Гаплогруппа D-M174 - Haplogroup D-M174
| Гаплогруппа D-M174 | |
|---|---|
| Возможное время происхождения | 50,000[1] - 60,000[2] лет BP 65 200 [95% ДИ 62 100 <-> 68 300] лн[3] |
| Возраст коалесценции | 46 300 [95% ДИ 43 500 <-> 49 100] лн.[3] |
| Возможное место происхождения | Азия[2][4][5] (возможно Центральная Азия[6] или же Юго-Восточная Азия[2]) |
| Предок | D (D-CTS3946) |
| Потомки | D-Z27276 (D1a1) D-M55 (D1a2a) D-Y34637 (D1a2b) |
| Определение мутаций | M174, IMS-JST021355, PAGES00003 |
Гаплогруппа D1 или же D-M174 является субкладом Гаплогруппа D-CTS3946. Эта гаплогруппа встречается в основном в Восточная Азия и Андаманские острова, хотя он также регулярно с низкой частотой встречается в Центральная Азия и Юго-Восточная Азия.
Происхождение
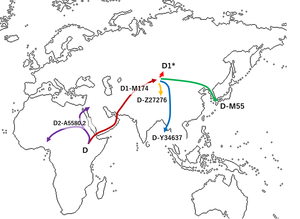
Считается, что гаплогруппа D-M174 возникла в Азия примерно за 60 000 лет до настоящего времени, это было предположено Ши и др. (2008), чтобы иметь происхождение из Юго-Восточной Азии (а не из Северной или Центральной Азии) из-за его редкости в Северной и Центральной Азии.[2][4] В то время как гаплогруппа D-M174 вместе с гаплогруппа E содержит отличительные YAP полиморфизм (который указывает на их общее происхождение), хромосомы гаплогруппы D-M174 не были обнаружены нигде за пределами Азии.[4]
Несколько исследований (Hammer et al. 2006, Shinoda 2008, Matsumoto 2009, Cabrera et al. 2018) предполагают, что отцовская гаплогруппа D-M174 возникла где-то в Центральная Азия. Согласно Хаммеру и др., Гаплогруппа D-M174 возникла между Тибетом и Горным Алтаем. Он предполагает, что в Восточную Евразию было несколько волн.[7][Почему? ]
Исследование 2017 г., проведенное Mondal et al. считает, что Люди Рианга (тибето-бирманское население) и андаманцы имеют свои ближайшие родственные линии в Восточной Азии. Джарава и Онге разделили эту линию передачи D1a2b друг с другом в течение последних ~ 7000 лет, но отклонились от японской линии передачи D1a2a ~ 53000 лет назад ". Они также предполагают, что:" Это убедительно свидетельствует о том, что гаплогруппа D не указывает на отдельное происхождение. Для андаманских популяций. Скорее, гаплогруппа D была частью постоянной вариации, принесенной расширением OOA, и позже потеряна у большинства популяций, за исключением Андаманских островов и частично в Японии и Тибете ».[8]
Исследование (Haber et al. 2019) обнаружило предложенную гаплогруппу под названием «D0» в трех нигерийских выборках. Частично из-за вероятной глубоко укоренившейся гаплогруппы «D0», а также недавно рассчитанного раннего времени расхождения для нее и ее родительской гаплогруппы, авторы предполагают африканское происхождение D0 и ее родительской гаплогруппы DE. Гаплогруппа «D0» находится за пределами M174, но разделяет с ней 7 SNP, которых нет у E.[9] «D0» также был назван «D2», а бывший D (D-M174) теперь также был назван «D1» из-за этого открытия.
Три других образца D2 также были обнаружены в Западной Азии (также в 2019 году): два в Саудовской Аравии и один в Сирии. Образец, обнаруженный в Сирии, на сегодняшний день является самым базальным образцом D2. Недавние доказательства (как также предложено Haber et al.) Предполагают, что D2 является сильно дивергентной гаплогруппой, близкой к расщеплению DE, но на ветви D и лишенной мутации M174, присущей другим линиям D.[10]
Обзор
Сегодня это встречается очень часто среди населения в Тибет, северный Мьянма, Цинхай, то Японский архипелаг, а Андаманские острова, хотя, как ни странно, не так много в остальных Индия. В Айны Японии и различных Тибето-бирманский люди (такие как Люди трипури ) примечательны тем, что обладают почти исключительно хромосомами гаплогруппы D-M174. Хромосомы гаплогруппы D-M174 также встречаются с низкой или средней частотой среди Бай, Дай, Хан, Хуэй, