Иммунопреципитация хроматина - Chromatin immunoprecipitation
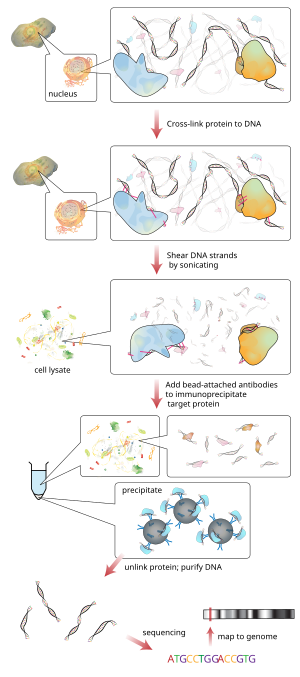
Иммунопреципитация хроматина (ЧИП) является разновидностью иммунопреципитация экспериментальная техника, используемая для исследования взаимодействия между белки и ДНК в камере. Он направлен на определение того, связаны ли определенные белки с конкретными областями генома, такими как факторы транскрипции на промоутеры или другой Сайты связывания ДНК, и, возможно, определяя цистромы. ChIP также направлен на определение конкретного места в геноме, которое различные гистон модификации связаны с, указывая на цель модификаторов гистонов.[1]
Вкратце, обычный метод выглядит следующим образом:
- ДНК и ассоциированные белки на хроматин в живых клетках или тканях сшиваются (этот шаг опущен в Native ChIP).
- Комплексы ДНК-белок (хроматин-белок) затем разрезаются на фрагменты ДНК размером ~ 500 п.н. обработка ультразвуком или расщепление нуклеазой.
- Сшитый Фрагменты ДНК, связанные с представляющим интерес белком (белками), избирательно иммунопреципитируют из клеточного дебриса с использованием подходящего специфичного для белка антитела.
- Связанные фрагменты ДНК очищаются и определяется их последовательность. Обогащение конкретных последовательностей ДНК представляет области в геноме, с которыми связан интересующий белок. in vivo.
Типичный чип
В основном существует два типа ChIP, в первую очередь различающиеся исходным препаратом хроматина. Первый использует обратимо сшитый хроматин, разрезанный обработка ультразвуком называется сшитым ЧИП (ХЧИП). Native ChIP (NChIP) использует нативный хроматин, разрезанный микрококковый нуклеаза пищеварение.[нужна цитата ]
Сшитый чип (XChIP)
Сшитый ChIP в основном подходит для картирования ДНК-мишени факторов транскрипции или других белков, связанных с хроматином, и используется обратимо. сшитый хроматин в качестве исходного материала. Агент для обратимого сшивания может быть формальдегид[2] или же УФ-излучение.[3] Затем поперечно сшитый хроматин обычно разрезается ультразвуком, получая фрагменты размером 300 - 1000 пар оснований (п.н.) в длину. Мягкое поперечное сшивание формальдегидом с последующим расщеплением нуклеазами было использовано для сдвига хроматина.[4] Фрагменты хроматина размером 400-500 п.н. оказались подходящими для анализов ChIP, поскольку они охватывают от двух до трех нуклеосомы.
Клеточный дебрис в разрезанном лизате затем очищается путем осаждения, и комплексы белок-ДНК избирательно иммунопреципитируются с использованием специфических антитела к интересующим белкам. Антитела обычно связаны с агароза, сефароза или магнитные бусины. Альтернативно, комплексы хроматин-антитело можно селективно удерживать и элюировать дисками из инертного полимера.[5][6] Иммунопреципитированные комплексы (т. Е. Комплекс бусина-антитело-белок-последовательность ДНК-мишени) затем собирают и промывают для удаления неспецифически связанного хроматина, белок-ДНК. перекрестная ссылка отменяется, и белки удаляются перевариванием с протеиназа К. An эпитоп -помеченная версия интересующего белка, или in vivo биотинилирование [7] могут использоваться вместо антител к интересующему природному белку.
