Раковые стволовые клетки - Cancer stem cell
Эта статья слишком полагается на Рекомендации к основные источники. (Июль 2019) (Узнайте, как и когда удалить этот шаблон сообщения) |
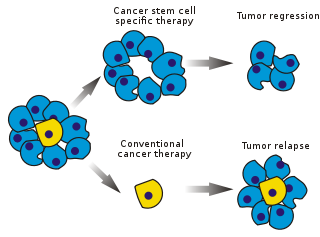
Раковые стволовые клетки (CSC) находятся рак клетки (находятся внутри опухоли или же гематологические раки ), которые обладают характеристиками, связанными с нормальным стволовые клетки, в частности, способность дать начало всем типы клеток найдено в конкретном образце рака. Таким образом, CSC онкогенный (образующие опухоль), возможно, в отличие от других неканцерогенных раковых клеток.[1] CSCs могут генерировать опухоли посредством процессов самообновления и дифференцировки стволовых клеток на несколько типов клеток. Предполагается, что такие клетки сохраняются в опухолях как отдельная популяция и вызывают рецидив и метастаз вызывая новые опухоли. Таким образом, разработка специфических методов лечения, направленных на РСК, дает надежду на улучшение выживаемости и качества жизни онкологических больных, особенно пациентов с метастатическое заболевание.
Существующие методы лечения рака в основном были разработаны на основе животные модели, где методы лечения, способствующие уменьшению опухоли, были признаны эффективными. Однако животные не могут служить полной моделью болезни человека. В частности, у мышей, продолжительность жизни которых не превышает двух лет, трудно исследовать рецидив опухоли.
На начальных этапах тестирования эффективность лечения рака часто измеряется долей абляции массы опухоли (дробное убийство ). Поскольку CSC образуют небольшую часть опухоли, это не обязательно означает выбор препаратов, которые действуют специфически на стволовые клетки. Теория предполагает, что обычные химиотерапия убивают дифференцированные или дифференцирующиеся клетки, которые составляют основную часть опухоли, но не генерируют новые клетки. Популяция РСК, которая его породила, могла остаться нетронутой и вызвать рецидив.
Раковые стволовые клетки были впервые идентифицированы Джон Дик в острый миелоидный лейкоз в конце 1990-х гг. С начала 2000-х они стали интенсивным исследования рака фокус.[2] Сам термин был придуман биологами в очень цитируемой статье 2001 года. Танништа Рейя, Шон Дж. Моррисон, Майкл Ф. Кларк и Ирвинг Вайсман.[3]
Модели распространения опухоли
В разных опухоль подтипы, клетки в популяции опухоли проявляют функциональные неоднородность и опухоли образуются из клетки с различными пролиферативными и дифференциация мощности.[4] Эта функциональная неоднородность среди раковые клетки привело к созданию нескольких моделей распространения для учета гетерогенности и различий в способности к регенерации опухолей: раковых стволовых клеток (CSC) и стохастической модели. Тем не менее, некоторые точки зрения утверждают, что это разграничение является искусственным, поскольку оба процесса действуют взаимодополняющим образом, когда речь идет о реальных популяциях опухолей.[1]


Модель раковых стволовых клеток
Модель раковых стволовых клеток, также известная как Иерархическая модель, предполагает, что опухоли иерархически организованы (ОСК, лежащие на вершине[5] (Рис. 3).) Внутри раковой популяции опухолей есть раковые стволовые клетки (РСК), которые являются онкогенными клетками и биологически отличаются от других субпопуляций.[6] У них есть две определяющие особенности: их долговременная способность к самообновлению и их способность дифференцироваться в потомство, которое не является канцерогенным, но все же способствует росту опухоли. Эта модель предполагает, что только определенные субпопуляции раковых стволовых клеток обладают способностью управлять прогрессированием рака, а это означает, что существуют определенные (внутренние) характеристики, которые могут быть идентифицированы и затем нацелены на долгосрочное уничтожение опухоли без необходимости бороться с целая опухоль.[7]
Стохастическая модель
Чтобы клетка стала злокачественной, она должна претерпеть значительное количество изменений в своей последовательности ДНК. Эта клеточная модель предполагает, что эти мутации могут произойти в любой клетке тела, что приведет к раку. По сути, эта теория предполагает, что все клетки обладают способностью быть онкогенными, что делает все опухолевые клетки равносильными со способностью к самообновлению или дифференцировке, что приводит к гетерогенности опухоли, в то время как другие могут дифференцироваться в не-ОСК. [6][8] На потенциал клетки могут влиять непредсказуемые генетические или эпигенетический факторы, приводящие к образованию фенотипически различных клеток как в онкогенных, так и в неканцерогенных клетках, составляющих опухоль. Согласно «стохастической модели» (или «модели клональной эволюции») каждая раковая клетка в опухоли может получить способность к самообновлению и дифференцировке в многочисленные и гетерогенные клоны раковых клеток, которые ставят под угрозу опухоль.[9]
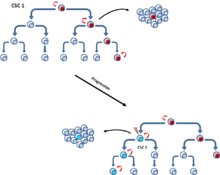
Эти мутации могут постепенно накапливаться и повышать устойчивость и приспособленность клеток, что позволяет им побеждать другие опухолевые клетки, более известные как соматическая эволюция модель.[6] Модель клональной эволюции, которая встречается как в модели CSC, так и в стохастической модели, постулирует, что мутантные опухолевые клетки с преимуществом роста опережают другие. Клетки в доминирующей популяции обладают аналогичным потенциалом инициирования роста опухоли.[10] (Рис. 4).

[11] Эти две модели не исключают друг друга, так как сами ОСК претерпевают клональную эволюцию. Таким образом, вторичные более доминантные РСК могут появиться, если мутация придает более агрессивные свойства.[12] (Рис. 5).
Связывание CSC и стохастических моделей вместе
Исследование, проведенное в 2014 году, утверждает, что разрыв между этими двумя противоречивыми моделями можно преодолеть, предоставив альтернативное объяснение неоднородности опухоли. Они демонстрируют модель, которая включает аспекты как стохастической, так и CSC модели.[8] Они исследовали рак пластичность стволовых клеток при котором раковые стволовые клетки могут переходить между нераковыми стволовыми клетками (не-CSC) и CSC через на месте поддержка более стохастической модели.[8][13] Но существование как биологически разных популяций, не относящихся к CSC, так и CSC, поддерживает модель, в большей степени CSC, предполагая, что обе модели могут играть жизненно важную роль в гетерогенности опухоли.[8]
Иммунологическая модель раковых стволовых клеток
Эта модель предполагает, что иммунологические свойства могут быть важны для понимания туморогенеза и гетерогенности. Таким образом, CSC могут быть очень редкими в некоторых опухолях,[14] но некоторые исследователи обнаружили, что большая часть опухолевых клеток может инициировать опухоли, если их трансплантировать мышам с тяжелым иммунодефицитом,[15] и, таким образом, подвергает сомнению актуальность редких CSC. Однако обе стволовые клетки[16] и CSC[17] обладают уникальными иммунологическими свойствами, которые делают их очень устойчивыми к иммунному надзору. Таким образом, только CSC могут быть способны засеивать опухоли у пациентов с функциональным иммунным надзором, и иммунная привилегия может быть ключевым критерием для идентификации CSC.[18] Более того, модель предполагает, что ОСК могут изначально зависеть от ниш стволовых клеток и что ОСК могут функционировать там как резервуар, в котором мутации могут накапливаться в течение десятилетий без ограничений со стороны иммунной системы. Клинически явные опухоли могут расти, если: A) ОСК теряют зависимость от нишевых факторов (менее дифференцированные опухоли), B) их потомки высокопролиферативных, но изначально иммуногенных нормальных опухолевых клеток развиваются, что позволяет избежать иммунного надзора или C) иммунная система может потерять свою способность подавлять опухоли, например из-за старения.[18]
Дебаты
Существование CSC обсуждается, потому что многие исследования не обнаружили клеток с их специфическими характеристиками.[14] Раковые клетки должны обладать способностью к непрерывной пролиферации и самообновлению, чтобы сохранять множество мутаций, необходимых для канцерогенез и для поддержания роста опухоли, поскольку дифференцированные клетки (сдерживаемые Предел Хейфлика[19]) не может делиться бесконечно. С терапевтической точки зрения, если большинство опухолевых клеток наделены свойствами стволовых клеток, прямое нацеливание на размер опухоли является действенной стратегией. Если CSC составляют незначительное меньшинство, нацеливание на них может быть более эффективным. Другой спор идет о происхождении РСК - от нарушения регуляции нормальных стволовых клеток или от более специализированной популяции, которая приобрела способность к самообновлению (что связано с проблемой стволовых клеток. пластичность ). Эти дебаты мешают открытию того, что многие раковые клетки демонстрируют фенотипическая пластичность под терапевтическим вызовом, изменяя свои транскриптомы до более стеблевого состояния, чтобы избежать разрушения.[нужна цитата ]
Свидетельство
Первое убедительное доказательство наличия РСК появилось в 1997 году. Боннет и Дик выделили субпопуляцию лейкозных клеток, экспрессирующих поверхностный маркер. CD34, но нет CD38.[20] Авторы установили, что CD34+/ CD38− субпопуляция способна инициировать опухоли в NOD /SCID мышей, гистологически похожих на донора. Первые доказательства наличия стволовых клеток рака солидной опухоли последовали в 2002 году с открытием клоногенной сферической клетки, выделенной и охарактеризованной из мозга взрослого человека. глиомы. Человеческий корковый глиальный опухоли содержат нервные стволовые клетки, экспрессирующие астроглиальные и нейрональные маркеры in vitro.[21] Было показано, что раковые стволовые клетки, выделенные из глиом взрослого человека, индуцируют опухоли, которые напоминают исходную опухоль, при трансплантации в интракраниальные модели голых мышей.[22]
В исследования рака эксперименты, опухолевые клетки иногда вводят в экспериментальное животное установить опухоль. Затем вовремя отслеживают прогрессирование заболевания, и можно проверить эффективность новых лекарств. Для образования опухоли необходимо ввести тысячи или десятки тысяч клеток. Классически это объяснялось плохой методологией (то есть опухолевые клетки теряли свою жизнеспособность во время переноса) или критической важности микросреды, конкретного биохимического окружения вводимых клеток. Сторонники парадигмы CSC утверждают, что только небольшая часть инъецированных клеток, CSC, имеет потенциал для образования опухоли. В человеческом острый миелоидный лейкоз частота этих клеток составляет менее 1 из 10 000.[20]
Дальнейшие доказательства получены из гистология. Многие опухоли неоднородный и содержать несколько типы клеток родной для принимающего органа. Гетерогенность опухоли обычно сохраняется за счет опухоли. метастазы. Это говорит о том, что клетка, которая их произвела, обладала способностью генерировать несколько типов клеток, что является классическим признаком стволовые клетки.[20]
Существование стволовых клеток лейкемии побудило исследовать другие виды рака. РСК недавно были идентифицированы в нескольких солидных опухолях, включая:
- Мозг[23]
- Грудь[24]
- Двоеточие[25]
- Яичник[26][27]
- Поджелудочная железа[28]
- Предстательная железа[29][30]
- Меланома[31][32][33][34]
- Множественная миелома[35][36]
- Немеланомный рак кожи[37][38]
Механистические и математические модели
После того, как будут выдвинуты гипотезы о путях к раку, можно разработать прогнозные математический модели[39] например, на основе метод сотового отсека. Например, рост аномальных клеток может быть обозначен определенной вероятностью мутации. Такая модель предсказывала, что повторное повреждение зрелых клеток увеличивает образование аномального потомства и риск рака.[40] Клиническая эффективность таких моделей[41] остается неустановленным.
