Евоплоцефал - Euoplocephalus
| Евоплоцефал | |
|---|---|
 | |
| Череп экз. ТМП 1991.127.1 | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Заказ: | †Орнитишия |
| Семья: | †Анкилозавриды |
| Подсемейство: | †Анкилозаврины |
| Род: | †Евоплоцефал Ламбе, 1910 |
| Типовой вид | |
| †Стереоцефальные пачки Ламбе, 1902 г. | |
| Разновидность | |
| |
Евоплоцефал (/juˌɒплoʊˈsɛжələs/ тыOP-лох-SEF-ə-ləs ) - род очень крупных, травоядных анкилозавр динозавры, живущие во время Поздний мел из Канада. У него есть только один названный вид, Euoplocephalus пачки.
Первая окаменелость Евоплоцефал был основан в 1897 году в Альберта. В 1902 году он получил название Стереоцефалия, но это имя уже был отдан насекомому, поэтому его заменили в 1910 году. Позже, многие другие анкилозаврид останки были найдены из Кампанский Северной Америки и часто делали отдельные роды. В 1971 г. Уолтер Кумбс пришел к выводу, что все они принадлежали Евоплоцефал который тогда стал бы одним из самых известных динозавров. Однако в последнее время специалисты пришли к противоположному выводу, ограничивая подлинные находки Евоплоцефал около десятка экземпляров. К ним относятся несколько почти полных скелетов, но, тем не менее, о строении животного известно очень много.
Евоплоцефал был около пяти с половиной метров в длину и весил около двух с половиной тонн. Его тело было с низкой посадкой, очень плоским и широким, стояло на четырех крепких ногах. На голове у него была короткая висящая морда с роговым клювом, чтобы кусать растения, перевариваемые в большом кишечнике. Как и другие анкилозавриды, Евоплоцефал был в основном покрыт костлявым броня пластинки, среди них ряды крупных овальных с высокими ребрами щитков. Шею защищали два костяных кольца. Он также мог активно защищаться от хищников, таких как Горгозавр используя тяжелый клуб на конце его хвоста.
Описание
Размер
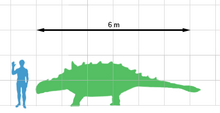
Среди анкилозавридов, Евоплоцефал был превышен по размеру только на Анкилозавр, и, возможно Тархия и Cedarpelta. Евоплоцефал был около 5,5 метров в длину и весил около 2,5 тонны (2,8 коротких тонны).[1] Он был также шириной 2,4 метра (7,9 фута). Как и другие анкилозавриды, у него было очень широкое и плоское туловище с низкой посадкой, около четырех футов высотой, расположенное на четырех коротких ногах.
Отличительные черты
Череп Евоплоцефал можно отличить от большинства других анкилозаврид по нескольким анатомическим деталям, включая: рисунок костной скульптуры в области перед глазами; форма пальпебральный кости (мелкие косточки над глазами),[2] которые могли служить костлявыми веками;[3] поверхностность преддверие носа у входа в полость носа;[2] то медиальный изгиб зубных рядов верхней челюсти; и зубы, которые относительно малы, не имеют настоящих цингула, и с переменным рифлением зубчики.[2] Однако эти черты характерны для ряда близкородственных форм, некоторые из которых в прошлом были включены в этот род.[4] Комбинируя такие формы, Уолтер Кумбс и Тереза Марьянская в 1990 году заявил, что Евоплоцефал можно выделить по четырем признакам. В предчелюстные кости, передние кости рыла не покрыты дермальными окостенениями. Наружные костные ноздри имеют щелевидную форму, обращены вперед, каждая из них разделена вертикальным костным листом или перегородкой. Клюв имеет ширину, равную или превышающую расстояние между задними частями. верхнечелюстной, верхняя щека, зубные ряды. На стопе три пальца, плюсневые кости с пальцами ног.[5]

В 2013, Виктория Арбор и Фил Карри проведена дифференциальная диагностика, установка Евоплоцефал кроме ближайших родственников. По сравнению с Анодонтозавр и Сколозавр, Евоплоцефал отсутствуют круглые остеодермы у основания чешуйчатых и квадратично-скуловых рогов. По сравнению с Анодонтозавр у него отсутствуют маленькие остеодермы в основании больших остеодерм на первом шейном полукольце, но, в отличие от этого рода, на виде сверху действительно есть большие округлые остеодермы по бокам хвостовой булавы. Он отличается от Диоплозавр крестцовые ребра направлены перпендикулярно наружу. Он отличается от Сколозавр в обладании килеватыми остеодермами с круглым или овальным основанием на вершине и по бокам первого шейного полукольца и с более короткой задней лопастью подвздошной кости. Евоплоцефал отличается от Анкилозавр в наличии направленных вперед наружных ноздрей и отсутствии непрерывного киля между чешуйчатым рогом и надглазничными костью.[4]
Скелет

Череп Евоплоцефал напоминает усеченный равносторонний треугольник если смотреть сверху, он немного шире, чем длина. Самый крупный из известных черепов, экземпляр AMNH 5403, имеет длину 411 миллиметров и ширину 478 миллиметров. Большинство элементов черепа полностью слиты, и у динозавров обычно есть два отверстия в черепе. анторбитальное окно и верхнее височное отверстие закрылись. Череп имеет от девятнадцати до двадцати четырех зубов на каждой верхней челюсти. Передняя кость носа, предчелюстная кость, беззубый. Зубы очень маленькие, максимальная высота и ширина всего 7,5 миллиметра. Сильно опущенная морда тупая, широкая и высокая, заполнена очень сложными дыхательными путями и пазухами, форма и функция которых еще полностью не изучены. С каждой стороны по две внешние ноздри. Нижняя челюсть имеет очень вогнутый верхний обод с двадцатью одним зубом. На его передней части имеется короткое низкое удлинение, обеспечивающее контакт с широкой костной частью нижнего рогового клюва, которая входит в верхний клюв морды.[2]
Как и у большинства четвероногих орнитизхов, шея умеренно короткая. Лопатка массивная и крепкая, а очень крепкие передние конечности короче задних.[6] Хвост длинный и оканчивается костлявой булавой. Старые реставрации Евоплоцефал и отклоненные синонимы (Диоплозавр, Сколозавр ) часто показывают дубинку с двумя большими вертикальными шипами. Это ошибка, вызванная восстановлением Сколозавр к Франц Нопца; у экземпляра, который он использовал, был неполный хвост, который останавливался сразу за парой конических шипов, которые, как теперь известно, были расположены на полпути вдоль его длины. Он восстановил конец хвоста сразу после шипов. Другие художники сочетали шипы с хвостовой булавой, усугубляя неточность.[6] Узкая дистальная половина хвоста укреплена пучками окостеневшие сухожилия.
В позвоночник из Евоплоцефал состоит как минимум из семи шейные позвонки, по крайней мере, одиннадцать «свободных» спинных позвонков, обычно четыре крестцовых позвонка, образующие слитый «крестцовый стержень» перед крестец собственно, три "истинных" крестцовый позвонки и от одного до четырех хвостово-крестцовых.[4] Как и другие анкилозавры,[7] последние четыре спинных позвонка и первый хвостовой позвонок, таким образом, сливаются с крестцом, образуя усиленный синсакрум не менее восьми позвонков,[6] шипы которых образуют сросшуюся надбранную пластинку, также включающую зигапофизы. Есть по крайней мере двадцать один хвостовой позвонок; общее количество хвостовых позвонков неизвестно, потому что примерно десять слились, образуя часть хвостовой булавы,[6] доведя общее количество до тридцати. Это слияние также наблюдается у других анкилозаврид; возможно, что степень слияния является возрастной особенностью.[8][9]
В плечевая кость очень прочный, с сильно расширенными верхними и нижними шарнирами в сочетании с узким валом. На верхнем валу огромная дельто-грудной гребень нижняя часть которого не сливается постепенно с валом, а изогнута вперед, образуя толстый выступ или выступ. Все это говорит об очень тяжелой мускулатуре. В нижнем рычаге прочная локтевая кость имеет хорошо развитую олекранонный отросток. Кости запястья и кисти малоизвестны. в таз, переднее лезвие подвздошная кость растопырен вперед, достигая самого широкого места живота, чтобы поддерживать кишечник. Это лезвие также образует костяную полку на задней стороне тела. Задняя лопасть подвздошной кости короче диаметра тазобедренной впадины, за которой она находилась, что означает, что нога расположена на заднем конце таза, у основания хвоста и намного ближе к средней линии, чем по бокам живота. В лобковая кость неизвестно. В седалищная кость представляет собой короткую изогнутую вертикально расположенную костную ленту. Бедренная кость короткая, крепкая, прямая, с низким четвертый вертел расположен ниже средней точки вала. Крепкая большеберцовая кость короче бедра. Стопа малоизвестна, но функционально трехактильная с копытообразной формой вместо острых когтей.[4]
Броня

Голова и тело Евоплоцефал были покрыты костным панцирем, за исключением частей конечностей и, возможно, дистального отдела хвоста. Доспех состоял из остеодермы, окостенения кожи, которые не являются частью собственно скелета. Этот доспех был подробно описан в 1982 г. Кеннет Карпентер, который, однако, в значительной степени основывался на очень полном экземпляре NHMUK R5161, голотипе Сколозавр,[6] какой род больше не рассматривается как синоним Евоплоцефал. Если ограничиться определенным материалом последнего рода, мало что известно о точной конфигурации доспехов, за исключением головы и шеи. Тогда наиболее информативным образцом в этом отношении был бы ROM 1930, сохранивший некоторые остеодермы туловища в их исходном положении.[4]
