Корневой узелок - Root nodule

Корневые узелки находятся на корни из растения, в первую очередь бобовые, которые образуют симбиоз с азотфиксирующий бактерии.[1] Под азот -ограничивающие условия, способные растения формируют симбиотические отношения со специфическим для хозяина штаммом бактерий, известным как ризобия.[2] Этот процесс многократно развивался в бобовых, а также у других видов, встречающихся в Росид клады.[3] Бобовые культуры включают бобы, горох, и соевые бобы.
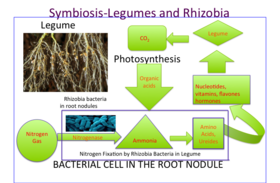
Внутри корневых клубеньков бобовых азот (N2) из атмосферы преобразуется в аммиак (NH3), который затем ассимилируется в аминокислоты (строительные блоки белков), нуклеотиды (строительные блоки ДНК и РНК а также важная энергетическая молекула АТФ ) и другие клеточные компоненты, такие как витамины, флавоны, и гормоны. Их способность исправить газообразный азот делает бобовые идеальным сельскохозяйственным организмом, поскольку их потребность в азотных удобрениях снижается. Действительно, высокое содержание азота блокирует развитие клубеньков, поскольку для растения нет пользы от формирования симбиоза. Энергия для расщепления газообразного азота в клубеньках поступает от сахара, который перемещается из листа (продукт фотосинтез ). Малат как продукт распада сахарозы является прямым источником углерода для бактериоида. Фиксация азота в узелке очень чувствительна к кислороду. В клубеньках бобовых содержится железосодержащий белок, называемый леггемоглобин, близкий к животному миоглобин, чтобы облегчить диффузию газообразного кислорода, используемого при дыхании.
Симбиоз
Бобовое семейство
Растения, способствующие азотфиксации, включают бобовые семья - Fabaceae - с такими таксонами, как кудзу, клевер, соевые бобы, люцерна, люпин, арахис, и Ройбуш. Они содержат симбиотический бактерии под названием ризобия внутри клубеньков, производя азотные соединения, которые помогают растению расти и конкурировать с другими растениями. Когда растение умирает, фиксированный азот высвобождается, делая его доступным для других растений, что помогает удобрять почва.[4][5] Подавляющее большинство бобовых культур имеют эту ассоциацию, но несколько родов (например, Стифнолобий ) не. Во многих традиционных методах земледелия поля чередуются с выращиванием различных культур, которые обычно включают в себя растение, состоящее в основном или полностью из клевера, чтобы воспользоваться этим.
Не зернобобовые
Хотя на сегодняшний день большинство растений, способных образовывать азотфиксирующие корневые клубеньки, относятся к семейству бобовых. Fabaceae, есть несколько исключений:
- Параспония, тропический род в Cannabaceae также способен взаимодействовать с ризобиями и образовывать узелки, фиксирующие азот[6]
- Актиноризальные растения такие как ольха и байберри также могут образовывать узелки, фиксирующие азот, благодаря симбиотической ассоциации с Frankia бактерии. Эти растения относятся к 25 родам[7] распределен среди 8 семейств растений.
Способность фиксировать азот у этих семейств присутствует далеко не повсеместно. Например, из 122 родов в Розоцветные, всего 4 роды способны фиксировать азот. Все эти семьи принадлежат к заказы Тыквенные, Fagales, и Росалес, который вместе с Fabales сформировать кладу евросиды. В этой кладе Fabales были первой ветвью ветви; таким образом, способность фиксировать азот может быть плезиоморфный и впоследствии утерян у большинства потомков исходного азотфиксирующего растения; однако может оказаться, что основной генетический и физиологический требования присутствовали в начальной стадии в последние общие предки всех этих растений, но только у некоторых из них полноценно функционируют:
| Семья: Genera Betulaceae: Альнус (ольха) | ...... | ...... | ...... | ...... |
|
Классификация

Описаны два основных типа узелков: детерминантные и неопределенные.[8]
Определенные узелки встречаются на некоторых племенах тропических бобовых, например, на племенах родов Глицин (соя), Phaseolus (фасоль обыкновенная) и Vigna. и на некоторых бобовых культурах умеренного климата, таких как Лотос. Эти детерминированные клубеньки теряют меристематическую активность вскоре после инициации, таким образом, рост происходит из-за размножения клеток, что приводит к образованию зрелых клубеньков сферической формы. Другие типы детерминированных клубеньков встречаются в самых разных травах, кустарниках и деревьях, таких как Арахис (арахис ). Они всегда связаны с пазухами боковых или придаточных корней и образуются после заражения через трещины, в которых эти корни выходят, и не используются. корневые волоски. Их внутреннее устройство сильно отличается от такового у соя тип узелка.[9]
Неопределенные узелки присутствуют в большинстве бобовых из всех трех подсемейств, будь то в регионах с умеренным климатом или в тропиках. Их можно увидеть в Faboideae бобовые, такие как Pisum (горох), Medicago (люцерна), Trifolium (клевер) и Vicia (вика) и все мимозоид бобовые, такие как акация s, несколько узловатых цезальпиниоид бобовые, такие как куропатка гороховая. Они получили название «индетерминантные», потому что поддерживают активную апикальную меристема который производит новые клетки для роста в течение жизни узелка. Это приводит к тому, что узелок имеет в целом цилиндрическую форму, которая может быть сильно разветвленной.[9] Поскольку они активно растут, неопределенные клубеньки обнаруживают зоны, которые разграничивают различные стадии развития / симбиоза:[10][11][12]

- Зона I - активная меристема. Здесь формируется новая ткань узелка, которая позже дифференцируется на другие зоны узелка.