Источник

Происхождение CSC - активная область исследований. Ответ может зависеть от типа опухоли и фенотип. До сих пор гипотеза о том, что опухоли происходят из одной «клетки происхождения», не была продемонстрирована с использованием модели раковых стволовых клеток. Это потому, что раковые стволовые клетки не присутствуют в опухолях на конечной стадии.
Гипотезы происхождения включают мутанты в развивающихся стволовых или предшественниках, мутанты в взрослые стволовые клетки или взрослые клетки-предшественники и мутантные дифференцированные клетки, которые приобретают свойства стебля. Эти теории часто сосредотачиваются на «клетке происхождения» опухоли.
Гипотезы
Мутации стволовых клеток
«Мутация в ниша стволовых клеток Популяции во время развития »гипотеза утверждает, что эти развивающиеся стволовые популяции мутируют, а затем воспроизводятся так, что мутация является общей для многих потомков. Эти дочерние клетки гораздо ближе к тому, чтобы стать опухолями, и их количество увеличивает вероятность раковой мутации.[42]
Взрослые стволовые клетки
Другая теория связывает взрослые стволовые клетки (ASC) с образованием опухоли. Чаще всего это связано с тканями с высокой скоростью обновления клеток (такими как кожа или же кишка ). В этих тканях ASC являются кандидатами из-за их частого деления клеток (по сравнению с большинством ASC) в сочетании с длительным сроком службы ASC. Эта комбинация создает идеальный набор обстоятельств для накопления мутаций: накопление мутаций является основным фактором, движущим начало рака. Фактические данные показывают, что эта связь представляет собой реальное явление, хотя определенные виды рака были связаны с определенной причиной.[43][44]
Де-дифференциация
Дедифференцировка мутировавших клеток может создавать характеристики, подобные стволовым клеткам, предполагая, что любая клетка может стать раковой стволовой клеткой. Другими словами, полностью дифференцированная клетка претерпевает мутации или внеклеточные сигналы, которые возвращают ее в стеблевое состояние. Эта концепция была продемонстрирована совсем недавно в рак простаты модели, в которых клетки подвергаются андрогенная депривационная терапия временно изменяют свой транскриптом на транскриптом нервный гребень стволовая клетка с инвазивными и мультипотентными свойствами этого класса стволовых клеток.[нужна цитата ]
Иерархия
Концепция иерархии опухолей утверждает, что опухоль представляет собой гетерогенную популяцию мутантных клеток, каждая из которых имеет общие мутации, но различается по специфическим характеристикам. фенотип. Опухоль содержит несколько типов стволовых клеток, один из которых оптимален для конкретной среды, а другие - менее успешные. Эти вторичные линии могут быть более успешными в других средах, позволяя опухоли адаптироваться, включая адаптацию к терапевтическому вмешательству. Если это правильно, эта концепция влияет на режимы лечения, специфичные для раковых стволовых клеток.[45] Такая иерархия усложнила бы попытки установить происхождение.
Идентификация
РСК, о которых в настоящее время сообщается в большинстве опухолей человека, обычно идентифицируют и обогащают с помощью стратегий выявления нормальных стволовых клеток, сходных во всех исследованиях.[46] Эти процедуры включают сортировка клеток с активацией флуоресценции (FACS), с антителами, направленными на маркеры клеточной поверхности, и функциональными подходами, включая анализ побочной популяции или анализ Aldefluor.[47] Затем результат, обогащенный CSC, имплантируют в различных дозах мышам с иммунодефицитом для оценки способности развития опухоли. Этот in vivo анализ называется анализом предельного разведения. Подмножества опухолевых клеток, которые могут инициировать развитие опухоли при низком количестве клеток, дополнительно проверяются на способность к самообновлению в серийных исследованиях опухолей.[48]
РСК также можно идентифицировать по оттоку инкорпорированных Hoechst красит через множественная лекарственная устойчивость (MDR) и АТФ-связывающая кассета (ABC) Транспортеры.[47]
Другой подход - это сферические анализы. Многие нормальные стволовые клетки Такие как кроветворный или стволовые клетки из ткани в особых условиях культивирования образуют трехмерные сферы, которые могут различаться. Как и нормальные стволовые клетки, РСК, выделенные из мозг или же опухоли простаты также имеют возможность формировать сферы, не зависящие от якоря.[49]
Неоднородность (маркеры)
Гетерогенность ОСК представляет собой пул дифференцированных и недифференцированных опухолевых клеток, которые пополняются клетками, обладающими свойствами как опухолевым, так и стволовыми клетками и фенотипической и метаболической гетерогенностью внутри единой опухолевой массы. Существуют две теории, объясняющие фенотипическую и метаболическую гетерогенность РСК; клональные вариации и теория раковых стволовых клеток. В то время как предыдущая теория диктует роль генетической, эпигенетической и микросреды, в которой обитают опухолевые клетки, для приобретения недифференцированных онкогенных признаков. Последняя теория больше сосредотачивается на признаках злокачественности, приобретаемых стволовыми клетками, когда эти недифференцированные и сильно туморогенные стволовые клетки повторно заселяют дифференцированную опухолевую массу. [50]
CSC были идентифицированы в различных солидные опухоли. Обычно маркеры, специфичные для нормальных стволовых клеток, используются для выделения CSC из солидных и гематологических опухолей. Маркеры, наиболее часто используемые для выделения CSC, включают: CD133 (также известный как ПРОМ1 ), CD44, ALDH1A1, CD34, CD24 и EpCAM (молекула адгезии эпителиальных клеток, также известный как эпителиальный специфический антиген, ЕКА ).[51]
CD133 (проминин 1) - пятикомпонентныйтрансмембранный домен гликопротеин выражено на CD34+ стебель и клетки-предшественники, в предшественниках эндотелия и нервные стволовые клетки плода. Он был обнаружен с помощью гликозилированный эпитоп известный как AC133.
EpCAM (молекула адгезии эпителиальных клеток, ESA, TROP1) представляет собой гемофильный Ca2+-независимая молекула клеточной адгезии, экспрессирующаяся на базолатеральной поверхности большинства эпителиальные клетки.
CD90 (THY1) - это гликозилфосфатидилинозитол гликопротеин закреплен в плазматической мембране и участвует в преобразование сигнала. Он также может опосредовать адгезию между тимоциты и строма тимуса.
CD44 (PGP1) представляет собой молекулу адгезии, которая имеет плейотропный роли в передаче сигналов, миграции и хоуминга клеток. Он имеет несколько изоформ, включая CD44H, который проявляет высокое сродство к гиалуронат и CD44V, обладающий метастатическими свойствами.
CD24 (HSA) - это гликозилированный гликозилфосфатидилинозит-заякоренная адгезионная молекула, которая играет костимуляторную роль в B и Т-клетки.
CD200 (OX-2) - мембрана типа 1 гликопротеин, который доставляет подавляющий сигнал иммунным клеткам, включая Т-клетки, естественные клетки-киллеры и макрофаги.
