Эксперимент Мезельсона – Шталя - Meselson–Stahl experiment
В Эксперимент Мезельсона – Шталя это эксперимент Мэтью Мезельсон и Франклин Шталь в 1958 г. Watson и Крик гипотеза о том, что Репликация ДНК был полуконсервативный. При полуконсервативной репликации, когда реплицируется двухцепочечная спираль ДНК, каждая из двух новых двухцепочечных ДНК спирали состояли из одной цепи исходной спирали и одной вновь синтезированной. Его назвали «самым красивым экспериментом в биологии».[1] Мезельсон и Шталь решили, что лучший способ пометить родительскую ДНК - это изменить один из атомов в родительской молекуле ДНК. Поскольку азот содержится в азотистых основаниях каждого нуклеотида, они решили использовать изотоп азота, чтобы различать родительскую и вновь скопированную ДНК. Изотоп азота имел дополнительный нейтрон в ядре, что сделало его тяжелее.
Гипотеза
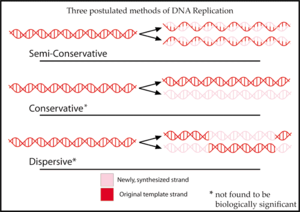
Для метода репликации ДНК ранее были предложены три гипотезы.
в полуконсервативный гипотеза, предложенная Watson и Крик, две цепи молекулы ДНК разделяются во время репликации. Каждая нить затем действует как шаблон для синтеза новой нити.[2]
В консервативный Гипотеза предполагала, что вся молекула ДНК действовала как матрица для синтеза совершенно новой. Согласно этой модели, гистон белки связываются с ДНК, вращая цепь и открывая нуклеотидные основания (которые обычно выстилают внутреннюю часть) для образования водородных связей.[3]
В диспергирующий гипотеза иллюстрируется моделью, предложенной Макс Дельбрюк, который пытается решить проблему раскручивания двух цепей двойной спирали с помощью механизма, который разрывает основную цепь ДНК каждые 10 нуклеотидов или около того, раскручивает молекулу и прикрепляет старую цепь к концу вновь синтезированной. Это позволит синтезировать ДНК короткими фрагментами, чередующимися от одной цепи к другой.[4]
Каждая из этих трех моделей дает разные прогнозы о распределении «старой» ДНК в молекулах, образовавшихся после репликации. Согласно консервативной гипотезе, после репликации одна молекула является полностью законсервированной «старой» молекулой, а другая - полностью синтезированной ДНК. Полуконсервативная гипотеза предсказывает, что каждая молекула после репликации будет содержать одну старую и одну новую цепь. Дисперсионная модель предсказывает, что каждая нить каждой новой молекулы будет содержать смесь старой и новой ДНК.[5]
Методика эксперимента и результаты
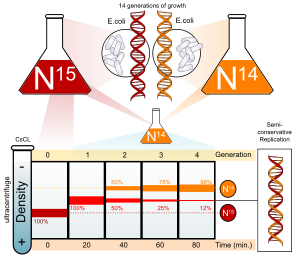
Азот является одним из основных компонентов ДНК. 14N безусловно самый распространенный изотоп азота, но ДНК с более тяжелым (но нерадиоактивным) 15N изотоп также функционален.
Кишечная палочка выращивали в течение нескольких поколений на среде, содержащей NH4Cl с 15N. Когда ДНК извлекается из этих клеток и центрифугируется на соли (CsCl ) градиент плотности ДНК отделяется в точке, в которой ее плотность равна плотности раствора соли. ДНК клеток, выращенных в 15Среда N имела более высокую плотность, чем клетки, выращенные в нормальных условиях. 14N средний. После того, Кишечная палочка ячейки только с 15N в их ДНК были перенесены в 14N средний и дали возможность разделиться; за ходом деления клеток следили с помощью микроскопического подсчета клеток и анализа колоний.