В любом случае, большая часть доспехов состояла из маленьких косточек, костлявых круглых щитки диаметром менее пяти миллиметров, из которых сотни часто встречаются на одном экземпляре. Если броня была настроена так же, как у Сколозаврмногие из этих косточек срослись в своего рода мостовую, образуя поперечные полосы на теле. Полосчатая конструкция, как полагают, давала некоторую свободу передвижения.[6] Четыре из этих полос могли присутствовать на передней половине хвоста, три - на тазу, возможно, слитые в единый «крестцовый щит», и четыре на передней части туловища. Вставкой в эти полосы были горизонтальные ряды более крупных овальных, плоских или килевидных щитков. Типы крупных щитков различаются по регионам тела. Возможно, щитки на плече, около средней линии тела, были самыми большими и высокими; ROM 1930 включает несколько остеодерм с длиной основания пятнадцать сантиметров. О броне конечностей известно немного. Большие килевидные пластины присутствовали на плечах, как показано на образце TMP 1997.132.01, сохраняющем круглую остеодерму около плечевой кости диаметром двадцать сантиметров и более узкие шипы, связанные с нижней частью руки. Шею защищали два костяных кольца, открытые с нижней стороны, которые называются шейными полукольцами. Ранее рассматривался как слияние остеодерм,[6] в этом сомневались Arbor et al. в 2013 году, который указал, что они образуют нижний слой, возможно, состоящий из окостеневшего хряща, на что указывает гладкая поверхность и тканая текстура кости. Каждое полукольцо состоит из шести прямоугольных вогнутых пластин, по три с каждой стороны. Каждая пластина имеет сверху большую килевидную остеодерму, часто не сросшуюся с ней. С Евоплоцефалэти шейные остеодермы не имеют более мелких остеодерм в основании, а их кили не выступают за задние края.[4]
Панцирь черепа состоит из большого количества косточек, называемых caputegulae ("плитки головы"), которые слились с обычными элементами черепа, в значительной степени потускневшие швы. На морде они образуют хаотичную и асимметричную мозаику. В задней части носа по средней линии присутствует одна шестиугольная пластинка большего размера. Кильчатая пластинка с каждой стороны, немного дальше назад, образует ободок морды. За уровнем глазниц caputegulae сливаются в единую поверхность кости. Верхний край глазниц образован двумя остеодермами пирамидальной формы, направленными в стороны и назад. Кроме того, Евоплоцефал было два пирамида плоские чешуйчатые «рога», растущие из задних углов головы. Между ними затылочный гребень покрыт двумя остеодермами с каждой стороны. В нижней задней части черепа имеется четырехъядерный рог в виде огромной остеодермы в форме языка, выступающей снизу.[4]
Открытие и виды

Канадский палеонтолог Лоуренс Моррис Ламбе обнаружил первый экземпляр 18 августа 1897 г. в районе нынешнего Провинциальный парк динозавров, в долине Ред Дир Ривер, Альберта, Канада. В 1902 году это ископаемое, CMN 210 (также NMC 210) был обозначен как голотип образец типовой вид Стереоцефальные пачки. Этот образец состоит из верхней части черепа и поперечной серии из пяти щитков, которые были частью шейного полукольца. Родовое название произошло от греческого στερεός, стереосистемы, «твердое тело» и κεφαλή, кефале, «голова», что означает грозную броню.[10] Однако название рода уже было озабоченный - имя уже было дано насекомому, жук Стереоцефалия Линч 1884 - поэтому Ламбе изменил его на Евоплоцефал в 1910 г. Combinatio nova (название новой комбинации) Euoplocephalus пачки.[11] В типовой вид останки Стереоцефальные пачки. В 1915 г. Эдвин Хенниг классифицированный E. пачки под родом Палеосцинк Leidy 1856, чеканка Пачки Palaeoscincus.[12] Однако сегодня Палеосцинк считается номен дубиум на основе неопределенных анкилозавров. В 1964 г. Евоплоцефал был от Оскар Кун упомянутый Анкилозавр, как Пачки анкилозавра.[13]
Название рода Евоплоцефал, что означает «хорошо вооруженная голова», происходит от Греческий слова Европа (εὖ) означает "хорошо", хопло~ (ὁπλο ~) означает «вооруженный», и кефал (κεφαλή), что означает «голова».[14] В официальной научной литературе это имя было написано с более чем дюжиной различных ошибок. В конкретное имя пачки означает "надежно защищенный" в латинский. Единственный действительный разновидность известный сегодня Euoplocephalus пачки.