ALDH повсеместно альдегиддегидрогеназа семейство ферментов, которое катализирует окисление ароматические альдегиды к карбоксильные кислоты. Например, он играет роль в превращении ретинола в ретиноевая кислота, что необходимо для выживания.[52][53]
Первым солидным злокачественным новообразованием, из которого были выделены и идентифицированы ОСК, было рак молочной железы и они наиболее изучены. РСК груди обогащены CD44+CD24- / низкий,[54] SP[55] и ALDH+ субпопуляции.[56][57] РСК груди очевидно фенотипически разнообразный. Экспрессия маркера CSC в клетках рака молочной железы, по-видимому, неоднородна, и популяции CSC груди варьируются в зависимости от опухоли.[58] Оба CD44+CD24− и CD44+CD24+ популяции клеток представляют собой клетки, инициирующие опухоль; однако наиболее высокообогащаются CSC при использовании профиля маркера CD44.+CD49fЗдравствуйCD133 / 2Здравствуй.[59]
РСК были зарегистрированы во многих опухолях головного мозга. Стеблоподобные опухолевые клетки были идентифицированы с использованием маркеров клеточной поверхности, включая CD133,[60] SSEA-1 (стадийно-специфический эмбриональный антиген-1),[61] EGFR[62] и CD44.[63] Использование CD133 для идентификации стволовых клеток опухоли головного мозга может быть проблематичным, поскольку онкогенные клетки обнаруживаются в обоих CD133.+ и CD133− клетки в некоторых глиомы и немного CD133+ опухолевые клетки головного мозга могут не обладать способностью инициировать опухоль.[62]
Сообщалось о CSC у человека рак толстой кишки.[25] Для их идентификации маркеры клеточной поверхности, такие как CD133,[25] CD44[64] и ABCB5,[65] функциональный анализ, включая клональный анализ [66] и Aldefluor анализ.[67] Использование CD133 в качестве положительного маркера для CSC толстой кишки дало противоречивые результаты. Эпитоп AC133, но не белок CD133, специфически экспрессируется в CSC толстой кишки, и его экспрессия теряется при дифференцировке.[68] Кроме того, CD44+ клетки рака толстой кишки и дополнительное субфракционирование CD44+EpCAM+ Популяция клеток с CD166 увеличивает успех приживления опухоли.[64]
Несколько CSC были зарегистрированы в предстательная железа,[69] легкое и многие другие органы, в том числе печень, поджелудочная железа, почка или же яичник.[52][70] В рак простаты клетки, инициирующие опухоль, были идентифицированы в CD44+[71] подмножество клеток как CD44+α2β1+,[72] TRA-1-60+CD151+CD166+ [73] или ALDH+ [74] клеточные популяции. Предполагаемые маркеры для легкое Сообщалось о CSC, включая CD133+,[75] ALDH+,[76] CD44+ [77] и онкофетальный белок 5T4+.[78]
Метастаз
Метастазы - основная причина летальности опухоли. Однако не все опухолевые клетки могут давать метастазы.[79] Этот потенциал зависит от факторов, определяющих рост, ангиогенез, вторжение и другие основные процессы.
Эпителиально-мезенхимальный переход
В эпителиальных опухолях эпителиально-мезенхимальный переход (ЕМТ) считается важным событием.[80] EMT и обратный переход от мезенхимальный к эпителиальному фенотипу (ВСТРЕТИЛИСЬ ) участвуют в эмбриональное развитие, который включает нарушение эпителиальной клетки гомеостаз и приобретение мигрирующего мезенхимального фенотипа.[81] EMT, по-видимому, контролируется каноническими путями, такими как WNT и трансформирующий фактор роста β.[82]
Важной особенностью EMT является потеря мембраны E-кадгерин в прилипает к стыкам, куда β-катенин может сыграть значительную роль. Транслокация β-катенина из соединений адгезивов в ядро может привести к потере Е-кадгерина и впоследствии к ЕМТ. Ядерный β-катенин, по-видимому, может напрямую транскрипционно активировать ЕМТ-ассоциированную мишень. гены, например ген E-кадгерина репрессор SLUG (также известный как SNAI2 ).[83] Механические свойства микросреда опухоли, Такие как гипоксия, может способствовать выживанию РСК и метастатическому потенциалу за счет стабилизации факторы, индуцируемые гипоксией через взаимодействие с ROS (активные формы кислорода ).[84][85]
Опухолевые клетки, подвергающиеся EMT, могут быть предшественниками метастатических раковых клеток или даже метастатических РСК.[86][87] В инвазивном краю карцинома поджелудочной железы, подмножество CD133+CXCR4+ (рецептор для CXCL12 хемокин также известный как SDF1 лиганд ) клеток. Эти клетки проявляли значительно более сильную миграционную активность, чем их аналог CD133.+CXCR4− клеток, но оба показали сходную способность к развитию опухоли.[88] Более того, подавление Рецептор CXCR4 снижение метастатического потенциала без изменения онкогенной способности.[89]
Двухфазный образец выражения
При раке груди CD44+CD24- / низкий клетки обнаруживаются в метастатических плевральных выпотах.[24] Напротив, повышенное количество CD24+ клетки были идентифицированы в отдаленных метастазах у пациентов с раком груди.[90] Возможно, что CD44+CD24- / низкий клетки первоначально метастазируют и в новом месте изменяют свой фенотип и претерпевают ограниченную дифференцировку.[91] Гипотеза о двухфазном паттерне экспрессии предполагает две формы раковых стволовых клеток - стационарные (SCS) и мобильные (MCS). SCS встраиваются в ткань и сохраняются в дифференцированных областях на протяжении всего развития опухоли. MCS расположены на границе опухоль-хозяин. Эти клетки, по-видимому, происходят из SCS посредством получения временной EMT (Рисунок 7).[92]

Подразумеваемое
CSC имеют значение для терапии рака, в том числе для идентификации заболевания, селективных мишеней для лекарств, предотвращения метастазов и стратегий вмешательства.
Уход
CSC по своей природе более устойчивы к химиотерапевтические агенты. Этому способствуют 5 основных факторов:[93]
- 1. Их ниша защищает их от контакта с большими концентрациями противораковых препаратов.
- 2. Они экспрессируют различные трансмембранные белки, такие как MDR1 и BCRP, которые выкачивают лекарства из цитоплазмы.
- 3. Они делятся медленно, как взрослые стволовые клетки имеют тенденцию к этому и, таким образом, не уничтожаются химиотерапевтическими агентами, которые нацелены на быстро реплицирующиеся клетки посредством повреждения ДНК или ингибирования митоза.
- 4. Они активируют белки восстановления повреждений ДНК.
- 5. Для них характерна чрезмерная активация антиапоптотических сигнальных путей.
После химиотерапевтического лечения выжившие ОСК способны повторно заселить опухоль и вызвать рецидив. Для предотвращения этого необходимо дополнительное лечение, направленное на удаление РСК в дополнение к раковым соматическим клеткам.
Таргетинг
Селективное нацеливание на CSC может позволить лечить агрессивные, неоперабельные опухоли, а также предотвратить метастазирование и рецидив. Гипотеза предполагает, что после удаления РСК рак может регрессировать из-за дифференцировки и / или гибели клеток.[нужна цитата ] Неясна доля опухолевых клеток, которые являются РСК и поэтому необходимо удалить.[94]
Исследования искали конкретные маркеры[24] и для протеомный и геномный сигнатуры опухолей, которые отличают РСК от других.[95] В 2009 году ученые идентифицировали соединение салиномицин, что избирательно снижает долю РСК груди у мышей более чем в 100 раз по сравнению с Паклитаксел, широко используемый химиотерапевтический агент.[96] Некоторые типы раковых клеток могут выжить при лечении салиномицином через аутофагия,[97] посредством чего клетки используют кислые органеллы Такие как лизосомы разлагать и перерабатывать определенные типы белков. Использование ингибиторов аутофагии может убить раковые стволовые клетки, которые выживают за счет аутофагии.[98]
Рецептор интерлейкина-3-альфа рецептора клеточной поверхности (CD123) сверхэкспрессируется на CD34 + CD38- лейкозных стволовых клетках (LSC) в острый миелолейкоз (AML), но не на нормальных CD34 + CD38- Костный мозг клетки.[99] Лечение мышей NOD / SCID с привитым AML CD123-специфическим моноклональное антитело нарушение перемещения LSC в костный мозг и снижение общей репопуляции клеток AML, включая долю LSC у вторичных реципиентов мышей.[100]
В исследовании 2015 года наночастицы miR-34a и бикарбонат аммония и доставили их в CSC простаты на мышиной модели. Затем они облучали область ближним инфракрасным лазером. Это привело к тому, что наночастицы набухли в три раза или более в размере, разрывая эндосомы и рассеивая РНК в клетке. miR-34a может снизить уровень CD44.[101][102]
В исследовании 2018 года были выявлены ингибиторы семейства ферментов ALDH1A и показано, что они могут избирательно истощать предполагаемые раковые стволовые клетки в нескольких линиях клеток рака яичников.[103]
Пути
В разработка новых лекарств для нацеливания на ОСК требуется понимание клеточных механизмов, регулирующих пролиферацию клеток. Первые успехи в этой области были сделаны с использованием гемопоэтических стволовых клеток (HSC) и их трансформированных аналогов в лейкемия, заболевание, для которого лучше всего изучено происхождение CSC. Стволовые клетки многих органов имеют те же клеточные пути, что и HSC, происходящие от лейкемии.
Нормальный стволовая клетка может трансформироваться в CSC посредством нарушения регуляции пролиферации и дифференцировки пути контролируя это или побуждая онкопротеин Мероприятия.
ИМТ-1
В Группа Поликомб транскрипционный репрессор Бми-1 был обнаружен как обычное онкоген активирован в лимфома[104] и позже показано, что они регулируют HSC.[105] Роль Bmi-1 была продемонстрирована на нервных стволовых клетках.[106] Путь, по-видимому, активен в РСК педиатрический опухоли головного мозга.[107]
Notch
В Notch pathway играет роль в контроле пролиферации стволовых клеток для нескольких типов клеток, включая гематопоэтические, нервные и молочные[108] СК. Было высказано предположение, что компоненты этого пути действуют как онкогены в молочных железах.[109] и другие опухоли.
Ответвление сигнального пути Notch, в котором участвует фактор транскрипции. Hes3 регулирует количество культивируемых клеток с характеристиками CSC, полученными от пациентов с глиобластомой.[110]
Соник-ежик и Wnt
Эти пути развития являются регуляторами SC.[111][112] Обе Соник ежик (SHH) и Wnt проводящие пути обычно гиперактивированы в опухолях и необходимы для поддержания роста опухоли. Однако факторы транскрипции Gli, которые регулируются SHH, получили свое название от глиомы, где они сильно выражены. Степень перекрестные помехи существует между двумя путями, и они обычно активируются вместе.[113] Напротив, при раке толстой кишки передача сигналов hedgehog, по-видимому, противодействует Wnt.[114]
Доступны блокаторы звукового ежа, такие как циклопамин. Водорастворимый циклопамин может быть более эффективным при лечении рака. DMAPT, водорастворимое производное партенолид, вызывает окислительный стресс и подавляет NF-κB сигнализация[115] для AML (лейкемии) и, возможно, миеломы и рака простаты. Теломераза является предметом изучения физиологии CSC.[116] GRN163L (Иметелстат ) недавно были начаты испытания, направленные на нацеливание на стволовые клетки миеломы.
