Тоготовирус - Thogotovirus - Wikipedia
| Тоготовирус | |
|---|---|
 | |
| Электронная микрофотография из Вирус бурбона (шкала: 100 нм) | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область: | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Негарнавирикота |
| Учебный класс: | Insthoviricetes |
| Заказ: | Articulavirales |
| Семья: | Ортомиксовирусы |
| Род: | Тоготовирус |
| Типовой вид | |
| Тогото тоготовирус | |
| Разновидность | |
| Синонимы[1] | |
Тогото-подобные вирусы | |
Тоготовирус это род из окутанный РНК-вирусы, один из семи родов в вирус семья Ортомиксовирусы. Их одноцепочечный, отрицательный смысл РНК геном имеет шесть или семь сегментов. Тоготовирусы отличаются от большинства других ортомиксовирусов.[3] будучи арбовирусы - вирусы, передающиеся через членистоногие, в этом случае обычно клещи. Тоготовирусы могут реплицироваться как в клетках клещей, так и в позвоночное животное клетки; один подтип также был выделен из комары. Следствие передачи кровососом векторов заключается в том, что вирус должен распространяться системно в позвоночном хозяине - в отличие от вирусы гриппа, которые передаются воздушно-капельным путем и обычно ограничиваются дыхательная система.[4]
Тогото тоготовирус (THOV) - это типовой вид; род также содержит виды Вирус Дори (DHOV) и его подтип Баткенского вируса, а также виды или штаммы Вирус Арагуари, Вирус Аранзасского залива (ABV), Вирус бурбона, Jos вирус (JOSV) и Вирус Уполу (УПОВ), которые еще не подтверждены Международный комитет по таксономии вирусов (ICTV). Широкий спектр млекопитающие заражены представителями рода; некоторые типы также заражают птицы. THOV вызывает заболевание домашнего скота. THOV, DHOV и вирус Бурбона могут инфицировать людей и иногда были связаны с человеческими заболеваниями.
История
THOV и DHOV были выявлены в начале 1960-х гг. в Кения и Индия, соответственно.[5][6] Два случая заболевания человека, связанного с THOV, произошли в 1966 году, а авария в российской лаборатории в 1980-х годах показала, что DHOV также может вызывать заболевание у людей.[7][8] Эти два вируса изначально считались буньявирусы, но характеристика в 1980-х и начале 1990-х годов выявила сходство с вирусы гриппа.[9][10] Род «Thogoto-подобных вирусов» внутри Ортомиксовирусы был предложен в 1995 г. и признан ICTV под именем Тоготовирус в следующем году.[11][12] Название происходит от Лес Тогото в Кении, где впервые был открыт THOV.[13] С тех пор анализ последовательности пяти вирусов, обнаруженных в 1960–70-х годах, но неклассифицированных или предположительно отнесенных к Bunyaviridae привело к тому, что они были предложены в качестве дополнительных членов рода.[2][9][14][15] Другой предложенный член рода характеризовался секвенирование следующего поколения в 2014.[7]
Вирусология
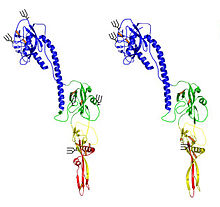
Вирусная частица окутанный. Обычно он сферический или яйцевидный, диаметром 80–120 нм.[16] Некоторые нитчатые формы наблюдаются в ТОВ, Баткен и Вирусы бурбона.[7][9][17] В одноцепочечный, –РНК геном является линейным и сегментированным, с шестью или семью сегментами размером 0,9–2,3 т.п.н. и общим размером около 10 т.п.н.[16][18] Перегруппировка сегментов между штаммами наблюдали как у клещей, так и у млекопитающих, экспериментально инфицированных более чем одним тоготовирусом, но его значение в естественных инфекциях неизвестно.[19]
Вирусные белки
Геном кодирует 7–9 белков, в том числе тримерный РНК-полимераза фермент (PA, PB1, PB2) и структурные белки нуклеопротеин (NP), который связывает вирусный геном; матричный белок (M1), который выравнивает конверт; и конверт гликопротеин (GP), который действует как рецептор вируса.[15][16]
Гликопротеин тоготовируса не похож на гликопротеины вируса гриппа (гемагглютинин и нейраминидаза ), а вместо этого показывает некоторое сходство с гликопротеином gp64 бакуловирусы, поражающие насекомых.[4][15][20] Он также имеет некоторое сходство с гемагглютинином Каранфил вирус, типовой вид новый род ортомиксовирусов, передающихся клещами.[21] Механизм, с помощью которого тоготовирусы приобрели бакуловирусоподобный гликопротеин, неизвестен. Пэт Наттолл и коллеги предположили, что приобретение позволило этим вирусам заразить клещей.[22] Эта очевидная рецепторная специфичность для клеток членистоногих не предотвращает заражение большинства тоготовирусов позвоночными.[15] Гликопротеин тоготовируса классифицируется как пенетрен класса III или γ, в котором отсутствует слитый пептид, присутствующий в гемагглютинине гриппа (пенетрен класса I или α).[23]
THOV и JOSV[2] также кодируют белок M-long (ML), который противодействует врожденный иммунитет, в частности, подавляя производство интерферон. Это уклонение от иммунитета важно для системного заражения вирусом позвоночные, но не требуется в членистоногие, у которых отсутствует интерфероновый ответ. Механизм действия ML полностью отличается от аналогичного белка в вирусы гриппа (NS1 ).[15][16][24][25]
Как и у всех ортомиксовирусов, три самых больших сегмента (1–3) кодируют три субъединицы РНК-полимеразы. В тоготовирусах сегмент 4 кодирует гликопротеин, а сегмент 5 - нуклеопротеин. В информационная РНК (мРНК) из сегмента 6 может быть сращенный для кодирования матричного белка или неслицированного для кодирования ML, который имеет 38 дополнительных аминокислоты на своем C-конец.[4][15][16][18] Для седьмого сегмента, наблюдаемого в DHOV, еще не обнаружено ни одного продукта.[18]
Жизненный цикл

Рецептор на клетке-хозяине позвоночного сиаловая кислота, который связывается вирусным гликопротеином. Вход осуществляется эндоцитоз, причем слияние вирусной и клеточной мембран происходит после подкисления везикулы. Как и другие ортомиксовирусы, вирусные транскрипция и репликация происходит в ядро клетки.[15][16] Было показано, что у некоторых представителей этого рода репликация чувствительна к Mx1 / MxA белок, который индуцируется у мышей и людей в ответ на интерферон.[9][26] В одном исследовании было показано, что этот ингибирующий эффект вызван MxA, препятствующим транспорту генома THOV в ядро.[27]
Поскольку ортомиксовирусы не кодируют кэпирующий фермент, инициация транскрипции включает разрезание вируса колпачок с 5'-конца мРНК хозяина, так что мРНК распознается машиной трансляции хозяина. Похожий "кепка «процесс используется другими ортомиксовирусами, но гораздо более длинная последовательность РНК хозяина расщепляется вместе с кэпом и включается в вирусную мРНК.[28][29]
Вирус собирается клеточная мембрана и покидает клетку почкованием.[16] Для THOV, выращенных в клетки почек детеныша хомяка вирусные частицы начинают высвобождаться через 6–8 часов после заражения, при этом значительные количества вируса все еще образуются через 24 часа после заражения. Эта скорость роста ниже, чем у вирусов гриппа, и больше похожа на вирус Quaranfil.[17][21]
Эпидемиология
Было показано, что большинство тоготовирусов заражают членистоногих, как правило, жесткий или же мягкий клещи, которые паукообразные,[7] но в одном случае комары, которые насекомые.[9] Члены также заражают птицы[15][30] и широкий спектр диких и домашних млекопитающие, включая сумчатые,[14] грызуны,[4] зайцы,[31] мангуста,[32] лошади,[6] верблюды, козы, овца и крупный рогатый скот.[2][32] Три типа - THOV, DHOV и Вирус бурбона - было показано, что они заражают людей.[7] У них широкий географический диапазон.[7]
Передача позвоночным обычно происходит через клеща. вектор. THOV сохраняется у клеща, оставаясь в организме по мере прохождения стадий своего развития; это называется трансстадиальная передача.[17] Вирус может быть передан другому хосту в течение дня после прикрепления к хосту.[33] THOV может передаваться между клещами, когда они одновременно питаются явно неинфицированными морские свинки при отсутствии определяемого уровня вируса в крови.[34][35] Такая невиремическая передача также наблюдалась с другими РНК-вирусами, преимущественно передаваемыми клещами, включая блютанга, Крымско-Конго геморрагическая лихорадка, больной, клещевой энцефалит, вирус везикулярного стоматита и вирус Западного Нила вирусы.[26] Также наблюдалась передача DHOV респираторным аэрозолем.[6]
Взаимодействие с хозяином и болезнь
Клещи
Никаких серьезных патологических изменений не наблюдается. Rhipicephalus appendiculatus клещи, инфицированные THOV.[17] Вирус сконцентрирован в синганглион (галочка мозг ) в начале кормление кровью процесс, при этом доля вируса, находящегося в слюнные железы возрастает на поздней стадии кормления кровью.[17][33] Более низкие уровни вируса обнаруживаются в трахея, пищеварительный тракт и женские половые органы, но не мужские половые органы или выделительная система. Было высказано предположение, что высокий уровень вируса, присутствующего в синганглии, помогает вирусу сохраняться через метаморфоза клеща, поскольку нервная система подвергается меньшему ремоделированию, чем другие системы.[17]
Позвоночные
В лабораторных условиях несколько представителей этого рода вызывают тяжелую болезнь у мышей и хомяки.[5][6] Происходит системное распространение вируса с патологическими эффектами во многих органах и системах, включая мозг, печень, лимфатическая система, а иногда легкие и тонкий кишечник.[5][6] Лимфоциты являются основной клеткой-мишенью для DHOV.[6] Инфекция DHOV у мышей напоминает экспериментальную инфекцию гриппа у мышей и хорьки а также фатальный H5N1 грипп у людей, и был предложен в качестве модели этого заболевания.[6]
Естественные инфекции тоготовирусов у млекопитающих обычно не вызывают симптомов. THOV - серьезный ветеринарный патоген, например, вызывающий лихорадочное заболевание и аборт у овец.[5][36][37][38][39] По состоянию на февраль 2015 г. было зарегистрировано только восемь случаев заболевания человека, связанного с тоготовирусами: два - с THOV, пять - с DHOV и один - с вирусом Бурбона; два человека погибли.[7] В период инкубации для THOV - 4–5 дней.[39] Все три вируса были связаны с высокая температура. THOV и DHOV также вызвали неврологические симптомы: менингит и оптический нейромиелит в случае THOV; энцефалит в случае DHOV.[7][8][13][39] Гепатит наблюдалось с THOV.[39] Единственный случай заболевания у человека, инфицированного вирусом Бурбона, был связан со снижением уровня крови. тромбоциты и белые клетки; неврологических симптомов не наблюдалось.[7] Гриппоподобный респираторные симптомы не поступало.[7][8]
Лечение и профилактика
Нет специального лечения или вакцина доступен для тоготовирусов с февраля 2015 года. противовирусный препарат рибавирин, который имеет широкий спектр активности, включая некоторые другие ортомиксовирусы,[40] было показано, что ингибирует репликацию DHOV in vitro в одном исследовании.[41] Поддерживающая терапия используется при болезни THOV,[39] и был рекомендован США Центры по контролю и профилактике заболеваний на заражение вирусом Бурбона. Как и в случае с другими арбовирусы, предотвращение контакта с переносчиком имеет ключевое значение для профилактики.[42]
Виды и сорта
Два вида были подтверждены ICTV, THOV (типовой вид) и DHOV.[43] Два вируса имеют низкую степень идентичности последовательностей (37% для нуклеопротеина; 31% для гликопротеина оболочки), и их антитела не вступают в перекрестную реакцию.[13] Баткенский вирус - это подтип DHOV.[13] По состоянию на февраль 2015 г. к этому роду были предложены еще пять видов или штаммов.[7]
| Виды / штамм | Сегменты РНК | Диаметр (нм) | Векторы | Хозяева позвоночных | Распределение |
|---|---|---|---|---|---|
| Арагуари | 6 | 105 | Неизвестный | Серый четырехглазый опоссум, мышь | Южная Америка |
| Аранзас Бэй | 6 | 75–140 | Орнитодорос клещи | Мышь | Северная Америка |
| Баткен | 50–100 | Гиаломма клещи Aedes и Culex комары | Курица, хомяк, мышь | Азия | |
| Бурбон | ≥6 | ~100–130 | Неизвестный | Человек | Северная Америка |
| Дори | 7 | Гиаломма клещи | Птицы, заяц, лошадь, человек, мышь, жвачные животные | Африка, Азия, Европа | |
| Jos | ≥6 | 85–120 | Амблиомма и Рипицефал клещи | Мышь, зебу | Африка |
| Тогото | 6 | 100 | Амблиомма, Гиаломма и Рипицефал клещи | Полосатый мангуст, осел, человек, грызуны, жвачные животные | Африка, Азия, Европа |
| Уполу | 6 | 75–120 | Орнитодорос клещи | Мышь | Австралия |
Типовой вид выделен жирным шрифтом. Обратитесь к тексту для ссылок.
THOV-подобные вирусы

Вирус Тогото (THOV)
THOV типовой вид рода. Впервые выделен от клещей, собранных от крупного рогатого скота в Лес Тогото регион Кения, возле Найроби, в 1960 г.[5] теперь известно, что он распространен на африканском континенте, а также был обнаружен в Италия и Португалия в Европе и Иран на Ближнем Востоке.[19][31][44] Несмотря на такой широкий географический диапазон, вирус демонстрирует лишь ограниченное разнообразие.[44] Его векторы включают различные твердотельные клещи, в том числе Амблиомма, Гиаломма и Рипицефал разновидность.[5][13][19][31][45]
Антитела к THOV были обнаружены у крыс и многих домашних животных, включая коз, овец, ослов, верблюдов, крупный рогатый скот и буйволов, а вирус был выделен из дикой природы. полосатый мангуст (Монго мунго).[13][32] Это вызывает серьезные заболевания домашнего скота, включая лихорадку и аборт у овец.[5][36][37][38] При искусственных лабораторных инфекциях он очень патогенен в хомяки а также заражает мышей.[5] Известно, что вирус заражает людей в естественных условиях.[7][31]
Вирусная частица обычно имеет сферическую форму с некоторыми нитевидными формами; диаметр составляет около 100 нм.[17] В геноме шесть сегментов РНК.[4]
Вирус Арагуари
Вирус Арагуари был впервые выделен из Серый четырехглазый опоссум (Филандер опоссум) в Серра-ду-Навиу, Амапа, Бразилия в 1969 г.[14] Его способ передачи неизвестен.[6] При лабораторных инфекциях он патогенен для мышей. Вирион имеет диаметр около 105 нм. В геноме шесть сегментов РНК. На основании данных о частичной последовательности было установлено, что вирус наиболее близок к THOV.[14]
Вирус Аранзас-Бей (ABV)
ABV был обнаружен в роду мягкотелых клещей. Орнитодорос в гнездах морских птиц на юге Техас, США, 1975 г .; это был первый представитель рода, обнаруженный в Северной Америке.[7][15] Естественных позвоночных-хозяев выявлено не было, но вирус является высокопатогенным для мышей при лабораторных инфекциях. Частица вируса имеет сферическую или яйцевидную форму с диапазоном размеров от 75 нм × 85 нм до 120 нм × 140 нм. В геноме шесть сегментов РНК. Он больше всего похож на UPOV, с некоторым сходством с THOV и JOSV.[15]
Вирус Джос (JOSV)

JOSV был впервые изолирован от зебу (Bos indicus) в Jos, Нигерия в 1967 году. С тех пор было обнаружено, что он заражает Амблиомма и Рипицефал твердотельные клещи в нескольких странах Африки. В лаборатории вызывает у мышей тяжелую патологию. Вирусная частица имеет изменчивую морфологию, обычно яйцевидную, диаметром 85–120 нм. Геном содержит не менее шести сегментов РНК.[2] Он имеет некоторое сходство в последовательности с UPOV и ABV.[15]
Вирус Уполу (УПОВ)
УПОВ был впервые выделен на Upolu Cay в Большой Барьерный риф, Австралия в 1966 г. от мягкотелых клещей вида Ornithodoros capensis связанный с сажистая крачка (Onychoprion fuscatus). Естественных позвоночных-хозяев не выявлено, но вирус является высокопатогенным для мышей при лабораторных инфекциях. Вирион может быть сферическим с диаметром в диапазоне 75–95 нм или слегка яйцевидным с диапазоном размеров от 75 нм × 85 нм до 105 нм × 120 нм. В геноме шесть сегментов РНК. Он больше всего похож на ABV, в некоторой степени похож на THOV и JOSV.[15]
DHOV-подобные вирусы
Вирус Дори (DHOV)
DHOV был впервые изолирован от Гиаломма дромедарий заражение твердотельными клещами верблюды в Гуджарат, Индия, в 1961 г.[6] С тех пор это наблюдается в восточной Россия, Пакистан, Египет, Саудовская Аравия, Кения и южный Португалия. Переносчик обычно представляет собой вид Гиаломма, Такие как H. marginatum.[6][13][31][46][47]