Упомянутый материал

В начале двадцатого века в Северной Америке было обнаружено гораздо больше окаменелостей анкилозаврид. Некоторые были упомянуты Евоплоцефал, другие названы как отдельные роды. Однако в 1971 г. Уолтер Кумбс представил диссертация содержащий важную переоценку североамериканских анкилозавров. Он отметил, что среди множества экземпляров, похожих на Евоплоцефал, их черепа так сильно различались, что либо каждый известный экземпляр должен быть новым видом, либо все они представляли индивидуальные вариации в пределах одного вида: Euoplocephalus пачки.[8] Исходя из этого предположения, что на кампанском этапе верхнего мела существовал только один вид анкилозавров, Кумбс назвал эти роды синонимами. Анодонтозавр, Диоплозавр, и Сколозавр с Евоплоцефал и виды А. ламбей, D. acutosquameus, и С. катлери с E. пачки, создавая вид, который существовал почти десять миллионов лет, или весь Кампанский.[15] Окаменелости, которые теперь относятся к этому виду, содержат более сорока особей, обнаруженных в Альберта, Канада и Монтана в Соединенных Штатах, что сделало бы Евоплоцефал самый известный анкилозаврид. Это включало пятнадцать черепов, зубы и несколько почти целых скелетов, найденных с еще прикрепленными доспехами.[7] Чаще всего встречаются отдельные бронеплиты. В 1978 году Кумбс даже включил азиатских анкилозавридов. Тархия в роде, переименовав его в Euoplocephalus giganteus.[16]
Синонимия всех кампанских североамериканских анкилозаврид прослеживалась несколько десятилетий.[7] пока ученые из Университет Альберты начал заново исследовать окаменелости. Исследование 2009 года показало, что Диоплозавр на самом деле действительный таксон и идентифицировал уникальные характеристики, которые отличают его от Евоплоцефал, включая его треугольные когти.[15] Виктория Арбор (2010) утверждали, что Анодонтозавр (известный из формации каньона Подкова) отличается от Евоплоцефал и также является допустимым таксоном; по словам Арбор, Анодонтозавр отличается от Евоплоцефал характерным орнаментом черепа и шейного полукольца, а также морфологией хвостовой булавы, в том числе наличием остеодермы остеодермы треугольного выступа в Анодонтозавр. Кроме того, Арбор (2010) предложил переназначить все образцы анкилозаврина из формации Хорсшу-Каньон из Евоплоцефал к Анодонтозавр.[17] Срок действия Анодонтозавр был принят в двух последующих исследованиях. Первый, опубликованный Пол Пенкальски и Уильям Т. Блоуз в 2013 году повторно валидировано Сколозавр также.[18] Второе исследование, Пенкальски (2013), назвало и описало Охкотокия из Монтаны на основании останков, которые первоначально считались относящимися к Евоплоцефал.[19]
Palaeoscincus asper, "грубый", теперь считается Евоплоцефал. Это сомнительный зуб таксон из позднего кампана Формирование парка динозавров Альберты, названный Ламбе в 1902 году.[20] Он состоит из одного зуба, экз. NMC 1349.[21]
В 2013 году Арбор ограничила экземпляры, которые можно достоверно отнести к Евоплоцефал до самых низких тридцати метров формации Парка динозавров. В этом случае материал, помимо голотипа, будет состоять из частичных скелетов с черепом AMNH 5337, AMNH 5403, AMNH 5404, AMNH 5405, ROM 1930 и UALVP 31; частичный скелет без черепа AMNH 5406; CMN 842, шейное полукольцо; CMN 8876, череп, ТМП 1979.14.74, фрагмент черепа; и UALVP 47977, часть крыши черепа. Поэтому руки, ноги и хвост, включая булаву, полностью не известны.[4]
Классификация

В 1910 году Ламбе назначил Евоплоцефал к Стегозаврия, группа, включающая все формы бронированных динозавров и, таким образом, имеющая гораздо более широкий диапазон, чем нынешняя концепция. В 1917 г. Чарльз Уитни Гилмор назначил его Анкилозавриды.[22] Сегодня, Евоплоцефал все еще рассматривается как анкилозаврид, но как член Анкилозаврия а не стегозаврия. Вероятно, он также является членом производной подгруппы Анкилозаврины. Недавнее расщепление анкилозавридного кампанского материала Северной Америки усложнило вопрос о прямом родстве Евоплоцефал. Пенкальски (2013) исполнил небольшой филогенетический анализ некоторых анкилозаврин образцы. Единственный Анодонтозавр экземпляр, который был включен в этот анализ, был его голотипом. Анодонтозавр был помещен в политомия с голотипом Евоплоцефал и некоторые экземпляры, которые к нему относятся, а Охкотокия был помещен в кладу с Диоплозавр, и образцы, которые, как считается, представляют Диоплозавр или же Сколозавр.[19]
