j.nlm.2004.03.004. PMID 15183166. S2CID 36475737.
- ^ Parron, C .; Poucet, B .; Сохранить, Э. (2004). «Поражения энторинальной коры затрудняют использование дистальных, но не проксимальных ориентиров во время навигации места у крысы». Поведенческие исследования мозга. 154 (2): 345–352. Дои:10.1016 / j.bbr.2004.03.006. PMID 15313022. S2CID 919685.
- ^ Nagahara, H.A .; Отто, Т .; Галлахер, М. (1995). «Энторинально-периринальные поражения ухудшают производительность крыс по двум версиям обучения в водном лабиринте Морриса». Поведенческая неврология. 109 (1): 3–9. Дои:10.1037/0735-7044.109.1.3. PMID 7734077.
- ^ Hebert, A.E .; Даш, П. К. (2002). «Активность киназы, регулируемая внеклеточными сигналами, в энторинальной коре головного мозга необходима для долговременной пространственной памяти». Обучение и память. 9 (4): 156–166. Дои:10.1101 / лм. 48502. ЧВК 182586. PMID 12177229.
- ^ Pratt, W. E .; Мизумори, С.Дж.Ю. (2001). «Нейроны медиальной префронтальной коры головного мозга крысы показывают ожидаемые изменения скорости предсказуемой дифференциальной награды в задаче пространственной памяти». Поведенческие исследования мозга. 123 (2): 165–183. Дои:10.1016 / S0166-4328 (01) 00204-2. PMID 11399329. S2CID 3075976.
- ^ Kesner, R.P .; Холбрук, Т. (1987). «Диссоциация элемента и порядка пространственной памяти у крыс после поражения медиальной префронтальной коры». Нейропсихология. 25 (4): 653–664. Дои:10.1016 / 0028-3932 (87) 90056-X. PMID 3658148. S2CID 23596034.
- ^ Slotnick, S.D .; Му, Л. Р. (2006). «Специализация полушарий префронтальной коры на категориальную и координатную зрительную пространственную память». Нейропсихология. 44 (9): 1560–1568. Дои:10.1016 / j.neuropsychologia.2006.01.018. PMID 16516248. S2CID 14396813.
- ^ Becker, J. T .; Уокер, Дж. А .; Олтон, Д. С. (1980). «Нейроанатомические основы пространственной памяти». Исследование мозга. 200 (2): 307–320. Дои:10.1016/0006-8993(80)90922-1. PMID 7417818. S2CID 1429885.
- ^ Aggleton, J. P .; Neave, N .; Nagle, S .; Сахгал, А. (1995). «Сравнение влияния поражений медиальной префронтальной части, поясной коры и поясной связки на тесты пространственной памяти: доказательство двойной диссоциации между лобными и поясными связками». Журнал неврологии. 15 (11): 7270–7281. Дои:10.1523 / JNEUROSCI.15-11-07270.1995. ЧВК 6578066. PMID 7472481.
- ^ Lacroix, L .; Белый, I .; Фелдон, Дж. (2002). «Влияние эксайтотоксических поражений медиальной префронтальной коры головного мозга крыс на пространственную память». Поведенческие исследования мозга. 133 (1): 69–81. Дои:10.1016 / S0166-4328 (01) 00442-9. PMID 12048175. S2CID 24280276.
- ^ Cooper, B.G .; Manka, T. F .; Мизумори, С.Дж.Ю. (2001). «Ориентируясь в темноте: ретроспленальная кора головного мозга способствует пространственной памяти и навигации без визуальных подсказок». Поведенческая неврология. 115 (5): 1012–1028. Дои:10.1037/0735-7044.115.5.1012. PMID 11584914.
- ^ Vann, S.D .; Агглетон, Дж. П. (2002). «Обширные цитотоксические поражения ретроспленальной коры головного мозга крыс выявляют постоянный дефицит в задачах, требующих аллоцентрической пространственной памяти». Поведенческая неврология. 116 (1): 85–94. Дои:10.1037/0735-7044.116.1.85. PMID 11895186.
- ^ Vann, S.D .; Wilton, L.A .; Muir, J. L .; Агглетон, Дж. П. (2003). «Проверка важности каудальной ретроспленальной коры для пространственной памяти у крыс». Поведенческие исследования мозга. 140 (1–2): 107–118. Дои:10.1016 / S0166-4328 (02) 00274-7. PMID 12644284. S2CID 10400806.