Там, где преобладает DHOV, антитела к вирусу были зарегистрированы у верблюдов, коз, лошадей, крупного рогатого скота и людей.[6][13] Вирус был выделен из диких заяц, Lepus europaeus.[31][48] DHOV может заразить людей аэрозольным путем после случайного лабораторного воздействия, вызывая фебрильный болезнь и энцефалит.[6][8][31] В лабораторных условиях он очень патоген для мышей и был предложен в качестве модельной системы для высокопатогенного гриппа.[6] Также было показано, что он заражает птиц, причем вирус изолирован от баклан,[49] и антитела, наблюдаемые в водоплавающая птица.[13][15]

Баткенский вирус
Баткенский вирус был впервые выделен от твердотельных клещей вида Hyalomma plumbeum plumbeum заражение овец близ города Баткен, Киргизия, Сейчас в Кыргызстан, в 1970 году.[9][30] Также было обнаружено, что он может инфицировать комары вида Аедес каспиус Паллас и Culex hortensis Ficalbi, также в Кыргызстане.[50] Его географический диапазон ограничен Центральной Азией, Закавказье и район к северу от Каспийское море.[50] В лабораторных условиях он очень патогенен для мышей, хомяков и кур.[30] Форма вириона изменчива, наблюдается сферическая и нитчатая формы; он имеет диаметр 50–100 нм.[9] Баткен считается подтипом DHOV; вирусы обладают высокой степенью идентичности последовательностей (90% в гликопротеине оболочки; 96–98% в других белках), а их антитела перекрестно реагируют.[13][50]
Вирус бурбона
Вирус бурбона был идентифицирован в 2014 году секвенирование следующего поколения пробы крови мужчины из Округ Бурбон, Канзас, США, который заболел через несколько дней после укуса множественных клещей и впоследствии умер. Это единственный известный тоготовирус, который ассоциируется с болезнями человека в Западное полушарие. По состоянию на февраль 2015 года вирус Бурбона не был изолирован от клещей, насекомых или позвоночных животных, кроме человека. Вирус разнообразной формы, с нитевидными и сферическими формами; он имеет диаметр в широком диапазоне от 100 до 130 нм. Геном содержит не менее шести сегментов РНК. Больше всего похож на вирус DHOV и Batken.[7]
Примечания и ссылки
- ^ Прингл, К. Р. (1996). "Таксономия вирусов 1996 - бюллетень X Международного конгресса вирусологии в Иерусалиме" (PDF). Arch Virol. 141 (11). Получено 4 июн 2019.
- ^ а б c d е Буссетти А.В., Паласиос Дж., Травассос да Роса А. и др. (2012), «Геномная и антигенная характеристика вируса Джос», Журнал общей вирусологии, 93 (2): 293–98, Дои:10.1099 / vir.0.035121-0, ЧВК 3352346, PMID 21994326
- ^ Члены недавно ратифицированного рода Quaranjavirus также передаются клещами.[2]
- ^ а б c d е Кохс Г., Бауэр С., Фогт С. и др. (2010), «Инфекция вируса Тогото вызывает устойчивые ответы интерферона типа I, которые зависят от передачи сигналов RIG-I-подобной геликазой обычных дендритных клеток», Журнал вирусологии, 84 (23): 12344–50, Дои:10.1128 / jvi.00931-10, ЧВК 2976394, PMID 20861272
- ^ а б c d е ж грамм час Хейг Д.А., Вудалл Дж. П., Данскин Д. (1965), «Вирус Тогото: до сих пор не описанный агент, выделенный от клещей в Кении» (PDF), Журнал общей микробиологии, 38 (3): 389–94, Дои:10.1099/00221287-38-3-389, PMID 14329965
- ^ а б c d е ж грамм час я j k л м Матео Р.И., Сяо С.Ю., Лей Х., Д.А. Роса А.П., Теш Р.Б. (2007), «Вирус Дори (Orthomyxoviridae: тоготовирус) инфекция у мышей: модель патогенеза тяжелой ортомиксовирусной инфекции », Американский журнал тропической медицины и гигиены, 76 (4): 785–90, Дои:10.4269 / ajtmh.2007.76.785
- ^ а б c d е ж грамм час я j k л м п Косой О.И., Ламберт А.Дж., Хокинсон Д.Д., Пастула Д.М., Goldsmith CS, Hunt DC, Staples JE (май 2015 г.). «Новый тоготовирус, связанный с лихорадкой и смертью, США, 2014 г.». Возникновение инфекции. Дис. 21 (5): 760–4. Дои:10.3201 / eid2105.150150. ЧВК 4412252. PMID 25899080.