Следующее кладограмма основан на 2015 г. филогенетический анализ Ankylosaurinae, проведенное Арбором и Карри:[23]
| Анкилозаврины |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Результаты более раннего анализа дерева анкилозаврид, проведенного Thompson et al. (2011), показан на этой кладограмме.[24]
| Анкилозавриды |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеоэкология
Евоплоцефалпосле синонимизации, предложенной Кумбсом (1971), считалось, что он существует гораздо дольше и был членом более различных фаун, чем любой из его современников, поскольку эти окаменелости датируются от 76,5 до 67 лет.миллион лет назад, в Кампанский -Маастрихтский возраст последнего Меловой период, и пришел из Парк динозавров и Образования Подковообразного Каньона Альберты, Формирование двух лекарств Монтаны и, возможно, из Формация Олдмана Монтаны.[15][19] Ископаемые останки, которые первоначально считались Формация реки Джудит Монтаны,[15] на самом деле из формирования парка динозавров.[25] Однако недавние исследования относили все образцы формации каньона подковы к Анодонтозавр,[17] и все образцы формации двух лекарств в Охкотокия.[19] Образец из самого нижнего Парка динозавров или, возможно, из нижележащей формации Олдмана, был снова назначен Сколозавр.[18] Хотя стратиграфический диапазон голотипа Евоплоцефал сомнительно, все образцы, которые можно надежно отнести к E. пачки пришли из нижних 40 м и верхних> 10 м формации парка динозавров. Анкилозавриды из кровли 20–25 м свиты неизвестны. Таким образом, все Евоплоцефал датируются от 76,4 (или меньше) до 75,6 миллионов лет назад, на позднем кампанском этапе.[19]
Палеобиология
По словам Кумбса, Евоплоцефал возможно, имел беглый способности наравне с современными носорог и бегемот.[26] На основании формы плечевого сочленения и расположения плечевой кости. растягивающие мышцы плеча, похоже, что плечо отклонено от тела.[27] Кумбс и Марьянска (1990) отметили, что Евоплоцефал образцы обычно обнаруживаются в виде отдельных элементов или частичных скелетов, что свидетельствует о том, что это животное имело уединенные привычки и обычно было либо уединенно, либо участвовало в небольших групповых скоплениях.[5]
Доспехи Евоплоцефал возможно, имел ороговевший покрытие, или оно могло плавать в коже, как это видно на современном крокодилы. Помимо защиты, сильно васкуляризированный броня могла сыграть роль в терморегуляция.[6] В глазные кости над глазами, возможно, обеспечивали дополнительную защиту глаз. Такие кости с Евоплоцефал были обнаружены в верхней части глазницы, а не перед верхним краем глазницы, что является более распространенным положением. Кумбс объяснил это предположением, что эти кости расположены в мускулатуре век и, вероятно, достаточно подвижны, чтобы перемещаться по глазам.[3]
Защита
Хвостовая булава анкилозаврид часто интерпретируется как защитное оружие. В Евоплоцефал, Наличие окостеневшие сухожилия только дистальная половина хвоста может поддерживать такую функцию.[28] Поскольку только дистальная половина хвоста была укреплена сухожилиями, передняя половина все еще могла свободно перемещаться из стороны в сторону. Закостеневшие сухожилия передавали бы силу удара клюшке и укрепляли опорные позвонки.[6] Скорее всего, булаву держали прямо над землей, поскольку у нее не было достаточной мускулатуры, чтобы поднять хвост очень высоко.[29] Исследование 2009 года пришло к выводу, что «большие анкилозаврические булавы могут создавать достаточную силу, чтобы сломать кость при ударах, в то время как средние и маленькие - нет».[30] Он также пришел к выводу, что «раскачивание хвоста возможно у анкилозаврид, но остается неизвестным, использовался ли хвост для межвидовой защиты, внутривидовой борьбы или того и другого».[30] Хвостовую булаву можно было повернуть низко, в сторону хрупких плюсневых костей или костей голени атакующего. тероподы.