Затем ДНК, связанная с комплексом, очищается и идентифицируется с помощью полимеразной цепной реакции (ПЦР), микрочипы (Чип-на-чипе ), молекулярное клонирование и секвенирование или прямое высокопроизводительное секвенирование (ChIP-Seq ).[нужна цитата ]
Родной ЧИП (НЧИП)
Native ChIP в основном подходит для картирования ДНК-мишени гистон модификаторы. Обычно в качестве исходного хроматина используется нативный хроматин. Поскольку гистоны обвивают ДНК с образованием нуклеосом, они естественным образом связаны. Затем хроматин расщепляется микрококковой нуклеазой, которая разрезает ДНК по длине линкера, оставляя нуклеосомы нетронутыми и обеспечивая фрагменты ДНК длиной от одной нуклеосомы (200 пар оснований) до пяти нуклеосом (1000 пар оснований). После этого используются методы, аналогичные XChIP. для очистки клеточного дебриса, иммунопреципитации интересующего белка, удаления белка из иммунопреципитированного комплекса и очистки и анализа ДНК, ассоциированной с комплексом.[нужна цитата ]
Сравнение ХЧИП и НЧИП
Основное преимущество НЧИП: антитело специфичность. Важно отметить, что большинство антител к модифицированным гистонам вырабатываются против нефиксированных синтетических пептидных антигенов и что эпитопы они должны распознать в XChIP могут быть нарушены или разрушены формальдегидом сшивание, особенно как перекрестные ссылки вероятно, будет включать лизин е-аминогруппы на N-концах, разрушающие эпитопы. Это, вероятно, объясняет неизменно низкую эффективность протоколов XChIP по сравнению с NChIP.
Но у ХЧИП и НЧИП разные цели и преимущества по отношению друг к другу. XChIP предназначен для картирования сайтов-мишеней факторов транскрипции и других белков, связанных с хроматином; NChIP предназначен для картирования целевых сайтов модификаторов гистонов (см. Таблицу 1).
Таблица 1 Преимущества и недостатки НЧИП и ХЧИП
| XChIP | НЧИП | |
|---|---|---|
| Преимущества | Подходит для факторов транскрипции или любых других слабо связывающихся белков, связанных с хроматином. Применимо к любым организмам, у которых сложно приготовить нативный белок | Тестируемая специфичность антител Лучшая специфичность антител в качестве целевого белка в естественном состоянии Лучшая эффективность восстановления хроматина и белка за счет лучшей специфичности антител |
| Недостатки | Неэффективное восстановление хроматина из-за разрушения эпитопа белка-мишени антитела Может вызвать ложноположительный результат из-за фиксации временных белков на хроматине. Широкий диапазон размеров хроматина за счет случайного разрезания ультразвуком. | Обычно не подходит для негистоновых белков. Нуклеосомы могут перестраиваться во время пищеварения |
История и новые методы ChIP
В 1984 г. Джон Т. Лис и Дэвид Гилмор, в то время аспирант в лаборатории Лиса, использовали УФ-облучение, сшивающий агент нулевой длины белок-нуклеиновая кислота, чтобы ковалентно перекрестная ссылка белки, связанные с ДНК в живых бактериальных клетках. После лизиса поперечно-сшитых клеток и иммунопреципитации бактериальной РНК-полимеразы ДНК, ассоциированная с обогащенной РНК-полимеразой, гибридизовали с зондами, соответствующими различным областям известных генов, для определения распределения in vivo и плотности РНК-полимеразы в этих генах. Год спустя они использовали ту же методологию для изучения распространения эукариотических РНК-полимераза II на гены теплового шока плодовой мухи. Эти отчеты считаются пионерскими исследованиями в области иммунопреципитации хроматина.[8][9] XChIP был дополнительно модифицирован и разработан Александр Варшавский и коллеги, которые изучили распределение гистон H4 на гены теплового шока с использованием формальдегидного сшивания.[10][11] В дальнейшем этот метод получил широкое развитие и усовершенствование.[12]Подход NChIP был впервые описан Hebbes и другие., 1988,[13] а также были быстро разработаны и усовершенствованы.[14] Типичный анализ ChIP обычно занимает 4–5 дней и требует 106~ 107 клетки по крайней мере. Теперь новые методы на ChIP могут быть реализованы всего в 100 ~ 1000 ячеек и завершены в течение одного дня.
- ЧИП без шариков: В этом новом методе ChIP используются диски из инертного пористого полимера, функционализированные протеином A или G, в спин-колонках или микропланшетах. Комплекс хроматин-антитело селективно удерживается диском и элюируется для получения обогащенной ДНК для последующих применений, таких как количественная ПЦР и секвенирование. Пористая среда специально разработана для максимального увеличения эффективности захвата и уменьшения неспецифического связывания. Благодаря меньшему количеству ручной обработки и оптимизированным протоколам ChIP может быть выполнен за 5 часов.[6]
- Несущий чип (CChIP): этот подход может использовать всего 100 ячеек путем добавления Дрозофила клетки в качестве хроматина-носителя для уменьшения потерь и облегчения осаждения целевого хроматина. Однако для этого требуются высокоспецифичные праймеры для обнаружения хроматина клетки-мишени на фоне хроматина чужеродного носителя, и это занимает от двух до трех дней.[15]
- Быстрый чип (qChIP): Быстрый анализ ChIP сократил время за счет сокращения двух этапов в типичном анализе ChIP: (я) ультразвуковая ванна увеличивает скорость связывания антител с белками-мишенями и тем самым сокращает время иммунопреципитации (ii) процедура выделения ДНК на основе смолы (Chelex-100) сокращает время перекрестная ссылка обращение и выделение ДНК. Однако быстрый протокол подходит только для больших образцов клеток (в диапазоне 106~107).[16][17] Можно обработать до 24 образцов хроматина, подвергнутых сдвигу, для получения ДНК, готовой к ПЦР, за 5 часов, что позволяет одновременно исследовать несколько факторов хроматина и / или изучать геномные события в нескольких временных точках.[18]
- Быстрый и количественный чип (Q2ChIP): анализ использует 100000 клеток в качестве исходного материала и подходит для до 1000 гистоновых ChIP или 100 ChIP факторов транскрипции. Таким образом, многие образцы хроматина могут быть приготовлены параллельно и сохранены, а Q2ЧИП можно провести за день.[19]
- MicroChIP (µChIP): хроматин обычно получают из 1000 клеток, и до 8 ChIP можно делать параллельно без носителей. Анализ также может начинаться со 100 клеток, но подходит только для одного чипа. Также можно использовать небольшие (1 мм3) ткань биопсия и microChIP можно сделать за один день.[20][21]
- Матричный чип: Это микропланшет анализ на основе ChIP с повышенной производительностью и упрощением процедуры. Все этапы выполняются в лунках микропланшета без переноса образцов, что дает возможность автоматизации. Он позволяет проводить 96 анализов ChIP на гистоны и различные ДНК-связанные белки за один день.[22]
- Патология-ЧИП (PAT-ChIP): этот метод позволяет ChIP из фиксированных формалином и залитых парафином патологий тканей и, таким образом, использовать архивы патологий (даже тех, которым несколько лет) для эпигенетического анализа и идентификации возможных эпигенетических биомаркеров или мишеней.[23]
ChIP также применялся для анализа всего генома в сочетании с технологией микрочипов (Чип-на-чипе ) или технологии ДНК-секвенирования второго поколения (Чип-секвенирование ). ЧИП также может сочетаться с парные конечные теги последовательность в Анализ взаимодействия хроматина с использованием секвенирования парных конечных тегов (ChIA-PET), методика, разработанная для крупномасштабного анализа de novo структур хроматина более высокого порядка.[24][25][26]
Ограничения
- Крупномасштабные анализы с использованием ChIP затруднительны с использованием интактных модельных организмов. Это связано с тем, что для каждого TF должны быть созданы антитела или, альтернативно, необходимо получить трансгенные модельные организмы, экспрессирующие TF, меченные эпитопом.
- Исследователи, изучающие различные паттерны экспрессии генов у мелких организмов, также сталкиваются с проблемами, поскольку гены экспрессируются на низком уровне, в небольшом количестве клеток, в узком временном окне.
- ChIP эксперименты не могут различать разные изоформы TF (Изоформа белка ).
Смотрите также
- ЧИП-экзо, метод, который добавляет обработку экзонуклеазой к процессу ChIP для получения разрешения сайтов связывания до одной пары оснований
- Чип-на-чипе, сочетает в себе ChIP с технологией микрочипов
- DamID, альтернативный метод картирования местоположения, не требующий специфических антител
- RIP-чип, аналогичный метод анализа взаимодействий РНК-белок
Рекомендации
- ^ Коллас, Филипп. (Январь 2010 г.). «Современное состояние иммунопреципитации хроматина». Молекулярная биотехнология. 45 (1): 87–100. Дои:10.1007 / s12033-009-9239-8. PMID 20077036. S2CID 24225210.
- ^ Джексон, Вон (ноябрь 1978 г.). «Исследования по организации гистонов в нуклеосомах с использованием формальдегида в качестве обратимого сшивающего агента». Клетка. 15 (3): 945–54. Дои:10.1016/0092-8674(78)90278-7. PMID 569554. S2CID 25169609.
- ^ Гилмор Д.С., Лис Д.Т. (август 1985 г.). "Взаимодействие in vivo РНК-полимеразы II с генами Drosophila melanogaster". Молекулярная и клеточная биология. 5 (8): 2009–18. Дои:10.1128 / mcb.5.8.2009. ЧВК 366919. PMID 3018544.
- ^ Bauer UM, Daujat S, Nielsen SJ, Nightingale K, Kouzarides T (январь 2002 г.). «Метилирование аргинина 17 гистона H3 связано с активацией гена». EMBO отчеты. 3 (1): 39–44. Дои:10.1093 / embo-reports / kvf013. ЧВК 1083932. PMID 11751582.
- ^ Бейнон, Эми Л .; Паркс, Линдси Дж .; Тернер, Мэтью Л .; Рыцарь, Стив; Конлан, Стив; Фрэнсис, Льюис; Акции, Бен (сентябрь 2014 г.). «Chromatrap® 96: новая твердотельная платформа для высокопроизводительных чипов». Методы природы. 11 (9): i – ii. Дои:10.1038 / nmeth.f.372. ISSN 1548-7091.
- ^ а б «Хроматрап».Революционная твердотельная платформа для иммунопреципитации хроматина.
- ^ Viens A; и другие. (2004). «Использование биотинилирования белков in vivo для иммунопреципитации хроматина». Аналитическая биохимия. 325 (1): 68–76. Дои:10.1016 / j.ab.2003.10.015. PMID 14715286.
- ^ Гилмор Д.С., Лис Дж. Т. (1984). «Обнаружение взаимодействий белок-ДНК in vivo: распределение РНК-полимеразы по конкретным бактериальным генам». Proc Natl Acad Sci U S A. 81 (14): 4275–9. Bibcode:1984PNAS ... 81.4275G. Дои:10.1073 / пнас.81.14.4275. ЧВК 345570. PMID 6379641.
- ^ Гилмор Д.С., Лис Д.Т. (август 1985 г.). «Взаимодействие in vivo РНК-полимеразы II с генами Drosophila melanogaster». Мол. Клетка. Биол. 5 (8): 2009–18. Дои:10.1128 / mcb.5.8.2009. ЧВК 366919. PMID 3018544.
- ^ Варшавский А (декабрь 2008 г.). «Открытие клеточной регуляции деградацией белка». Журнал биологической химии. 283 (50): 34469–89. Дои:10.1074 / jbc.X800009200. ЧВК 3259866. PMID 18708349.
- ^ Соломон, Марк Дж; Ларсен Памела Л; Варшавский, Александр. (Июнь 1988 г.). «Картирование взаимодействий белок-ДНК in vivo с формальдегидом: доказательство того, что гистон H4 сохраняется на высокотранскрибируемом гене». Клетка. 53 (6): 937–47. Дои:10.1016 / S0092-8674 (88) 90469-2. PMID 2454748. S2CID 11169130.
- ^ Орландо V (март 2000 г.). «Картирование хромосомных белков in vivo с помощью иммунопреципитации сшитого формальдегидом хроматина». Тенденции в биохимических науках. 25 (3): 99–104. Дои:10.1016 / S0968-0004 (99) 01535-2. PMID 10694875.
- ^ Hebbes, Tim R; Торн, Алан В.; Крэйн-Робинсон С. (май 1988 г.). «Прямая связь между ацетилированием гистона ядра и транскрипционно активным хроматином». Журнал EMBO. 7 (5): 1395–402. Дои:10.1002 / j.1460-2075.1988.tb02956.x. ЧВК 458389. PMID 3409869.
- ^ О'Нил, Лаура П.; Тернер, Брайан М (сентябрь 2003 г.). «Иммунопреципитация нативного хроматина: НЧИП». Методы. 31 (1): 76–82. Дои:10.1016 / S1046-2023 (03) 00090-2. PMID 12893176.
- ^ О'Нил, Лаура П.; VerMilyea, Matthew D; Тернер, Брайан М (июль 2006 г.). «Эпигенетическая характеристика ранних эмбрионов с протоколом иммунопреципитации хроматина, применимым к небольшим популяциям клеток». Природа Генетика. 38 (7): 835–41. Дои:10,1038 / нг1820. PMID 16767102. S2CID 28311996.
- ^ Нельсон, Джоэл Д.; Денисенко, Олег; Сова, Павел; Бомштык, Кароль (2006). «Быстрый анализ иммунопреципитации хроматина». Исследования нуклеиновых кислот. 34 (1): e2. Дои:10.1093 / nar / gnj004. ЧВК 1325209. PMID 16397291.