- Зона II - зона заражения. Эта зона пронизана инфекционными нитями, полными бактерий. Клетки растений больше, чем в предыдущей зоне, и деление клеток прекращено.
- Интерзона II – III. Здесь бактерии проникли в клетки растений, которые содержат амилопласты. Они удлиняются и начинают окончательно дифференцироваться в симбиотические, азотфиксирующие бактероиды.
- Зона III - зона азотфиксации. Каждая ячейка в этой зоне содержит большой центральный вакуоль а цитоплазма заполнена полностью дифференцированными бактероидами, которые активно фиксация азота. Растение обеспечивает эти клетки леггемоглобин, в результате чего получается отчетливый розовый цвет.
- Зона IV - зона старения. Здесь происходит разложение растительных клеток и их бактериоидного содержимого. Распад гемового компонента леггемоглобина приводит к заметному зеленению у основания узелка.
Это наиболее широко изученный тип клубеньков, но детали сильно различаются в клубеньках арахиса и его родственников, а также некоторых других важных сельскохозяйственных культур, таких как люпин, где узелок образуется в результате прямого инфицирования ризобий через эпидермис и где инфекционные нити никогда не образуются. . Узелки разрастаются вокруг корня, образуя воротниковую структуру. В этих клубеньках и в клубеньках арахиса центральная инфицированная ткань однородна, без неинфицированных узлов, наблюдаемых в клубеньках сои и многих неопределенных типов, таких как горох и клевер.
Нодуляция

Выпуск бобовых органические соединения так как вторичные метаболиты называется флавоноиды от их корней, которые привлекают к себе ризобии и которые также активируют кивок гены в бактериях производить кивковые факторы и инициировать образование узелков.[13][14] Эти кивок факторы инициируют завивка корневых волос. Завивка начинается с того, что самый кончик корневых волосков завивается вокруг Ризобий. Внутри кончика корня образуется небольшая трубка, называемая инфекционной нитью, которая обеспечивает путь для Ризобий проникать в корневые клетки эпидермиса по мере того, как корневой волос продолжает скручиваться.[15]
Частичной завивки можно добиться даже кивок только фактор.[14] Это было продемонстрировано изолированием кивок факторы и их применение к частям корневых волос. Корневые волоски завивались в направлении нанесения, демонстрируя действие корневых волосков, пытающихся завиться вокруг бактерии. Даже нанесение на боковые корни вызывало скручивание. Это продемонстрировало, что это кивок сам фактор, а не бактерия, вызывающая стимуляцию завивки.[14]
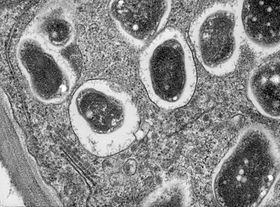
Когда нод-фактор ощущается корнем, происходит ряд биохимических и морфологических изменений: деление клеток запускается в корне, чтобы создать узелок, и корни волос рост перенаправляется, чтобы несколько раз обвиться вокруг бактерий, пока они полностью не охватят одну или несколько бактерий. Инкапсулированные бактерии многократно делятся, образуя микроколония. Из этой микроколонии бактерии попадают в развивающийся узелок через инфекционную нить, которая прорастает через корневые волоски в базальную часть эпидермис ячейку, а затем в корневая кора; затем они окружены растительным симбиосомная мембрана и дифференцируются на бактероиды, которые исправить азот.[16]
Эффективное клубенькование происходит примерно через четыре недели после посадки сельскохозяйственных культур размер и форма узелков зависят от урожая. Такие культуры, как соя или арахис, будут иметь более крупные клубеньки, чем кормовые бобовые, такие как красный клевер или люцерна, поскольку их потребности в азоте выше. Количество клубеньков и их внутренний цвет будут указывать на состояние азотфиксации в растении.[17]
Нодуляция контролируется множеством процессов, как внешних (тепло, кислые почвы, засуха, нитраты), так и внутренних (саморегуляция клубеньков, этилен). Ауторегуляция клубеньков[18] контролирует количество клубеньков на растении посредством системного процесса с участием листа. Ткань листа улавливает ранние события клубеньков в корне с помощью неизвестного химического сигнала, а затем ограничивает дальнейшее развитие клубеньков во вновь развивающихся тканях корня. Киназы рецепторов с высоким содержанием лейцина (LRR) (NARK в сое (Глицин макс); HAR1 в Лотос японский, СОЛНЦЕ через Medicago truncatula ) необходимы для ауторегуляции клубеньков (AON). Мутация, ведущая к потере функции этих киназ рецепторов AON, приводит к суперклубенькам или гиперклубенькам. Часто аномалии роста корней сопровождают потерю активности киназы рецептора AON, предполагая, что рост клубеньков и развитие корня функционально связаны. Исследования механизмов образования узелков показали, что ENOD40 ген, кодирующий белок из 12–13 аминокислот [41], активируется во время образования клубеньков [3].