- ^ а б Магуайр, Э. А. (2001). «Ретроспленальный вклад в навигацию человека: обзор поражений и результатов нейровизуализации». Скандинавский журнал психологии. 42 (3): 225–238. Дои:10.1111/1467-9450.00233. PMID 11501737.
- ^ Liu, P .; Билки, Д. К. (1998). "Вклад периринальной коры в работу в водном лабиринте Морриса". Поведенческая неврология. 112 (2): 304–315. Дои:10.1037/0735-7044.112.2.304. PMID 9588480.
- ^ Гутброд, К; Коэн, Р. Майер, Т; Мейер, Э (1987). «Память для пространственного и временного порядка у пациентов с афазией и повреждением правого полушария». Кора. 23 (3): 463–74. Дои:10.1016 / с0010-9452 (87) 80007-2. PMID 3677733. S2CID 4489682.
- ^ Nunn, JA; Graydon, FJ; Полки, CE; Моррис, Р.Г. (1999). «Дифференциальное ухудшение пространственной памяти после правой височной лобэктомии, продемонстрированное с помощью временного титрования». Мозг. 122 (1): 47–59. Дои:10.1093 / мозг / 122.1.47. PMID 10050894.
- ^ Такер, DM; Hartry-Speiser, A; McDougal, L; Луу, П; Дегранпре, Д. (1999). «Настроение и пространственная память: эмоции и вклад правого полушария в пространственное познание». Биол Психол. 50 (2): 103–25. Дои:10.1016 / S0301-0511 (99) 00005-8. PMID 10403200. S2CID 42669695.
- ^ Liang, KC; Hon, W; Тян Ю.М.; Ляо, WL (1994). «Участие гиппокампа рецепторов NMDA и AMPA в приобретении, формировании и восстановлении пространственной памяти в водном лабиринте Морриса». Chin J Physiol. 37 (4): 201–12. PMID 7796636.
- ^ Crusio, W. E .; Швеглер, Х. (2005). «Изучение задач пространственной ориентации в радиальном лабиринте и структурных вариаций в гиппокампе у инбредных мышей». Поведенческие и мозговые функции. 1 (1): 3. Дои:10.1186/1744-9081-1-3. ЧВК 1143776. PMID 15916698.
- ^ Lee, I .; Кеснер, Р. П. (2002). «Дифференциальный вклад рецепторов NMDA в субрегионах гиппокампа в пространственную рабочую память». Природа Неврология. 5 (2): 162–168. Дои:10.1038 / nn790. PMID 11780144. S2CID 17727921.
- ^ Моррис, Р. Г. М .; Андерсон, Э .; Lynch, G.S .; Бодри, М. (1986). «Избирательное нарушение обучения и блокада долгосрочного потенцирования анатагонистом рецептора N-метил-D-аспартата, AP5». Природа. 319 (6056): 774–776. Bibcode:1986Натура.319..774М. Дои:10.1038 / 319774a0. PMID 2869411. S2CID 4356601.
- ^ Lee, I .; Кеснер, Р. П. (2004). «Кодирование по сравнению с восстановлением пространственной памяти: двойная диссоциация между зубчатой извилиной и входами перфорантного пути в CA3 в дорсальном гиппокампе». Гиппокамп. 14 (1): 66–76. Дои:10.1002 / hipo.10167. PMID 15058484.
- ^ Старк, М; Coslett, HB; Saffran, EM (1996). Нарушение эгоцентрической карты локаций: последствия для восприятия и действий. 13. Cogn Neuropsychol. С. 481–523.
- ^ Тэ Сон Лим, Джузеппе Ярия, Со Юная Луна. «Топографическая дезориентация при легких когнитивных нарушениях: морфометрическое исследование на основе вокселей». 9 августа 2010 г. 16 апреля 2011 г. <http://www.neurolab.ca/2010(5)_Lim.pdf >.
- ^ Lewis, D.A .; Левитт, П. (2002). «Шизофрения как расстройство нервного развития». Ежегодный обзор нейробиологии. 25: 409–432. Дои:10.1146 / annurev.neuro.25.112701.142754. PMID 12052915.
- ^ Липска, Б.К .; Вайнбергер, Д. (2000). «Смоделировать психическое расстройство у животных: шизофрения как тест на реальность». Нейропсихофармакология. 23 (3): 223–239. Дои:10.1016 / S0893-133X (00) 00137-8. PMID 10942847.