- ^ Нельсон, Джоэл Д.; Денисенко, Олег; Бомштык, Кароль (2006). «Протокол метода быстрой иммунопреципитации хроматина (ChIP)». Протоколы природы. 1 (1): 179–85. Дои:10.1038 / nprot.2006.27. PMID 17406230. S2CID 20577722.
- ^ Нельсон Дж, Денисенко О, Бомштык К (2009). Метод быстрой иммунопреципитации хроматина. Методы молекулярной биологии. 567. С. 45–57. Дои:10.1007/978-1-60327-414-2_3. ISBN 978-1-60327-413-5. PMID 19588084.
- ^ Даль, Джон Арн; Коллас, Филипп (апрель 2007 г.). "Q2ChIP, быстрый и количественный анализ иммунопреципитации хроматина, раскрывает эпигенетическую динамику генов, регулируемых развитием, в клетках карциномы человека ». Стволовые клетки. 25 (4): 1037–46. Дои:10.1634 / стволовые клетки.2006-0430. PMID 17272500.
- ^ Даль, Джон Арн; Коллас, Филипп (2008). «Быстрый анализ иммунопреципитации микрохроматина (microChIP)». Протоколы природы. 3 (6): 1032–45. Дои:10.1038 / nprot.2008.68. PMID 18536650. S2CID 29529307.
- ^ Даль, Джон Арн; Коллас, Филипп (2009). MicroChIP: иммунопреципитация хроматина для небольшого числа клеток. Методы молекулярной биологии. 567. С. 59–74. Дои:10.1007/978-1-60327-414-2_4. ISBN 978-1-60327-413-5. PMID 19588085.
- ^ Фланагин, Стив; Нельсон, Джоэл Д.; Кастнер, Дэвид Дж; Денисенко, Олег; Бомштык, Кароль (февраль 2008 г.). «Метод иммунопреципитации хроматина на микропланшетах, Matrix ChIP: платформа для изучения передачи сигналов сложных геномных событий». Исследования нуклеиновых кислот. 36 (3): e17. Дои:10.1093 / nar / gkn001. ЧВК 2241906. PMID 18203739.
- ^ Fanelli, Mirco; Аматори, Стефано; Бароцци, Ирос; Сончини, Матиас; Зуффо, Роберто Даль; Буччи, Габриэле; Капра, Мария; Куарто, Микаэла; Деллино, Гаэтано Иван (14 декабря 2010 г.). «Патология иммунопреципитации ткани и хроматина в сочетании с высокопроизводительным секвенированием позволяет эпигенетическому профилированию образцов пациентов». Труды Национальной академии наук. 107 (50): 21535–21540. Bibcode:2010PNAS..10721535F. Дои:10.1073 / pnas.1007647107. ISSN 0027-8424. ЧВК 3003125. PMID 21106756.
- ^ Фуллвуд, Мелисса Дж; Хан, Юйюань; Вэй, Чиа-Линь; Жуань, Сяоань; Жуань, Ицзюнь (январь 2010 г.). Анализ взаимодействия хроматина с использованием секвенирования парных концевых меток. Текущие протоколы в молекулярной биологии. Глава 21. С. Раздел 21.15.1–25. Дои:10.1002 / 0471142727.mb2115s89. ISBN 978-0471142720. ЧВК 6924956. PMID 20069536.
- ^ Ли, Гуолян; Фуллвуд, Мелисса Дж; Сюй, Хань; Мулавади, Фабиан Хендриян; Велков, Стоян; Вега, Винсенсиус; Арияратне, Прамила Нуванта; Мохамед, Юсофф Бин; Оои, Хонг-Сайн; Теннакун, Чандана; Вэй, Чиа-Линь; Жуань, Ицзюнь; Сун, Вин-Кин (февраль 2010 г.). «Инструмент ChIA-PET для комплексного анализа взаимодействия хроматина с секвенированием парных концевых меток». Геномная биология. 11 (2): R22. Дои:10.1186 / gb-2010-11-2-r22. ЧВК 2872882. PMID 20181287.
- ^ "ChIA-PET: новый метод исследования трехмерного картирования всего генома". ScienceDaily. Агентство науки, технологий и исследований (A * STAR), Сингапур. 2009-11-08. Получено 14 марта 2010.
внешняя ссылка
- Хроматин + иммунопреципитация в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- EpigenomeNOE.com
- Иммунопрециптация хроматина (ChIP) на нефиксированном хроматине из клеток и тканей для анализа модификаций гистонов
- Иммунопреципитация хроматина (ChIP) белковых комплексов: картирование геномных мишеней ядерных белков в культивируемых клетках