Чувства и воздушный поток
Евоплоцефал у него были относительно маленькие глаза, но это не обязательно означает, что у него было ограниченное зрение. Сложные дыхательные пути, наблюдаемые в черепе, позволяют предположить, что Евоплоцефал обладал хорошим обонянием, хотя в 1978 г. при исследовании слепков эндокраниум не показали увеличенной обонятельной области мозга.[31] Тереза Марьянская, который много работал с монгольскими анкилозавридами, предположил, что дыхательные пути в основном использовались для обработки вдыхаемого воздуха, как у млекопитающих, на основе наличия и расположения специализированные кости,[9] которые присутствуют в Евоплоцефал.[32]
Исследование 2011 года показало, что носовые ходы Евоплоцефал были зацикленными и сложными; возможно приспособление для теплового и водного баланса и вокальный резонанс, и исследователи обнаружили увеличенную и васкуляризованную камеру в задней части носового тракта, что, по мнению авторов, было приспособлением для улучшения его обоняния. Исследователям также удалось восстановить внутреннее ухо динозавра и сделать вывод, что он способен слышать на низких частотах. Они предположили, что это могло быть адаптацией к слышанию низкочастотных резонансных звуков, издаваемых носовыми ходами.[33]
Рацион питания
Евоплоцефал, как и другие анкилозавры, считается травоядное животное.[7] У него была широкая морда, что могло указывать на то, что это была неселективная кормушка, возможно, похожая на бегемота. Это обеспечило бы разделение ниши из современников нодозавриды с более узкой мордочкой.[6] Исторически считалось, что анкилозавры питаются с помощью простых движений челюстей вверх и вниз. Георг Хаас (1969) исследовали данные о мускулах челюсти двух черепов (AMNH 5337 и 5405) и пришли к выводу, что, несмотря на большой размер черепов, связанная с ними мускулатура была относительно слабой. Он также считал, что движения челюсти в основном ортогональны, только в вертикальной плоскости. Хаас экстраполировал из этого то, что динозаврам нравится Евоплоцефал скорее всего ели относительно мягкую неабразивную растительность.[34]
Однако более поздние исследования показали, что движение челюстей вперед и в стороны возможно, а череп способен выдерживать значительные нагрузки.[7] Евоплоцефал похоже, мог делать более сложные движения. Износ зубов и суставы челюстей (в пределах нижней челюсти и на нижней челюсти-квадратный сустав) предполагают, что нижние челюсти были отведены назад во время кормления, а также немного повернуты внутрь. Это действие привело бы к резке еды.[35] Исследование, проведенное в 2014 году, показало, что анкилозавры были способны поедать волокнистый растительный материал, хотя и не в такой степени, как их родственники нодозавры или цератопсы и гадрозавры.[36]
Смотрите также
Рекомендации
- ^ Пол, Г.С., 2010, Принстонский полевой путеводитель по динозаврам, Princeton University Press, стр. 233
- ^ а б c d М. К. Викариус; А. П. Рассел (2003). "Повторное описание черепа Euoplocephalus пачки (Archosauria: Ornithischia): основа для сравнительных и систематических исследований анкилозавров динозавров ». Зоологический журнал Линнеевского общества. 137 (1): 157–86. Дои:10.1046 / j.1096-3642.2003.00045.x.
- ^ а б Кумбс В. (1972). "Костяное веко Евоплоцефал (Reptilia, Ornithischia) ». Журнал палеонтологии. 46 (5): 637–50. JSTOR 1303019..
- ^ а б c d е ж грамм час Беседка В.М .; Карри П.Дж. (2013). "Euoplocephalus пачки и разнообразие динозавров анкилозаврид в позднем меловом периоде Альберты, Канада, и Монтаны, США ». PLOS ONE. 8 (5): e62421. Bibcode:2013PLoSO ... 862421A. Дои:10.1371 / journal.pone.0062421. ЧВК 3648582. PMID 23690940.
- ^ а б Кумбс, У. П. младший; Марьянская, Т. (1990). «Анкилозаврия». В Weishampel, D. P .; Dodson, P .; Осмолка, Х. (ред.). Динозаврия. Калифорнийский университет Press. С. 456–483.
- ^ а б c d е ж грамм час я j К. Карпентер (1982). «Реконструкция скелетной и дермальной брони Euoplocephalus пачки (Ornithischia: Ankylosauridae) из позднемеловой формации Олдмана в Альберте ". Канадский журнал наук о Земле. 19 (4): 689–97. Bibcode:1982CaJES..19..689C. Дои:10.1139 / e82-058.
- ^ а б c d е Vickaryous, M.K., Maryanska, T., and Weishampel, D.B. (2004). «Анкилозаврия» У Д. Б. Вейшампеля, П. Додсона и Х. Осмольска (ред.), Динозаврия (второе издание). Калифорнийский университет Press, Беркли, 363–392.
- ^ а б Кумбс В. (1971) Анкилозавриды. Кандидат наук. диссертация, Колумбийский университет, Нью-Йорк, штат Нью-Йорк, 487 стр.
- ^ а б Марьянская Т. (1977). «Анкилозаврия (динозаврия) из Монголии». Палеонтология Полоника. 37: 85–151.
- ^ Л. М. Ламбе. (1902 г.). «Новые роды и виды из серии Belly River (средний меловой период)». Вклад геологической службы Канады в канадскую палеонтологию. 3 (2): 25–81.
- ^ Л. М. Ламбе. (1910). "Обратите внимание на теменный гребень Centrosaurus apertus и предлагаемое новое родовое название для Стереоцефальные пачки". Оттавский натуралист. 24: 149–51.
- ^ Хенниг, Э., 1915 г., Стегозаврия: Fossilium Catalogus I, Animalia pars 9, 16 п.
- ^ Кун, О., 1964, Орнитишия: Fossilium Catalogus, I: Animalia, Pars 105, 80 с.
- ^ Лидделл, Генри Джордж и Роберт Скотт (1980). Греко-английский лексикон (Сокращенное изд.). Соединенное Королевство: Издательство Оксфордского университета. ISBN 978-0-19-910207-5.
- ^ а б c d Беседка, В. М .; Burns, M.E .; Сиссонс, Р. Л. (2009). "Новое описание анкилозавридного динозавра. Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) и ревизия рода ». Журнал палеонтологии позвоночных. 29 (4): 1117–1135. Дои:10.1671/039.029.0405. S2CID 85665879.
- ^ Кумбс В.П. Младший (1978). «Семейства орнитисхиевых динозавров отряда Ankylosauria». Палеонтология. 21: 143–170.
- ^ а б Беседка, Виктория (2010). «Меловой арсенал: множество таксонов анкилозаврид в позднем меловом периоде Альберты, Канада и Монтана, США». Журнал палеонтологии позвоночных. 30 (Приложение 2): 1A – 198A. Дои:10.1080/02724634.2010.10411819. S2CID 220429286.
- ^ а б Пенкальски, П .; Удары, W. T. (2013). "Scolosaurus cutleri (Ornithischia: Ankylosauria) из формации Парка динозавров верхнего мела в Альберте, Канада ". Канадский журнал наук о Земле. 50 (2): 171–182. Bibcode:2013CaJES..50..171P. Дои:10.1139 / cjes-2012-0098.
- ^ а б c d е Пенкальский, П. (2013). «Новый анкилозаврид из двухмедицинской формации позднего мелового периода в штате Монтана, США». Acta Palaeontologica Polonica. Дои:10.4202 / app.2012.0125.
- ^ Ламбе, Л. М. (1902). «О позвоночных в среднем меловом периоде Северо-Западного края. 2. Новые роды и виды из серии Белли Ривер (средний меловой период)». Вклад в канадскую палеонтологию. 3: 25–81.