ДНК извлекалась периодически и сравнивалась с чистой 14N ДНК и 15N ДНК. После одной репликации было обнаружено, что ДНК имеет промежуточную плотность. Поскольку консервативная репликация привела бы к равному количеству ДНК более высокой и более низкой плотности (но не ДНК промежуточной плотности), консервативная репликация была исключена. Однако этот результат соответствовал как полуконсервативной, так и дисперсионной репликации. Полуконсервативная репликация приведет к получению двухцепочечной ДНК с одной цепью 15N ДНК, и одна из 14N ДНК, в то время как дисперсионная репликация привела бы к двухцепочечной ДНК с обеими цепями, имеющими смеси 15N и 14N ДНК, любая из которых могла бы появиться как ДНК промежуточной плотности.
Авторы продолжали отбирать образцы клеток по мере продолжения репликации. ДНК из клеток после завершения двух репликаций, как было обнаружено, состоит из равных количеств ДНК с двумя разными плотностями, одна из которых соответствует промежуточной плотности ДНК клеток, выращенных только за одно деление в 14N, другой соответствует ДНК из клеток, выращенных исключительно в 14N средний. Это несовместимо с дисперсионной репликацией, которая привела бы к единственной плотности, ниже, чем промежуточная плотность клеток одного поколения, но все же выше, чем у клеток, выращенных только в 14Среда N ДНК, как исходная 15N ДНК была бы равномерно разделена между всеми цепями ДНК. Результат соответствовал гипотезе полуконсервативной репликации.[6]
Рекомендации
- ^ От Джона Кэрнса к Горацию Ф. Джадсону в книге «Восьмой день сотворения мира: создатели революции в биологии» (1979). Книги Touchstone, ISBN 0-671-22540-5. 2-е издание: Cold Spring Harbor Laboratory Press, 1996 г., мягкая обложка: ISBN 0-87969-478-5.
- ^ Уотсон Дж. Д., Крик Ф. Х. (1953). «Строение ДНК». Холодная весна Харб. Symp. Quant. Биол. 18: 123–31. Дои:10.1101 / SQB.1953.018.01.020. PMID 13168976.
- ^ Блох Д.П. (декабрь 1955 г.). «Возможный механизм воспроизведения спиральной структуры дезоксирибонуклеиновой кислоты». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 41 (12): 1058–64. Дои:10.1073 / pnas.41.12.1058. ЧВК 528197. PMID 16589796.
- ^ Дельбрюк М. (сентябрь 1954 г.). «О репликации дезоксирибонуклеиновой кислоты (ДНК)» (PDF). Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 40 (9): 783–8. Дои:10.1073 / пнас.40.9.783. ЧВК 534166. PMID 16589559.
- ^ Дельбрюк, Макс; Стент, Гюнтер С. (1957). «О механизме репликации ДНК». В McElroy, William D .; Гласс, Бентли (ред.). Симпозиум по химическим основам наследственности. Johns Hopkins Pr. стр.699–736.
- ^ Мезельсон, М. и Шталь, Ф.В. (1958). «Репликация ДНК в Escherichia coli». PNAS. 44: 671–82. Дои:10.1073 / pnas.44.7.671. ЧВК 528642. PMID 16590258.
внешняя ссылка
- Краткое выступление Мэтью Мезельсона: «Полуконсервативная репликация ДНК»
- ДНК с самого начала Анимация, объясняющая эксперимент.
- Эксперимент Мезельсона – Шталя Еще одна полезная анимация.
- Мезельсон и Шталь Экспериментальная английская анимация
- Описание эксперимента Мезельсона-Шталя. написано Натан Х. Ленц, включая исходные данные Visionlearning
| Поля, дисциплины |  | ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Учреждения | |||||||||||||||||||||||||||
| Теории, концепции | |||||||||||||||||||||||||||
| История |
| ||||||||||||||||||||||||||
| Связанный | |||||||||||||||||||||||||||
| |||||||||||||||||||||||||||