- ^ а б c d Бутенко А.М., Лещинская Е.В., Семашко И.В., Донец М.А., Мартьянова Л.И. (1987), «[Вирус Дори - возбудитель болезней человека. 5 случаев лабораторного заражения]», Вопросы вирусологии (на русском), 32: 724–29
- ^ а б c d е ж грамм Фрезе М., Вебер М., Вебер Ф, Спет В., Халлер О. (1997), «Чувствительность к Mx1: вирус Баткена - ортомиксовирус, тесно связанный с вирусом Дори», Журнал общей вирусологии, 78 (10): 2453–58, Дои:10.1099/0022-1317-78-10-2453, PMID 9349464
- ^ Клеркс Дж. П., Фуллер Ф., Епископ Д. Х. (май 1983 г.). «Клещевые вирусы, структурно похожие на ортомиксовирусы». Вирусология. 127 (1): 205–19. Дои:10.1016/0042-6822(83)90384-7. PMID 6858001.
- ^ 6-й отчет ICTV (PDF), Международный комитет по таксономии вирусов, получено 9 марта 2015
- ^ Прингл CR, Таксономия вирусов 1996 - Бюллетень X Международного конгресса вирусологов в Иерусалиме (PDF), Международный комитет по таксономии вирусов, получено 9 марта 2015
- ^ а б c d е ж грамм час я j Бюхен-Осмонд С., изд. (2006), «Индекс вирусов - Orthomyxoviridae», ICTVdB - Универсальная база данных вирусов, версия 4, Колумбийский университет, архив из оригинал 2 апреля 2015 г., получено 3 июля 2020
- ^ а б c d Да Силва Е.В., Да Роса А.П., Нуньес М.Р., Диниз Дж.А., Теш Р.Б., Круз А.С. и др. (Декабрь 2005 г.). «Вирус Арагуари, новый член семейства Orthomyxoviridae: серологические, ультраструктурные и молекулярные характеристики». Являюсь. J. Trop. Med. Hyg. 73 (6): 1050–58. Дои:10.4269 / ajtmh.2005.73.1050. PMID 16354811.
- ^ а б c d е ж грамм час я j k л м Бризе Т., Чоудхари Р., Травассос да Роса А. и др. (2014), "Вирус Уполу и Вирус Аранзасского залива, два предполагаемых буньявируса, являются новыми членами семейства Ортомиксовирусы", Журнал вирусологии, 88 (10): 5298–09, Дои:10.1128 / jvi.03391-13, ЧВК 4019087, PMID 24574415
- ^ а б c d е ж грамм «Тоготовирус», ViralZone, Швейцарский институт биоинформатики, получено 6 марта 2015
- ^ а б c d е ж грамм Бут Т.Ф., Дэвис С.Р., Джонс Л.Д., Стонтон Д., Наттолл, Пенсильвания (май 1989 г.). «Анатомические основы инфекции вирусом Тогото в культуре клеток BHK и в векторе иксодовых клещей, Rhipicephalus appendiculatus». J. Gen. Virol. 70 (Pt 5) (5): 1093–104. Дои:10.1099/0022-1317-70-5-1093. PMID 2543769.
- ^ а б c Кинг AM, Адамс MJ, Carstens EB, Lefkowitz EJ, ред. (2011), Таксономия вирусов: девятый доклад Международного комитета по таксономии вирусов, Академическая пресса, п. 757
- ^ а б c Дэвис К.Р., Джонс Л.Д., Грин Б.М., Наттолл, Пенсильвания (1987), "В естественных условиях реассортация вируса Thogoto (вирус клещевого гриппа) после орального заражения Rhipicephalus appendiculatus клещи " (PDF), Журнал общей вирусологии, 68 (9): 2331–38, Дои:10.1099/0022-1317-68-9-2331, PMID 3655743
- ^ Morse MA, Marriott AC, Nuttall PA (1992), «Гликопротеин вируса Тогото (клещевой ортомиксоподобный вирус) связан с гликопротеином бакуловируса GP64», Вирусология, 186 (2): 640–46, Дои:10.1016 / 0042-6822 (92) 90030-с
- ^ а б Вирусы Presti RM, Zhao G, Beatty WL, Mihindukulasuriya KA, Travassos da Rosa APA (2009), "Quaranfil, Johnston Atoll и Lake Chad являются новыми членами этого семейства. Ортомиксовирусы", Журнал вирусологии, 83 (22): 11599–606, Дои:10.1128 / jvi.00677-09, ЧВК 2772707, PMID 19726499
- ^ Эллисон А.Б., Баллард Дж. Р., Теш Р. Б., Браун Дж. Д., Рудер М. Г., Кил М. К. и др. (Январь 2015 г.). «Циклическая массовая смертность птиц на северо-востоке США связана с новым ортомиксовирусом». Дж. Вирол. 89 (2): 1389–403. Дои:10.1128 / JVI.02019-14. ЧВК 4300652. PMID 25392223.
- ^ Гарри CE, Гарри RF (2008), «Вычислительные анализы протеомики показывают, что белки суперсемейства бакуловирусов GP64 являются пенетренами класса III», Журнал вирусологии, 5: 28, Дои:10.1186 / 1743-422x-5-28, ЧВК 2288602, PMID 18282283
- ^ Haller O, Kochs G, Weber F (2006), "Схема ответа интерферона: индукция и подавление патогенными вирусами", Вирусология, 344 (1): 119–30, Дои:10.1016 / j.virol.2005.09.024, ЧВК 7125643, PMID 16364743
- ^ Вирус Тогото: Молекулярная биология ортомиксовируса, передаваемого клещами, Universitäts Klinikum Freiburg, получено 7 марта 2015
- ^ а б Куно Дж., Чанг Дж. Дж. (2005), «Биологическая передача арбовирусов: пересмотр компонентов, механизмов и уникальных свойств, а также их эволюционные тенденции», Обзоры клинической микробиологии, 18 (4): 608–37, Дои:10.1128 / cmr.18.4.608-637.2005, ЧВК 1265912, PMID 16223950
- ^ Kochs G, Haller O (1999), "Интерферон-индуцированная человеческая MxA GTPase блокирует ядерный импорт нуклеокапсидов вируса Thogoto", Труды Национальной академии наук США, 96 (5): 2082–86, Bibcode:1999PNAS ... 96.2082K, Дои:10.1073 / пнас.96.5.2082, ЧВК 26740, PMID 10051598
- ^ Leahy MB, Dessens JT, Nuttall PA (1997), «Полимеразная активность вируса Thogoto in vitro: доказательства уникального механизма захвата крышки у клещевого ортомиксовируса», Журнал вирусологии, 71 (11): 8347–51, ЧВК 192294, PMID 9343188
- ^ Guilligay D, Kadlec J, Crépin T., Lunardi T., Bouvier D, et al. (2014), «Сравнительный структурный и функциональный анализ доменов, захватывающих кэп-полимеразы ортомиксовируса», PLOS ONE, 9 (1): e84973, Bibcode:2014PLoSO ... 984973G, Дои:10.1371 / journal.pone.0084973, ЧВК 3893164, PMID 24454773
- ^ а б c Львов Д.К., Карась Ф.Р., Цыркин Ю.М., Варгина С.Г., Тимофеев Е.М. и др. (1974), «Баткенский вирус, новый арбовирус, выделенный от клещей и комаров в Киргизской ССР», Archiv für die gesamte Virusforschung, 44: 70–73, Дои:10.1007 / bf01242183
- ^ а б c d е ж грамм Грац N (2006), Болезни, передаваемые переносчиками и грызунами, в Европе и Северной Америке: распространение, бремя общественного здравоохранения и меры борьбы, Издательство Кембриджского университета, стр. 108–9
- ^ а б c Оген-Одои А., Миллер Б.Р., Хапп С.М., Мопин Г.О., Буркот Т.Р. (март 1999 г.), «Выделение вируса тогото (Ортомиксовирусы) от полосатого мангуста, Монго мунго (Herpestidae), в Уганде ", Американский журнал тропической медицины и гигиены, 60 (3): 439–40, Дои:10.4269 / ajtmh.1999.60.439, PMID 10466973
- ^ а б Кауфман WR, Nuttall PA (2003) "Rhipicephalus appendiculatus (Акари: Иксодовые): динамика Вирус Тогото заражение самок клещей при кормлении морских свинок », Экспериментальная паразитология, 104 (1–2): 20–25, Дои:10.1016 / с0014-4894 (03) 00113-9
- ^ Джонс Л.Д., Дэвис К.Р., Стил Г.М., Наттолл, Пенсильвания (август 1987 г.). «Новый способ передачи арбовируса с участием невиремического хозяина». Наука. 237 (4816): 775–7. Bibcode:1987Наука ... 237..775J. Дои:10.1126 / science.3616608. PMID 3616608.
- ^ Джонс Л.Д., Наттолл П.А. (1989). «Невиремическая передача вируса Тогото: влияние времени и расстояния». Пер. R. Soc. Троп. Med. Hyg. 83 (5): 712–24. Дои:10.1016/0035-9203(89)90405-7. PMID 2617637.
- ^ а б Дункансон Г.Р. (2013), Медицина и хирургия сельскохозяйственных животных: для ветеринаров мелких животных, CABI, п. 110
- ^ а б Мараморош К., Макинтош А.Х. (1994), Системы культивирования клеток членистоногих, CRC Press, п. 165
- ^ а б Frese M, Kochs G, Meier-Dieter U, Siebler J, Haller O (1995), «Человеческий белок MxA ингибирует клещевой вирус Тогото, но не вирус Дори», Журнал вирусологии, 69: 3904–09
- ^ а б c d е Бергер С.А., Калишер С.Х., Keystone JS (2003 г.), Экзотические вирусные заболевания: глобальное руководство, BC Decker, стр. 179–80.
- ^ Сидвелл Р.В., Хаффман Дж. Х., Харе Г. П., Аллен Л. Б., Витковски Дж. Т., Робинс Р. К. (август 1972 г.), «Широкий спектр противовирусной активности виразола: 1-f8-D-рибофуранозил-1,2,4-триазол-3-карбоксамид. ", Наука, 177 (4050): 705–6, Bibcode:1972Научный ... 177..705С, Дои:10.1126 / science.177.4050.705, JSTOR 1734763, PMID 4340949
- ^ Белов А.В., Ларичев В.Ф., Галкина И.А., Хуторецкая Н.В., Бутенко А.М. и др. (2008), «[Активность российского рибавирина in vitro на моделях вируса крымско-конголезской геморрагической лихорадки, вируса лихорадки Рифт-Валли и вирусов Тахина и Дори]», Вопросы вирусологии (на русском), 53: 34–35
- ^ «Вирус Бурбона», Сайт CDC, Отдел трансмиссивных болезней, CDC, получено 4 марта 2015
- ^ Список основных видов ICTV 2014, версия 2, Международный комитет по таксономии вирусов, 2014, получено 7 марта 2015
- ^ а б Калишер Ч., Карабацос Н., Филипе А. Р. (1987), "Антигенная однородность штаммов топотипа вируса Тогото из Африки, Европы и Азии", Американский журнал тропической медицины и гигиены, 37 (3): 670–73, Дои:10.4269 / ajtmh.1987.37.670, PMID 3688319
- ^ Санг Р., Оньянго С., Гачоя Дж., Мабинда Э., Кононгой С. и др. (2006), «Эпиднадзор за арбовирусами, передающимися клещами, у товарного домашнего скота, Найроби, Кения», Возникающие инфекционные заболевания, 12 (7): 1074–1080, Дои:10.3201 / eid1207.060253, ЧВК 3291068, PMID 16836823
- ^ Darwish MA, Hoogstraal H, Roberts TJ, Ghazi R, Amer T (1983), «Сероэпидемиологическое исследование Bunyaviridae и некоторых других арбовирусов в Пакистане», Труды Королевского общества тропической медицины и гигиены, 77 (4): 446–50, Дои:10.1016/0035-9203(83)90108-6, PMID 6415873
- ^ Аль-Халифа М.С., Диаб FM, Халил Г.М. (2007), «Вирусы, угрожающие человеку, выделенные от клещей в Саудовской Аравии», Саудовский медицинский журнал, 28: 1864–67
- ^ Львов Д.Н., Джаркенов А.Ф., Аристова В.А., Ковтунов А.И., Громашевский В.Л. (2002), «[Выделение вирусов дхори (Orthomyxoviridae, Thogotovirus) и вируса крымско-конго геморрагической лихорадки (Bunyaviridae, Nairovirus) из вируса хакерской»).Lepus europaeus) и его клещи Hyalomma marginatum в средней зоне дельты Волги, Астраханская область, 2001] », Вопросы вирусологии (на русском), 47: 32–36
- ^ Яшкулов КБ; Щелканов М.И. Львов СС; Джамбинов С.Д .; Галкина И.В. и другие. (2008), «[Выделение вируса гриппа A (Orthomyxoviridae, вирус гриппа A), вируса Dhori (Orthomyxoviridae, Thogotovirus) и вируса болезни Ньюкасла (Paromyxoviridae, Avulavirus) на острове Малый Жемчужный в северо-западной части Каспийского моря». Море]", Вопросы вирусологии (на русском), 53: 34–38
- ^ а б c Альховский С.В.; Львов ДК; Щелканов М.И. Щетинин AM; Дерябин PG; и другие. (2014), «[Генетическая характеристика вируса Баткена (BKNV) (Orthomyxoviridae, Thogotovirus), выделенного из клещей Ixodidae Hyalomma marginatum Koch, 1844 и комаров Aedes caspius Pallas, 1771, а также Culex hortensis Ficalbi, 1889 г. Центральная Азия]", Вопросы вирусологии (на русском), 59: 33–37