Подключение к корневой структуре
Корневые узелки, по-видимому, трижды эволюционировали в пределах Fabaceae но редки вне этой семьи. Склонность этих растений к образованию корневых клубеньков, по-видимому, связана с их корневой структурой. В частности, склонность к развитию боковых корней в ответ на абсцизовая кислота может способствовать более позднему развитию корневых узелков.[19]
У других видов


Корневые клубеньки, встречающиеся на не бобовых родах, таких как Параспония в ассоциации с бактериями Rhizobium, а также те, которые возникают в результате симбиотических взаимодействий с Актинобактерии Frankia в некоторых родах растений, таких как Альнус, значительно отличаются от образовавшихся в симбиозе бобово-ризобийные. В этих симбиозах бактерии никогда не выходят из инфекционной нити. Frankia нодулирует около двухсот видов в следующих порядках (в скобках указаны семейства): Тыквенные (Coriariaceae и Datiscaceae ), Fagales (Betulaceae, Casuarinaceae, и Myricaceae ), Росалес (Рамновые, Elaeagnaceae и Розоцветные ).[20] На актиноризные симбиозы приходится примерно такое же количество азотфиксации, что и на ризобиальные симбиозы.[20] Все эти отряды вместе с Fabales образуют единую азотфиксирующую кладу внутри более широкой клады Росиды.
Немного грибы производить узловые структуры, известные как туберкулезные эктомикориза на корнях растений-хозяев. Suillus tomentosus, например, производит эти структуры с растением-хозяином Ложная сосна (Pinus contorta var. широколистная). Эти структуры, в свою очередь, могут принимать фиксация азота бактерии которые вносят значительный вклад азот и позволить соснам заселять бедные питательными веществами участки.[21]
Смотрите также
использованная литература
- ^ Вагнер СК (2011). «Биологическая азотфиксация». Знание природы образования. 3 (10): 15.
- ^ Ван, Ци и Ян, Шэнмин (2017). «Секретируемый хозяином антимикробный пептид усиливает симбиотическую селективность у Medicago truncatula». PNAS. 114 (26): 6854–6859. Дои:10.1073 / pnas.1700715114. ЧВК 5495241. PMID 28607058.
- ^ Дойл, Дж. Дж. И Луков, М. А. (2003). «Остальная часть айсберга. Разнообразие и эволюция бобовых в филогенетическом контексте». Физиология растений. 131 (3): 900–910. Дои:10.1104 / стр.102.018150. ЧВК 1540290. PMID 12644643.
- ^ Постгейт, Дж. (1998). Азотная фиксация, 3-е издание. Издательство Кембриджского университета, Кембридж, Великобритания.
- ^ Смил, В (2000). Циклы жизни. Научная американская библиотека.
- ^ Оп ден Камп, Рик; Streng, A .; и другие. (2010). "Микоризный рецептор LysM-типа, задействованный для симбиоза ризобий у животных, не являющихся бобовыми" Параспония". Наука. 331 (6019): 909–912. Bibcode:2011Sci ... 331..909O. Дои:10.1126 / science.1198181. PMID 21205637. S2CID 20501765.
- ^ Доусон, Дж. О. (2008). «Экология актиноризных растений». Актиноризный симбиоз, связывающий азот. Фиксация азота: происхождение, применение и результаты исследований. 6. Springer. С. 199–234. Дои:10.1007/978-1-4020-3547-0_8. ISBN 978-1-4020-3540-1. S2CID 85913801.
- ^ Мартин Креспи и Сусана Гальвес (2000). «Молекулярные механизмы в развитии корневых клубеньков». Журнал роста и регулирования растений. 19 (2): 155–166. Дои:10.1007 / s003440000023. PMID 11038225. S2CID 22216527.
- ^ а б Sprent 2009, Клубеньки бобовых: глобальная перспектива. Wiley-Blackwell
- ^ Фабрис Фуше и Ева Кондороси (2000). «Регуляция клеточного цикла в процессе органогенеза клубеньков в Medicago". Молекулярная биология растений. 43 (5–6): 773–786. Дои:10.1023 / А: 1006405029600. PMID 11089876. S2CID 11658948.
- ^ Ханна Монахан-Джованелли; Каталина Аранго Пинедо и Дэниел Дж. Гейдж (2006). «Архитектура сетей инфекционных потоков в развитии корневых узелков, индуцированных симбиотическими бактериями. Sinorhizobium meliloti на Medicago truncatula". Физиология растений. 140 (2): 661–670. Дои:10.1104 / стр. 105.072876. ЧВК 1361332. PMID 16384905.
- ^ Виллем Ван де Вельде; Хуан Карлос Перес Герра; Анник Де Кейзер; Рит Де Райке; и другие. (2006). «Старение в симбиозе бобовых. Молекулярный взгляд на старение клубеньков в Medicago truncatula". Физиология растений. 141 (2): 711–20. Дои:10.1104 / стр. 106.078691. ЧВК 1475454. PMID 16648219.
- ^ Эккардт Н.А. (июнь 2006 г.). «Роль флавоноидов в развитии корневых клубеньков и транспорте ауксина в Medicago truncatula». Растительная клетка. 18 (7): 1539–1540. Дои:10.1105 / tpc.106.044768. ЧВК 1488913.
- ^ а б c Эсселинг Дж. Дж., Люсье Ф. Г., Эмонс А. М. (август 2003 г.). «Завивание корневых волос, вызванное Nod-фактором: непрерывный полярный рост к точке применения Nod-фактора». Физиология растений. 132 (4): 1982–8. Дои:10.1104 / стр.103.021634. ЧВК 181283. PMID 12913154.
- ^ Слончевски Дж., Фостер Дж. В. (2017). Микробиология: развивающаяся наука (Четвертое изд.). Нью-Йорк. ISBN 978-0393614039. OCLC 951925510.
- ^ Mergaert, P .; Uchiumi, T .; Alunni, B .; Evanno, G .; Cheron, A .; Catrice, O .; и другие. (2006). «Эукариотический контроль бактериального клеточного цикла и дифференциации в симбиозе ризобий и бобовых». PNAS. 103 (13): 5230–35. Bibcode:2006ПНАС..103.5230М. Дои:10.1073 / pnas.0600912103. ISSN 1091-6490. ЧВК 1458823. PMID 16547129. Онлайн.
- ^ Аджей, М. «Азотфиксация и инокуляция кормовых бобовых культур» (PDF). Университет Флориды. Архивировано из оригинал (PDF) 2 декабря 2016 г.. Получено 1 декабря, 2016.
- ^ Reid, DE; Фергюсон, Б.Дж.; Хаяси, S; Линь, YH; Грессхофф, PM (октябрь 2011 г.). «Молекулярные механизмы, контролирующие ауторегуляцию клубеньков бобовых». Анналы ботаники. 108 (5): 789–95. Дои:10.1093 / aob / mcr205. ЧВК 3177682. PMID 21856632.
- ^ Ян Лян и Жанна М. Харрис (2005). «Реакция ветвления корней на абсцизовую кислоту коррелирует с образованием клубеньков как у бобовых, так и у небобовых». Американский журнал ботаники. 92 (10): 1675–1683. Дои:10.3732 / ajb.92.10.1675. PMID 21646084.
- ^ а б Джефф Дж. Дойл (1998). «Филогенетические взгляды на клубенькование: эволюционирующие взгляды на растения и симбиотические бактерии». Тенденции в растениеводстве. 3 (12): 473–778. Дои:10.1016 / S1360-1385 (98) 01340-5.
- ^ Paul, L.R .; Chapman, B.K .; Чануэй, К. (2007). «Азотфиксация, связанная с Suillus tomentosus Tuberculate Ectomycorrhizae on Pinus contorta var. широколистная". Анналы ботаники. 99 (6): 1101–1109. Дои:10,1093 / aob / mcm061. ЧВК 3243579. PMID 17468111.