- ^ Липска, Б.К .; Aultman, J.M .; Verma, A .; Weinberger, D.R .; Могхаддам Б. (2002). «Повреждение вентрального гиппокампа у новорожденных ухудшает рабочую память у крыс». Нейропсихофармакология. 27 (1): 47–54. Дои:10.1016 / S0893-133X (02) 00282-8. PMID 12062906.
- ^ Marquis, J. P .; Goulet, S .; Дор, Ф. Я. (2008). «Диссоциативное начало когнитивных и мотивационных дисфункций после неонатальных поражений вентрального гиппокампа у крыс». Поведенческая неврология. 122 (3): 629–642. Дои:10.1037/0735-7044.122.3.629. PMID 18513133.
- ^ Брэди, А. М. (2009). «Неонатальные вентральные поражения гиппокампа нарушают способность к переключению установки у взрослых крыс». Поведенческие исследования мозга. 205 (1): 294–298. Дои:10.1016 / j.bbr.2009.07.025. PMID 19646488. S2CID 29943421.
- ^ Marquis, J.P .; Goulet, S .; Доре, Ф. (2008). «Повреждения вентрального гиппокампа новорожденных нарушают экстрапространственный сдвиг и изменяют плотность дендритных шипов в медиальной префронтальной коре головного мозга молодых крыс». Нейробиология обучения и памяти. 90 (2): 339–346. Дои:10.1016 / j.nlm.2008.04.005. PMID 18490183. S2CID 26066133.
- ^ Винокур Г. и Миллс Дж. А. (1970). Передача между связанными и не связанными проблемами после поражения гиппокампа у крыс. Журнал сравнительной и физиологической психологии
- ^ Левин, Э.Д .; Кристофер, Северная Каролина (2006). «Влияние клозапина на функцию памяти на модели шизофрении неонатального гиппокампа у крыс». Прогресс нейропсихофармакологии и биологической психиатрии. 30 (2): 223–229. Дои:10.1016 / j.pnpbp.2005.10.018. PMID 16356617. S2CID 23594752.
- ^ Silva-Gomez, A.B .; Bermudez, M .; Quirion, R .; Srivastava, L.K .; Picazo, O .; Флорес, Г. (2003). «Сравнительные изменения в поведении самцов и самок постпубертатных крыс после неонатальных эксайтотоксических поражений вентрального гиппокампа». Исследование мозга. 973 (2): 285–292. Дои:10.1016 / S0006-8993 (03) 02537-X. PMID 12738072. S2CID 17456907.
- ^ Ishikawa, T .; Hiromichi, F .; Усама, I .; Ацуюки, О. (2008). «Ориентация с помощью мобильной навигационной системы на основе GPS: сравнение с картами и непосредственный опыт». Журнал экологической психологии. 28: 74–82. Дои:10.1016 / j.jenvp.2007.09.002.
- ^ а б Канугови К., Юн Дж.С., Фельдман Н.Х., Крото Д.Л., Маттсон М.П., Бор В.А. (сентябрь 2012 г.). «Эндонуклеаза VIII-подобная 1 (NEIL1) способствует кратковременному сохранению пространственной памяти и защищает от дисфункции мозга, вызванной ишемическим инсультом, и смерти мышей». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 109 (37): 14948–53. Bibcode:2012PNAS..10914948C. Дои:10.1073 / pnas.1204156109. ЧВК 3443144. PMID 22927410.
- ^ Mammarella, I.C .; Lucangeli, D .; Корнольди, К. (2010). «Пространственная рабочая память и арифметические нарушения у детей с невербальными трудностями в обучении». Журнал нарушений обучаемости. 43 (5): 455–468. Дои:10.1177/0022219409355482. PMID 20375290. S2CID 2596429.
- ^ а б c Феррара М., Ярия, Дж., Темпеста Д., Курсио Дж., Морони Ф., Марцано С., Де Дженнаро Л., Пачитти С. (2008). «Сон, чтобы найти свой путь: роль сна в укреплении памяти для навигации у людей». Гиппокамп. 18 (8): 844–851. Дои:10.1002 / hipo.20444. PMID 18493970.
- ^ Peigneux, P .; Laureys, S .; Fuchs, S .; Collette, F .; Perrin, F .; Reggers, J .; и другие. (2004). «Усиливаются ли пространственные воспоминания в гиппокампе человека во время медленного сна?». Нейрон. 44 (3): 535–545. Дои:10.1016 / j.neuron.2004.10.007. PMID 15504332. S2CID 1424898.