Передача сигналов Wnt может стать независимой от обычных стимулов посредством мутаций в последующих онкогенах и генах-супрессорах опухолей, которые становятся постоянно активированными, даже если нормальный рецептор не получил сигнала. β-катенин связывается с факторами транскрипции, такими как белок TCF4 и в сочетании молекулы активируют необходимые гены. LF3 сильно ингибирует это связывание in vitro, в клеточных линиях и снижение роста опухоли на моделях мышей. Это препятствовало репликации и уменьшало их способность к миграции, не затрагивая здоровые клетки. После лечения не осталось раковых стволовых клеток. Открытие было результатом "рациональный дизайн лекарств ", включая AlphaScreens и ELISA технологии.[117]
Рекомендации
- ^ а б Шрипадманабх М., Толи Б.Дж. (2018). «Исследования ниши раковых стволовых клеток с использованием трехмерных моделей опухолей in vitro и микрофлюидики». Достижения биотехнологии. 36 (4): 1094–1110. Дои:10.1016 / j.biotechadv.2018.03.009. PMID 29559382.
- ^ Мукерджи, Сиддхартха (29 октября 2010 г.). "Раковая спящая клетка". Нью-Йорк Таймс. Получено 15 июля 2014.
- ^ Рейя Т., Моррисон С.Дж., Кларк М.Ф., Вайсман И.Л. (ноябрь 2001 г.). «Стволовые клетки, рак и раковые стволовые клетки». Природа. 414 (6859): 105–11. Дои:10.1038/35102167. HDL:2027.42/62862. PMID 11689955.
- ^ Хеппнер GH, Миллер BE (1983). «Гетерогенность опухоли: биологические последствия и терапевтические последствия». Отзывы о метастазах рака. 2 (1): 5–23. Дои:10.1007 / BF00046903. PMID 6616442.
- ^ Боннет Д., Дик Дж. Э. (июль 1997 г.). «Острый миелоидный лейкоз человека организован в виде иерархии, происходящей от примитивной гемопоэтической клетки». Природа Медицина. 3 (7): 730–7. Дои:10,1038 / нм0797-730. PMID 9212098.
- ^ а б c Beck B, Blanpain C (2013), «Раскрытие потенциала раковых стволовых клеток», Нат Рев Рак, 13 (10): 727–38, Дои:10.1038 / nrc3597, PMID 24060864
- ^ Шеклтон М., Кинтана Е., Фирон ER, Моррисон С.Дж. (сентябрь 2009 г.). «Неоднородность рака: раковые стволовые клетки против клональной эволюции». Клетка. 138 (5): 822–9. Дои:10.1016 / j.cell.2009.08.017. PMID 19737509.
- ^ а б c d Wang W, Quan Y, Fu Q, Liu Y, Liang Y, Wu J, Yang G, Luo C, Ouyang Q, Wang Y (2014), «Динамика между субпопуляциями раковых клеток выявляет модель, согласованную как с иерархической, так и с стохастической концепциями» , PLOS ONE, 9 (1): e84654, Bibcode:2014PLoSO ... 984654W, Дои:10.1371 / journal.pone.0084654, ЧВК 3886990, PMID 24416258
- ^ Кресо А., Дик Дж. Э. (2014), «Эволюция модели раковых стволовых клеток», Стволовая клетка, 14 (3): 275–91, Дои:10.1016 / j.stem.2014.02.006, PMID 24607403
- ^ Барабе Ф., Кеннеди Дж. А., Хоуп К. Дж., Дик Дж. Э. (апрель 2007 г.). «Моделирование возникновения и прогрессирования острого лейкоза человека у мышей». Наука. 316 (5824): 600–4. Bibcode:2007 Наука ... 316..600B. Дои:10.1126 / science.1139851. PMID 17463288.
- ^ Nowell PC (октябрь 1976 г.). «Клональная эволюция популяций опухолевых клеток». Наука. 194 (4260): 23–8. Bibcode:1976Научный ... 194 ... 23N. Дои:10.1126 / science.959840. PMID 959840.
- ^ Кларк Э.А., Голуб Т.Р., Лендер Э.С., Хайнс РО (август 2000 г.). «Геномный анализ метастазов показывает важную роль RhoC». Природа. 406 (6795): 532–5. Дои:10.1038/35020106. PMID 10952316.
- ^ Cabrera MC, Hollingsworth RE, Hurt EM (2015), «Пластичность раковых стволовых клеток и иерархия опухолей», Стволовые клетки World J, 7 (1): 27–36, Дои:10.4252 / wjsc.v7.i1.27, ЧВК 4300934, PMID 25621103
- ^ а б Гупта П.Б., Чаффер К.Л., Вайнберг Р.А. (сентябрь 2009 г.). «Раковые стволовые клетки: мираж или реальность?». Природа Медицина. 15 (9): 1010–2. Дои:10,1038 / нм0909-1010. PMID 19734877.[постоянная мертвая ссылка ]>
- ^ Кинтана Э., Шеклтон М., Сабель М.С., Фуллен Д.Р., Джонсон TM, Моррисон С.Дж. (декабрь 2008 г.). «Эффективное образование опухолей отдельными клетками меланомы человека». Природа. 456 (7222): 593–8. Bibcode:2008Натура.456..593Q. Дои:10.1038 / природа07567. ЧВК 2597380. PMID 19052619.
- ^ Ичирю Н, Фэйрчайлд П.Дж. (2013). «Иммунные привилегии стволовых клеток». В Завазава Н. (ред.). Иммунобиология эмбриональных стволовых клеток. Методы молекулярной биологии. 1029. С. 1–16. Дои:10.1007/978-1-62703-478-4_1. ISBN 978-1-62703-477-7. PMID 23756938.
- ^ Maccalli C, Volontè A, Cimminiello C, Parmiani G (февраль 2014 г.). «Иммунология раковых стволовых клеток в солидных опухолях. Обзор». Европейский журнал рака. 50 (3): 649–55. Дои:10.1016 / j.ejca.2013.11.014. PMID 24333096.
- ^ а б Бруттель В.С., Вишхузен Дж. (2014). «Иммунология раковых стволовых клеток: ключ к пониманию туморогенеза и ускользания от опухолевого иммунитета?». Границы иммунологии. 5: 360. Дои:10.3389 / fimmu.2014.00360. ЧВК 4114188. PMID 25120546.
- ^ Хейфлик Л. (март 1965 г.). «Ограниченное время жизни штаммов диплоидных клеток человека in vitro». Экспериментальные исследования клеток. 37 (3): 614–36. Дои:10.1016/0014-4827(65)90211-9. PMID 14315085.
- ^ а б c Капот D, Дик Дж. Э. (Июль 1997 г.). «Острый миелоидный лейкоз человека организован в виде иерархии, происходящей от примитивной гемопоэтической клетки». Природа Медицина. 3 (7): 730–7. Дои:10,1038 / нм0797-730. PMID 9212098.
- ^ Игнатова Т.Н., Кукеков В.Г., Лэйвелл Э.Д., Суслов О.Н., Врионис Ф.Д., Штейндлер Д.А. (сентябрь 2002 г.). «Человеческие кортикальные глиальные опухоли содержат нервные стволовые клетки, экспрессирующие астроглиальные и нейрональные маркеры in vitro». Глия. 39 (3): 193–206. Дои:10.1002 / glia.10094. PMID 12203386.
- ^ Сундар С.Дж., Се Дж.К., Манджила С., Латиа Д.Д., Слоан А. (декабрь 2014 г.). «Роль раковых стволовых клеток в глиобластоме». Нейрохирургия. 37 (6): E6. Дои:10.3171 / 2014.9.FOCUS14494. PMID 25434391.
- ^ Сингх С.К., Кларк И.Д., Терасаки М., Бонн В.Э., Хокинс С., Сквайр Дж., Диркс П.Б. (сентябрь 2003 г.). «Идентификация раковых стволовых клеток в опухолях головного мозга человека». Исследования рака. 63 (18): 5821–8. PMID 14522905.
- ^ а б c Аль-Хадж М., Вича М.С., Бенито-Эрнандес А., Моррисон С.Дж., Кларк М.Ф. (апрель 2003 г.). «Проспективная идентификация онкогенных клеток рака груди». Труды Национальной академии наук Соединенных Штатов Америки. 100 (7): 3983–8. Bibcode:2003ПНАС..100.3983А. Дои:10.1073 / пнас.0530291100. ЧВК 153034. PMID 12629218.
- ^ а б c О'Брайен, Калифорния, Поллетт А., Галлинджер С., Дик Дж. Э. (январь 2007 г.). «Клетка рака толстой кишки человека, способная инициировать рост опухоли у мышей с иммунодефицитом». Природа. 445 (7123): 106–10. Bibcode:2007Натура.445..106O. Дои:10.1038 / природа05372. PMID 17122772.
- ^ Чжан С., Балч С., Чан М.В., Лай Х.С., Матей Д., Шильдер Дж.М., Ян П.С., Хуанг Т.Х., племянник К.П. (июнь 2008 г.). «Идентификация и характеристика клеток, вызывающих рак яичников из первичных опухолей человека». Исследования рака. 68 (11): 4311–20. Дои:10.1158 / 0008-5472.CAN-08-0364. ЧВК 2553722. PMID 18519691.
- ^ Альверо А.Б., Чен Р., Фу Х.Х., Монтанья М., Шварц П.Е., Резерфорд Т., Силаси Д.А., Стеффенсен К.Д., Вальдстром М., Визинтин И., Мор Г. (январь 2009 г.). «Молекулярное фенотипирование стволовых клеток рака яичников человека раскрывает механизмы восстановления и химиорезистентности». Клеточный цикл. 8 (1): 158–66. Дои:10.4161 / cc.8.1.7533. ЧВК 3041590. PMID 19158483.