- ^ Кумбс-младший, W.P. (1990). Зубы и таксономия анкилозавров. В: Карпентер, К. и Карри, П.Дж. (ред.). Систематика динозавров: подходы и перспективы. Издательство Кембриджского университета: Кембридж, 269-279. ISBN 0-521-36672-0
- ^ Гилмор C.W. (1917). "Брахицератопс, цератопсовый динозавр из формации Две Медицины в Монтане, с примечаниями о связанных ископаемых рептилиях ". Профессиональная газета геологической службы США. 103: 1–45.
- ^ Беседка, В. М .; Карри, П. Дж. (2015). «Систематика, филогения и палеобиогеография анкилозавридных динозавров». Журнал систематической палеонтологии. 14 (5): 1–60. Дои:10.1080/14772019.2015.1059985. S2CID 214625754.
- ^ Ричард С. Томпсон; Джолион С. Пэриш; Сюзанна К. Р. Мейдмент; Пол М. Барретт (2011). «Филогения анкилозавров динозавров (Ornithischia: Thyreophora)». Журнал систематической палеонтологии. 10 (2): 301–312. Дои:10.1080/14772019.2011.569091. S2CID 86002282.
- ^ Пенкальски, П. (2001). "Вариации образцов, относящихся к Euoplocephalus пачкиВ К. Карпентер (ред.). Бронированные динозавры. Издательство Индианского университета. С. 261–298. ISBN 978-0253339645.
- ^ Кумбс В. (1978). «Теоретические аспекты беговых приспособлений у динозавров». Ежеквартальный обзор биологии. 53 (4): 393–418. Дои:10.1086/410790. JSTOR 2826581.
- ^ Кумбс В. (1978). «Мышцы передних конечностей анкилозаврии (Reptilia, Ornithischia)». Журнал палеонтологии. 52 (3): 642–57. JSTOR 1303969.
- ^ Кумбс В. (1979). «Остеология и миология задних конечностей при анкилозаврии (рептилии, орнитишии)». Журнал палеонтологии. 53 (3): 666–84. JSTOR 1304004.
- ^ Кумбс В. (1995). «Хвостовые клубы анкилозавров от среднего кампана до раннего маастрихта из западной части Северной Америки, с описанием крошечной дубинки из Альберты и обсуждением ориентации хвоста и хвостовой булавы». Канадский журнал наук о Земле. 32 (7): 902–12. Дои:10.1139 / e95-075..
- ^ а б Арбор, В. М. (2009). «Оценка силы удара от ударов хвостовой дубинки динозавров-анкилозаврид». PLOS ONE. 4 (8): e6738. Bibcode:2009PLoSO ... 4.6738A. Дои:10.1371 / journal.pone.0006738. ЧВК 2726940. PMID 19707581..
- ^ Кумбс В. (1978). "Эндокраниальный слепок Евоплоцефал (Reptilia, Ornithischia) ». Palaeontographica, Abteilung A. 161: 176–82.
- ^ Кумбс В. (1978). «Семьи орнитишского отряда анкилозавров» (PDF). Палеонтология. 21 (1): 143–70. Архивировано из оригинал (PDF) 10 марта 2014 г.
- ^ Мияшита Т., Арбор В.М., Уитмер Л.М., Карри П.Дж. (2011). "Внутренняя морфология черепа бронированного динозавра. Евоплоцефал подтверждено рентгеновской компьютерной томографической реконструкцией ". Журнал анатомии. 219 (6): 661–75. Дои:10.1111 / j.1469-7580.2011.01427.x. ЧВК 3237876. PMID 21954840.
- ^ Хаас Г. (1969). "На мускулатуре челюсти Анкилозавр". Американский музей Novitates. 2399: 1–11.
- ^ Рыбчинский, Н .; М. К. Викариус (2001). "Свидетельства сложного движения челюстей у анкилозаврид позднего мела, Euoplocephalus пачки (Dinosauria: Thyreophora) ". В К. Карпентер (ред.). Бронированные динозавры. Издательство Индианского университета. С. 299–317. ISBN 978-0253339645.
- ^ «Функциональные и палеоэкологические последствия морфологии и износа зубов для крупногабаритных травоядных динозавров из формации динозавров (верхний кампан) в Альберте, Канада». PLOS ONE. 9 (6): e98605. 11 июня 2014 г. Дои:10.1371 / journal.pone.0098605. Получено 11 октября 2020.
внешняя ссылка
 Данные, относящиеся к Евоплоцефал в Wikispecies
Данные, относящиеся к Евоплоцефал в Wikispecies СМИ, связанные с Евоплоцефал в Wikimedia Commons
СМИ, связанные с Евоплоцефал в Wikimedia Commons
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||