- ^ Ли К., Хайдт Д.Г., Далерба П., Бурант К.Ф., Чжан Л., Адсей В., Вича М., Кларк М.Ф., Симеоне Д.М. (февраль 2007 г.). «Идентификация стволовых клеток рака поджелудочной железы». Исследования рака. 67 (3): 1030–7. Дои:10.1158 / 0008-5472.CAN-06-2030. PMID 17283135.
- ^ Maitland NJ, Collins AT (июнь 2008 г.). «Стволовые клетки рака простаты: новая мишень для терапии». Журнал клинической онкологии. 26 (17): 2862–70. Дои:10.1200 / JCO.2007.15.1472. PMID 18539965.
- ^ Ланг Ш., Фрейм FM, Коллинз А. Т. (январь 2009 г.). «Стволовые клетки рака простаты». Журнал патологии. 217 (2): 299–306. Дои:10.1002 / путь.2478. ЧВК 2673349. PMID 19040209.
- ^ Шаттон Т., Мерфи Г.Ф., Франк Нью-Йорк, Ямаура К., Ваага-Гассер А.М., Гассер М., Жан К., Джордан С., Дункан Л.М., Вейсхаупт С., Фульбригге Р.С., Куппер Т.С., Сайег М.Х., Франк М.Х. (январь 2008 г.). «Идентификация клеток, инициирующих меланомы человека». Природа. 451 (7176): 345–9. Bibcode:2008Натура.451..345S. Дои:10.1038 / природа06489. ЧВК 3660705. PMID 18202660.
- ^ Бойко А.Д., Разоренова О.В., ван де Рейн М., Светтер С.М., Джонсон Д.Л., Ли Д.П., Батлер П.Д., Ян Г.П., Джошуа Б., Каплан М.Дж., Лонгакер М.Т., Вайсман Иллинойс (июль 2010 г.). «Клетки, инициирующие меланому человека, экспрессируют рецептор фактора роста нервного гребня CD271». Природа. 466 (7302): 133–7. Bibcode:2010Натура.466..133Б. Дои:10.1038 / природа09161. ЧВК 2898751. PMID 20596026.
- ^ Шмидт П., Копецки С., Хомбах А., Зигрино П., Маух С., Абкен Н. (февраль 2011 г.). «Ликвидация меланом путем целенаправленного уничтожения незначительной субпопуляции опухолевых клеток». Труды Национальной академии наук Соединенных Штатов Америки. 108 (6): 2474–9. Bibcode:2011ПНАС..108.2474С. Дои:10.1073 / pnas.1009069108. ЧВК 3038763. PMID 21282657.
- ^ Чивенни Г., Вальтер А., Коберт Н., Михик-Пробст Д., Ципсер М., Беллони Б., Зейферт Б., Мох Н., Даммер Р., ван ден Брук М., Соммер Л. (апрель 2011 г.). «CD271-положительные стволовые клетки меланомы человека, ассоциированные с метастазами, создают гетерогенность опухоли и долгосрочный рост». Исследования рака. 71 (8): 3098–109. Дои:10.1158 / 0008-5472.CAN-10-3997. PMID 21393506.
- ^ Мацуи В., Хафф Калифорния, Ван К., Малехорн М.Т., Барбер Дж., Танехко Ю., Смит Б.Д., Сивин К.И., Джонс Р.Дж. (март 2004 г.). «Характеристика клоногенных клеток множественной миеломы». Кровь. 103 (6): 2332–6. Дои:10.1182 / кровь-2003-09-3064. ЧВК 3311914. PMID 14630803.
- ^ Мацуи В., Ван К., Барбер Дж. П., Бреннан С., Смит Б. Д., Боррелло И., Макнейс И., Лин Л., Амбиндер Р. Ф., Пикок С., Уоткинс Д. Н., Хафф Калифорния, Джонс Р. Дж. (Январь 2008 г.). «Клоногенные предшественники множественной миеломы, свойства стволовых клеток и лекарственная устойчивость». Исследования рака. 68 (1): 190–7. Дои:10.1158 / 0008-5472.CAN-07-3096. ЧВК 2603142. PMID 18172311.
- ^ Colmont CS, Benketah A, Reed SH, Hawk NV, Telford WG, Ohyama M, Udey MC, Yee CL, Vogel JC, Patel GK (январь 2013 г.). «CD200-экспрессирующие клетки базальноклеточной карциномы человека инициируют рост опухоли». Труды Национальной академии наук Соединенных Штатов Америки. 110 (4): 1434–9. Bibcode:2013PNAS..110.1434C. Дои:10.1073 / pnas.1211655110. ЧВК 3557049. PMID 23292936.
- ^ Патель Г.К., Йи К.Л., Терунума А., Телфорд В.Г., Вунг Н., Юспа С.Х., Фогель Дж.С. (февраль 2012 г.). «Идентификация и характеристика опухолевых клеток в первичной плоскоклеточной карциноме кожи человека». Журнал следственной дерматологии. 132 (2): 401–9. Дои:10.1038 / jid.2011.317. ЧВК 3258300. PMID 22011906.
- ^ Preziosi, Луиджи (2003). Моделирование рака и симуляция. Бока-Ратон: CRC Press. ISBN 978-1-58488-361-6.
- ^ Гангули Р, Пури И.К. (Февраль 2006 г.). «Математическая модель гипотезы раковых стволовых клеток». Распространение клеток. 39 (1): 3–14. Дои:10.1111 / j.1365-2184.2006.00369.x. ЧВК 6495990. PMID 16426418.
- ^ Гангули Р, Пури И.К. (Июнь 2007 г.). «Математическая модель эффективности химиотерапевтических препаратов в остановке роста опухоли на основе гипотезы раковых стволовых клеток». Распространение клеток. 40 (3): 338–54. Дои:10.1111 / j.1365-2184.2007.00434.x. ЧВК 6496895. PMID 17531079.
- ^ Ван Й, Ян Дж., Чжэн Х, Томасек Дж., Чжан П., Маккивер ЧП, Ли ЕЙ, Чжу Й (июнь 2009 г.). «Экспрессия мутантных белков р53 подразумевает родственную связь между нервными стволовыми клетками и злокачественной астроцитарной глиомой в мышиной модели». Раковая клетка. 15 (6): 514–26. Дои:10.1016 / j.ccr.2009.04.001. ЧВК 2721466. PMID 19477430.
- ^ Лопес-Ласаро М (01.01.2015). «Способность стволовых клеток к миграции может объяснить существование рака неизвестной первичной локализации. Переосмысление метастазов». Онкология. 2 (5): 467–75. Дои:10.18632 / oncoscience.159. ЧВК 4468332. PMID 26097879.
- ^ Лопес-Ласаро М (18.08.2015). "Теория деления стволовых клеток рака". Клеточный цикл. 14 (16): 2547–8. Дои:10.1080/15384101.2015.1062330. ЧВК 5242319. PMID 26090957.
- ^ Кларк М.Ф., Дик Дж. Э., Диркс П. Б., Ивз С. Дж., Джеймисон С. К., Джонс Д. Л., Висвадер Дж., Вайсман И. Л., Уол Г. М. (октябрь 2006 г.). «Раковые стволовые клетки - перспективы текущего состояния и будущих направлений: семинар AACR по раковым стволовым клеткам». Исследования рака. 66 (19): 9339–44. Дои:10.1158 / 0008-5472.CAN-06-3126. PMID 16990346.
- ^ Голебевска А., Бронс Н.Х., Бьерквиг Р., Никлоу С.П. (февраль 2011 г.). «Критическая оценка побочного популяционного анализа стволовых клеток и раковых стволовых клеток». Стволовая клетка. 8 (2): 136–47. Дои:10.1016 / j.stem.2011.01.007. PMID 21295271.
- ^ а б Шаренберг CW, Харки М.А., Torok-Storb B (январь 2002 г.). «Транспортер ABCG2 является эффективным оттоком Hoechst 33342 и предпочтительно экспрессируется незрелыми гематопоэтическими предшественниками человека». Кровь. 99 (2): 507–12. Дои:10.1182 / blood.V99.2.507. PMID 11781231.
- ^ Пастрана Э., Сильва-Варгас В., Дотч Ф. (май 2011 г.). «Глаза широко открыты: критический обзор сферообразования как метода определения стволовых клеток». Стволовая клетка. 8 (5): 486–98. Дои:10.1016 / j.stem.2011.04.007. ЧВК 3633588. PMID 21549325.
- ^ Николис СК (февраль 2007 г.). «Раковые стволовые клетки и гены« стволовости »в нейроонкологии». Нейробиология болезней. 25 (2): 217–29. Дои:10.1016 / j.nbd.2006.08.022. PMID 17141509.
- ^ Ядав УП, Сингх Т., Кумар П., Шарма П., Каур Х., Шарма С., Сингх С., Кумар С. и Мехта К. (2020). «Метаболические адаптации в раковых стволовых клетках». Границы онкологии (Метаболическая пластичность рака). Дои:10.3389 / fonc.2020.01010.
- ^ Ким Ю.С., Кайдина А.М., Чанг Дж.Х., Ярыгин К.Н., Лупатов А.Ю. (2017). «Молекулярные маркеры раковых стволовых клеток подтверждены in vivo». Biochem. Москва Доп. Сер. B. 11 (1): 43–54. Дои:10.1134 / S1990750817010036.
- ^ а б Менг Э., Митра А., Трипати К., Финан М.А., Скаличи Дж., Макклеллан С., Мадейра да Силва Л., Рид Е., Шевде Л.А., Палле К., Роккони Р.П. (12 сентября 2014 г.). «ALDH1A1 поддерживает свойства стволовых клеток рака яичников за счет изменения регуляции контрольной точки клеточного цикла и передачи сигналов сети репарации ДНК». PLOS ONE. 9 (9): e107142. Дои:10.1371 / journal.pone.0107142. ЧВК 4162571. PMID 25216266.
- ^ Visvader JE, Lindeman GJ (октябрь 2008 г.). «Раковые стволовые клетки в солидных опухолях: накопление доказательств и нерешенные вопросы». Обзоры природы. Рак. 8 (10): 755–68. Дои:10.1038 / nrc2499. PMID 18784658.
- ^ Аль-Хадж М., Вича М.С., Бенито-Эрнандес А., Моррисон С.Дж., Кларк М.Ф. (апрель 2003 г.). «Проспективная идентификация онкогенных клеток рака груди». Труды Национальной академии наук Соединенных Штатов Америки. 100 (7): 3983–8. Bibcode:2003ПНАС..100.3983А. Дои:10.1073 / пнас.0530291100. ЧВК 153034. PMID 12629218.
- ^ Hirschmann-Jax C, Foster AE, Wulf GG, Nuchtern JG, Jax TW, Gobel U, Goodell MA, Brenner MK (сентябрь 2004 г.). «Отчетливая« побочная популяция »клеток с высокой способностью оттока лекарственного средства в опухолевые клетки человека». Труды Национальной академии наук Соединенных Штатов Америки. 101 (39): 14228–33. Bibcode:2004PNAS..10114228H. Дои:10.1073 / pnas.0400067101. ЧВК 521140. PMID 15381773.
- ^ Ginestier C, Hur MH, Charafe-Jauffret E, Monville F, Dutcher J, Brown M, Jacquemier J, Viens P, Kleer CG, Liu S, Schott A, Hayes D, Birnbaum D, Wicha MS, Dontu G (ноябрь 2007 г.) . «ALDH1 является маркером нормальных и злокачественных стволовых клеток молочной железы человека и предиктором плохого клинического исхода». Стволовая клетка. 1 (5): 555–67. Дои:10.1016 / j.stem.2007.08.014. ЧВК 2423808. PMID 18371393.
- ^ Pece S, Tosoni D, Confalonieri S, Mazzarol G, Vecchi M, Ronzoni S, Bernard L, Viale G, Pelicci PG, Di Fiore PP (январь 2010 г.). «Биологическая и молекулярная гетерогенность рака груди коррелирует с содержанием в них раковых стволовых клеток». Клетка. 140 (1): 62–73. Дои:10.1016 / j.cell.2009.12.007. PMID 20074520.
- ^ Deng S, Yang X, Lassus H, Liang S, Kaur S, Ye Q, Li C, Wang LP, Roby KF, Orsulic S, Connolly DC, Zhang Y, Montone K, Bützow R, Coukos G, Zhang L (апрель 2010 г. ). Цао Y (ред.). «Определенные уровни экспрессии и паттерны маркера стволовых клеток, изоформы 1 альдегиддегидрогеназы (ALDH1), при раке эпителия человека». PLOS ONE. 5 (4): e10277. Bibcode:2010PLoSO ... 510277D. Дои:10.1371 / journal.pone.0010277. ЧВК 2858084. PMID 20422001.
- ^ Мейер MJ, Флеминг JM, Лин AF, Hussnain SA, Ginsburg E, Vonderhaar BK (июнь 2010 г.). «CD44posCD49fhiCD133 / 2hi определяет клетки, инициирующие ксенотрансплантат при раке молочной железы, отрицательном по рецепторам эстрогена». Исследования рака. 70 (11): 4624–33. Дои:10.1158 / 0008-5472.CAN-09-3619. ЧВК 4129519. PMID 20484027.
- ^ Сингх С.К., Хокинс С., Кларк И.Д., Сквайр Дж. А., Баяни Дж., Хиде Т., Хенкельман Р. М., Кузимано, доктор медицины, Диркс П. Б. (ноябрь 2004 г.). «Идентификация клеток-инициаторов опухоли головного мозга человека». Природа. 432 (7015): 396–401. Bibcode:2004Натура.432..396S. Дои:10.1038 / природа03128. PMID 15549107.
- ^ Сын MJ, Woolard K, Nam DH, Lee J, Fine HA (май 2009 г.). «SSEA-1 - маркер обогащения опухолевых клеток в глиобластоме человека». Стволовая клетка. 4 (5): 440–52. Дои:10.1016 / j.stem.2009.03.003. ЧВК 7227614. PMID 19427293.
- ^ а б Mazzoleni S, Politi LS, Pala M, Cominelli M, Franzin A, Sergi Sergi L., Falini A, De Palma M, Bulfone A, Poliani PL, Galli R (октябрь 2010 г.). «Экспрессия рецептора эпидермального фактора роста идентифицирует функционально и молекулярно различные инициирующие опухоль клетки в мультиформной глиобластоме человека и требуется для глиомагенеза». Исследования рака. 70 (19): 7500–13. Дои:10.1158 / 0008-5472.CAN-10-2353. PMID 20858720.
- ^ Анидо Дж., Саес-Бордериас А., Гонсалес-Хунка А., Родон Л., Фольч Г., Кармона М. А., Прието-Санчес Р. М., Барба И., Мартинес-Саес Е., Прудкин Л., Куартас I, Равентос С., Мартинес-Рикарте Ф., Пока Массачусетс, Гарсия-Дорадо Д., Лан М.М., Инлинг Дж. М., Родон Дж., Сахукильо Дж., Базельга Дж., Сеоан Дж. (Декабрь 2010 г.). «Ингибиторы рецептора TGF-β нацелены на популяцию клеток, инициирующих глиому CD44 (высокий) / Id1 (высокий уровень) в глиобластоме человека». Раковая клетка. 18 (6): 655–68. Дои:10.1016 / j.ccr.2010.10.023. PMID 21156287.
- ^ а б Далерба П., Дилла С.Дж., Пак И.К., Лю Р., Ван Икс, Чо Р.В., Хои Т., Герни А., Хуанг Э.Х., Симеоне Д.М., Шелтон А.А., Пармиани Дж., Кастелли С., Кларк М.Ф. (июнь 2007 г.). «Фенотипическая характеристика стволовых клеток колоректального рака человека». Труды Национальной академии наук Соединенных Штатов Америки. 104 (24): 10158–63. Bibcode:2007PNAS..10410158D. Дои:10.1073 / pnas.0703478104. ЧВК 1891215. PMID 17548814.
- ^ Уилсон Б.Дж., Шаттон Т., Жан К., Гассер М., Ма Дж., Сааб К.Р., Шанче Р., Ваага-Гассер А.М., Gold JS, Хуанг К., Мерфи Г.Ф., Франк М.Х., Франк Нью-Йорк (август 2011 г.). «ABCB5 идентифицирует резистентную к терапии популяцию опухолевых клеток у пациентов с колоректальным раком». Исследования рака. 71 (15): 5307–16. Дои:10.1158 / 0008-5472.CAN-11-0221. ЧВК 3395026. PMID 21652540.
- ^ Odoux C, Fohrer H, Hoppo T., Guzik L, Stolz DB, Lewis DW, Gollin SM, Gamblin TC, Geller DA, Lagasse E (сентябрь 2008 г.). «Стохастическая модель происхождения раковых стволовых клеток при метастатическом раке толстой кишки». Исследования рака. 68 (17): 6932–41. Дои:10.1158 / 0008-5472.CAN-07-5779. ЧВК 2562348. PMID 18757407.
- ^ Хуанг Э. Х., Хайнс М. Дж., Чжан Т., Гинестьер С., Донту Дж., Аппельман Х., Филдс Дж. З., Вича М. С., Боман Б. М. (апрель 2009 г.). «Альдегиддегидрогеназа 1 является маркером нормальных и злокачественных стволовых клеток (SC) толстой кишки человека и отслеживает перенаселение SC во время туморогенеза толстой кишки». Исследования рака. 69 (8): 3382–9. Дои:10.1158 / 0008-5472.CAN-08-4418. ЧВК 2789401. PMID 19336570.
- ^ Kemper K, Sprick MR, de Bree M, Scopelliti A, Vermeulen L, Hoek M, Zeilstra J, Pals ST, Mehmet H, Stassi G, Medema JP (январь 2010 г.). «Эпитоп AC133, но не белок CD133, теряется при дифференцировке раковых стволовых клеток». Исследования рака. 70 (2): 719–29. Дои:10.1158 / 0008-5472.CAN-09-1820. PMID 20068153.
- ^ Лю С., Келнар К., Лю Б., Чен Х, Калхун-Дэвис Т., Ли Х, Патравала Л., Ян Х., Джетер С., Онорио С., Уиггинс Дж. Ф., Бадер А.Г., Фэгин Р., Браун Д., Тан Д.Г. (февраль 2011 г.) . «МикроРНК miR-34a ингибирует стволовые клетки рака простаты и метастазирование путем прямого подавления CD44». Природа Медицина. 17 (2): 211–5. Дои:10,1038 / нм. 2284. ЧВК 3076220. PMID 21240262.
- ^ Хо М.М., Нг А.В., Лам С., Хунг Дж.Й. (май 2007 г.). «Побочная популяция в клеточных линиях и опухолях рака легких человека обогащена стволовыми раковыми клетками». Исследования рака. 67 (10): 4827–33. Дои:10.1158 / 0008-5472.CAN-06-3557. PMID 17510412.
- ^ Патравала Л., Калхун Т., Шнайдер-Бруссард Р., Ли Х, Бхатиа Б., Танг С., Рейли Дж. Г., Чандра Д., Чжоу Дж., Клейпул К., Коглан Л., Тан Д. Г. (март 2006 г.). «Высокоочищенные CD44 + клетки рака простаты из ксенотрансплантата опухолей человека обогащены онкогенными и метастатическими клетками-предшественниками». Онкоген. 25 (12): 1696–708. Дои:10.1038 / sj.onc.1209327. PMID 16449977.
- ^ Дубровска А., Ким С., Саламоне Р. Дж., Уокер Дж. Р., Майра С. М., Гарсия-Эчеверрия С., Шульц П. Г., Редди В. А. (январь 2009 г.). «Роль передачи сигналов PTEN / Akt / PI3K в поддержании и жизнеспособности популяций стволовых клеток рака простаты». Труды Национальной академии наук Соединенных Штатов Америки. 106 (1): 268–73. Bibcode:2009ПНАС..106..268D. Дои:10.1073 / pnas.0810956106. ЧВК 2629188. PMID 19116269.
- ^ Раджасекхар В.К., Студер Л., Джеральд В., Socci ND, Scher HI (январь 2011 г.). «Стволовые стволовые клетки, инициирующие опухоль, при раке предстательной железы человека демонстрируют повышенную передачу сигналов NF-κB». Nature Communications. 2 (1): 162. Bibcode:2011НатКо ... 2..162R. Дои:10.1038 / ncomms1159. ЧВК 3105310. PMID 21245843.
- ^ Ли Т., Су И, Мэй И, Ленг Кью, Ленг Б., Лю З., Стасс С.А., Цзян Ф. (февраль 2010 г.). «ALDH1A1 - это маркер злокачественных стволовых клеток простаты и предиктор исхода рака простаты». Лабораторные исследования; Журнал технических методов и патологии. 90 (2): 234–44. Дои:10.1038 / labinvest.2009.127. ЧВК 3552330. PMID 20010854.
- ^ Eramo A, Lotti F, Sette G, Pilozzi E, Biffoni M, Di Virgilio A, Conticello C, Ruco L, Peschle C, De Maria R (март 2008 г.). «Идентификация и расширение популяции стволовых клеток онкогенного рака легкого». Гибель клеток и дифференциация. 15 (3): 504–14. Дои:10.1038 / sj.cdd.4402283. PMID 18049477.
- ^ Салливан Дж. П., Спинола М., Додж М., Расо М. Г., Беренс К., Гао Б., Шустер К., Шао С., Ларсен Дж. , Минна JD (декабрь 2010 г.). «Активность альдегиддегидрогеназы приводит к отбору стволовых клеток аденокарциномы легких в зависимости от передачи сигналов notch». Исследования рака. 70 (23): 9937–48. Дои:10.1158 / 0008-5472.CAN-10-0881. ЧВК 3058307. PMID 21118965.
- ^ Леунг Э.Л., Фискус Р.Р., Тунг Дж.В., Тин В.П., Ченг Л.К., Сихое А.Д., Финк Л.М., Ма И, Вонг М.П. (ноябрь 2010 г.). Джин Д.Й. (ред.). «Клетки немелкоклеточного рака легких, экспрессирующие CD44, обладают свойствами, подобными стволовым клеткам». PLOS ONE. 5 (11): e14062. Bibcode:2010PLoSO ... 514062L. Дои:10.1371 / journal.pone.0014062. ЧВК 2988826. PMID 21124918.
- ^ Damelin M, Geles KG, Follettie MT, Yuan P, Baxter M, Golas J, DiJoseph JF, Karnoub M, Huang S, Diesl V, Behrens C, Choe SE, Rios C, Gruzas J, Sridharan L, Dougher M, Kunz A , Hamann PR, Evans D, Armellino D, Khandke K, Marquette K, Tchistiakova L, Boghaert ER, Abraham RT, Wistuba II, Zhou BB (июнь 2011 г.). «Определение клеточной иерархии при раке легкого позволяет выявить онкофетальный антиген, экспрессируемый на клетках, инициирующих опухоль». Исследования рака. 71 (12): 4236–46. Дои:10.1158 / 0008-5472.CAN-10-3919. PMID 21540235.
- ^ Гонсалес-Вильярреал, Калифорния, Кирос-Рейес АГ, Ислас Дж. Ф., Гарса-Тревиньо, EN (2020-08-20). «Стволовые клетки колоректального рака в развитии метастазов в печень». Границы онкологии. 10: 1511. Дои:10.3389 / fonc.2020.01511. ЧВК 7468493. PMID 32974184.
- ^ Thiery JP (июнь 2002 г.). «Эпителиально-мезенхимальные переходы при опухолевой прогрессии». Обзоры природы. Рак. 2 (6): 442–54. Дои:10.1038 / nrc822. PMID 12189386.
- ^ Ангерер Л. М., Ангерер Р. К. (июнь 1999 г.). «Регуляторное развитие эмбриона морского ежа: сигнальные каскады и градиенты морфогенов». Семинары по клеточной биологии и биологии развития. 10 (3): 327–34. Дои:10.1006 / scdb.1999.0292. PMID 10441547.
- ^ Мани С.А., Ян Дж., Брукс М., Шванингер Дж., Чжоу А., Миура Н., Куток Дж. Л., Хартвелл К., Ричардсон А. Л., Вайнберг Р. А. (июнь 2007 г.). «Мезенхима Forkhead 1 (FOXC2) играет ключевую роль в метастазировании и связана с агрессивным базальным раком молочной железы». Труды Национальной академии наук Соединенных Штатов Америки. 104 (24): 10069–74. Bibcode:2007ПНАС..10410069М. Дои:10.1073 / pnas.0703900104. ЧВК 1891217. PMID 17537911.
- ^ Коначчи-Соррелл М., Симха И., Бен-Йедидия Т., Блехман Дж., Савагнер П., Бен-Зеев А. (ноябрь 2003 г.). «Ауторегуляция экспрессии E-кадгерина посредством взаимодействий кадгерин-кадгерин: роль передачи сигналов бета-катенина, Slug и MAPK». Журнал клеточной биологии. 163 (4): 847–57. Дои:10.1083 / jcb.200308162. ЧВК 2173691. PMID 14623871.
- ^ Комито Дж., Кальвани М., Джаннони Е., Бьянкини Ф, Калорини Л., Торре Е., Мильоре С., Джордано С., Кьяруги П. (август 2011 г.). «Стабилизация HIF-1α митохондриальными АФК способствует Met-зависимому инвазивному росту и васкулогенной мимикрии в клетках меланомы». Свободная радикальная биология и медицина. 51 (4): 893–904. Дои:10.1016 / j.freeradbiomed.2011.05.042. HDL:2158/496457. PMID 21703345.
- ^ Разлив F, Рейнольдс Д.С., Камм Р.Д., Заман М.Х. (август 2016 г.). «Влияние физической микросреды на прогрессирование опухоли и метастазирование». Текущее мнение в области биотехнологии. 40: 41–48. Дои:10.1016 / j.copbio.2016.02.007. ЧВК 4975620. PMID 26938687.
- ^ Каплан Р.Н., Риба Р.Д., Захарулис С., Брамли А.Х., Винсент Л., Коста С. и др. (Декабрь 2005 г.). «VEGFR1-положительные гематопоэтические предшественники костного мозга инициируют дометастатическую нишу». Природа. 438 (7069): 820–7. Bibcode:2005Натура 438..820К. Дои:10.1038 / природа04186. ЧВК 2945882. PMID 16341007.
- ^ Гонсалес-Вильярреал, Калифорния, Кирос-Рейес АГ, Ислас Дж. Ф., Гарса-Тревиньо, EN (2020-08-20). «Стволовые клетки колоректального рака в развитии метастазов в печень». Границы онкологии. 10: 1511. Дои:10.3389 / fonc.2020.01511. ЧВК 7468493. PMID 32974184.
- ^ Hermann PC, Huber SL, Herrler T., Aicher A, Ellwart JW, Guba M, et al. (Сентябрь 2007 г.). «Различные популяции раковых стволовых клеток определяют рост опухоли и метастатическую активность при раке поджелудочной железы человека». Стволовая клетка. 1 (3): 313–23. Дои:10.1016 / j.stem.2007.06.002. PMID 18371365.
- ^ Ян З.Ф., Хо Д.В., Нг М.Н., Лау С.К., Ю.В.К., Нгаи П. и др. (Февраль 2008 г.). «Значение раковых стволовых клеток CD90 + при раке печени человека». Раковая клетка. 13 (2): 153–66. Дои:10.1016 / j.ccr.2008.01.013. PMID 18242515.
- ^ Шипицин М., Кэмпбелл Л.Л., Аргани П., Веремович С., Блуштайн-Кимрон Н., Яо Дж., Никольская Т., Серебряская Т., Бероухим Р., Ху М., Галушка М.К., Сукумар С., Паркер Л.М., Андерсон К.С., Харрис Л.Н., Гарбер Дж.Э., Ричардсон А.Л., Шнитт С.Дж., Никольский Ю., Гельман Р.С., Поляк К. (март 2007 г.). «Молекулярное определение неоднородности опухоли груди». Раковая клетка. 11 (3): 259–73. Дои:10.1016 / j.ccr.2007.01.013. PMID 17349583.
- ^ Шмелков С.В., Батлер Дж. М., Хупер А. Т., Хормиго А., Кушнер Дж., Милде Т., Сент-Клер Р., Бальевич М., Уайт И., Джин Д. К., Чадберн А., Мерфи А. Дж., Валенсуэла Д. М., Гейл Н. В., Терстон Г., Янкопулос Г. Д., Д. Анжелика М., Кемени Н., Лайден Д., Рафии С. (июнь 2008 г.). «Экспрессия CD133 не ограничивается стволовыми клетками, и как CD133 +, так и CD133- метастатические клетки рака толстой кишки инициируют опухоли». Журнал клинических исследований. 118 (6): 2111–20. Дои:10.1172 / JCI34401. ЧВК 2391278. PMID 18497886.
- ^ Brabletz T, Jung A, Spaderna S, Hlubek F, Kirchner T (сентябрь 2005 г.). «Мнение: мигрирующие раковые стволовые клетки - комплексная концепция прогрессирования злокачественной опухоли». Обзоры природы. Рак. 5 (9): 744–9. Дои:10.1038 / nrc1694. PMID 16148886.
- ^ Чжао Дж (2016). «Раковые стволовые клетки и химиорезистентность: самые умные выживают в рейде». Pharmacol Ther. 160: 145–58. Дои:10.1016 / j.pharmthera.2016.02.008. ЧВК 4808328. PMID 26899500.
- ^ Диркс П. (июль 2010 г.). «Раковые стволовые клетки: приглашение на второй тур». Природа. 466 (7302): 40–1. Bibcode:2010Натура.466 ... 40Д. Дои:10.1038 / 466040a. PMID 20596007.
- ^ Хаскинс В.Е., Эдала С., Джадхав Ю.Л., Лабхан М.С., Перичерла В.К., Перлман Э.Дж. (май 2012 г.). «Понимание неопластических стволовых клеток на основе протеомики гелевых опухолей детских половых клеток». Детская кровь и рак. 58 (5): 722–8. Дои:10.1002 / pbc.23282. ЧВК 3204330. PMID 21793190.
- ^ Гупта П.Б., Ондер Т.Т., Цзян Г., Тао К., Купервассер К., Вайнберг Р.А., Лендер Э.С. (август 2009 г.). «Идентификация селективных ингибиторов раковых стволовых клеток с помощью высокопроизводительного скрининга». Клетка. 138 (4): 645–659. Дои:10.1016 / j.cell.2009.06.034. ЧВК 4892125. PMID 19682730.
- ^ Джангамредди Дж. Р., Гавами С., Грабарек Дж., Кратц Дж., Вичек Е., Фредрикссон Б. А., Рао Парити Р. К., Цеслар-Побуда А., Паниграхи С., Ос М. Дж. (Сентябрь 2013 г.). «Салиномицин вызывает активацию аутофагии, митофагии и влияет на полярность митохондрий: различия между первичными и раковыми клетками». Biochimica et Biophysica Acta (BBA) - Исследование молекулярных клеток. 1833 (9): 2057–69. Дои:10.1016 / j.bbamcr.2013.04.011. PMID 23639289.
- ^ Vlahopoulos S, Critselis E, Voutsas IF, Perez SA, Moschovi M, Baxevanis CN, Chrousos GP (2014). «Новое применение старых препаратов? Предполагаемые цели хлорохинов в терапии рака». Текущие цели в отношении лекарств. 15 (9): 843–51. Дои:10.2174/1389450115666140714121514. PMID 25023646.
- ^ Jordan CT, Upchurch D, Szilvassy SJ, Guzman ML, Howard DS, Pettigrew AL, Meyerrose T, Rossi R, Grimes B, Rizzieri DA, Luger SM, Phillips GL (октябрь 2000 г.). «Альфа-цепь рецептора интерлейкина-3 является уникальным маркером стволовых клеток острого миелогенного лейкоза человека». Лейкемия. 14 (10): 1777–84. Дои:10.1038 / sj.leu.2401903. PMID 11021753.
- ^ Джин Л., Ли Э.М., Рамшоу Х.С., Басфилд С.Дж., Peoppl AG, Уилкинсон Л., Гатридж М.А., Томас Д., Барри Э.Ф., Бойд А., Геринг Д.П., Вайро Г., Лопес А.Ф., Дик Дж. Э., Лок РБ (июль 2009 г.). «Опосредованное моноклональными антителами нацеливание на CD123, альфа-цепь рецептора IL-3, устраняет человеческие стволовые клетки острого миелоидного лейкоза». Стволовая клетка. 5 (1): 31–42. Дои:10.1016 / j.stem.2009.04.018. PMID 19570512.
- ^ "'Нанобомбы, взрывающие раковые клетки | KurzweilAI ". www.kurzweilai.net. 2015-12-07. Получено 2016-02-20.
- ^ Ван Х, Агарвал П., Чжао С., Ю Дж, Лу Х, Хе Х (январь 2016 г.). "Нанобомба, активируемая лазером в ближнем инфракрасном диапазоне, для преодоления барьеров на пути доставки микроРНК". Современные материалы. 28 (2): 347–55. Дои:10.1002 / adma.201504263. PMID 26567892.
- ^ Huddle BC, Grimley E, Buchman CD, Chtcherbinine M, Debnath B, Mehta P, Yang K, Morgan CA, Li S, Felton JA, Sun D, Metha G, Neamati N, Buckanovich RJ, Hurley TD, Larsen SD (2018) . «Оптимизация на основе структуры нового класса селективных ингибиторов подсемейства альдегиддегидрогеназы 1А (ALDH1A) в качестве потенциальных дополнений к химиотерапии рака яичников». J Med Chem. 61 (19): 8754–8773. Дои:10.1021 / acs.jmedchem.8b00930. ЧВК 6477540. PMID 30221940.
- ^ Haupt Y, Bath ML, Harris AW, Adams JM (ноябрь 1993 г.). «Трансген bmi-1 индуцирует лимфомы и взаимодействует с myc в онкогенезе». Онкоген. 8 (11): 3161–4. PMID 8414519.
- ^ Пак И.К., Цянь Д., Киль М., Беккер М.В., Пихаля М., Вайсман И.Л., Моррисон С.Дж., Кларк М.Ф. (май 2003 г.). «Bmi-1 необходим для поддержания самообновляющихся гемопоэтических стволовых клеток взрослых» (PDF). Природа. 423 (6937): 302–5. Bibcode:2003Натура.423..302П. Дои:10.1038 / природа01587. HDL:2027.42/62508. PMID 12714971.
- ^ Молофски А.В., Пардал Р., Ивашита Т., Парк И.К., Кларк М.Ф., Моррисон С.Дж. (октябрь 2003 г.). «Зависимость от Bmi-1 отличает самообновление нервных стволовых клеток от пролиферации предшественников». Природа. 425 (6961): 962–7. Bibcode:2003Натура.425..962М. Дои:10.1038 / природа02060. ЧВК 2614897. PMID 14574365.
- ^ Hemmati HD, Nakano I, Lazareff JA, Masterman-Smith M, Geschwind DH, Bronner-Fraser M, Kornblum HI (декабрь 2003 г.). «Раковые стволовые клетки могут возникать из детских опухолей головного мозга». Труды Национальной академии наук Соединенных Штатов Америки. 100 (25): 15178–83. Bibcode:2003ПНАС..10015178Н. Дои:10.1073 / pnas.2036535100. ЧВК 299944. PMID 14645703.
- ^ Донту Г., Джексон К.В., МакНиколас Э., Кавамура М.Дж., Абдалла В.М., Вича М.С. (2004). «Роль передачи сигналов Notch в определении судьбы стволовых клеток / клеток-предшественников молочной железы человека». Исследование рака груди. 6 (6): R605–15. Дои:10.1186 / bcr920. ЧВК 1064073. PMID 15535842.
- ^ Diévart A, Beaulieu N, Jolicoeur P (октябрь 1999 г.). «Участие Notch1 в развитии опухолей молочной железы мышей». Онкоген. 18 (44): 5973–81. Дои:10.1038 / sj.onc.1202991. PMID 10557086.
- ^ Парк Д.М., Юнг Дж., Масджкур Дж., Макрогкикас С., Эберманн Д., Саха С., Роглиано Р., Паолилло Н., Пачони С., Маккей Р. Д., Позер С., Андруцеллис-Теотокис А. (2013). «Hes3 регулирует количество клеток в культурах мультиформной глиобластомы с характеристиками стволовых клеток». Научные отчеты. 3: 1095. Bibcode:2013НатСР ... 3Э1095П. Дои:10.1038 / srep01095. ЧВК 3566603. PMID 23393614.
- ^ Милошевич В. и др. Аутокринные цепи Wnt / IL-1β / IL-8 контролируют химиорезистентность в инициирующих клетках мезотелиомы, индуцируя ABCB5.Int. J. Рак, https://doi.org/10.1002/ijc.32419
- ^ Бичи PA, Karhadkar SS, Berman DM (ноябрь 2004 г.). «Ремонт тканей и обновление стволовых клеток в канцерогенезе». Природа. 432 (7015): 324–31. Bibcode:2004Натура.432..324Б. Дои:10.1038 / природа03100. PMID 15549094.
- ^ Чжоу Б.П., Хунг МС (июнь 2005 г.). «Wnt, hedgehog и snail: сестринские пути, которые контролируют GSK-3beta и beta-Trcp в регуляции метастазирования». Клеточный цикл. 4 (6): 772–6. Дои:10.4161 / cc.4.6.1744. PMID 15917668.
- ^ Акиёси Т., Накамура М., Кога К., Накашима Х., Яо Т., Цунэёси М., Танака М., Катано М. (июль 2006 г.). «Gli1, подавленный при колоректальном раке, подавляет пролиферацию клеток рака толстой кишки, включая активацию передачи сигналов Wnt». Кишечник. 55 (7): 991–9. Дои:10.1136 / gut.2005.080333. ЧВК 1856354. PMID 16299030.
- ^ She M, Chen X (январь 2009 г.). «Нацеливание на сигнальные пути, активные в раковых стволовых клетках, для преодоления устойчивости к лекарствам». Чжунго Фэй AI За Чжи = Китайский журнал рака легких. 12 (1): 3–7. Дои:10.3779 / j.issn.1009-3419.2009.01.001. PMID 20712949.
- ^ Боллманн FM (август 2008 г.). «Многоликая теломераза: новые экстрарателомерные эффекты». BioEssays. 30 (8): 728–32. Дои:10.1002 / bies.20793. PMID 18623070.
- ^ Hodge R (25 января 2016 г.). «Взлом программ раковых стволовых клеток». medicalxpress.com. Медицинский Экспресс. Получено 2016-02-12.
дальнейшее чтение
- Раджасекхар В.К., изд. (2014). Раковые стволовые клетки. Хобокен, Нью-Джерси: Wiley. ISBN 9781118356166.
- Милошевич V и др. (Январь 2020 г.). «Аутокринные цепи Wnt / IL-1β / IL-8 контролируют химиорезистентность в инициирующих клетках мезотелиомы, индуцируя ABCB5». Int. J. Рак. 146 (1): 192–207. Дои:10.1002 / ijc.32419.
| Источники / типы | |
|---|---|
| Клеточная активность | |
| Статьи по Теме | |
| |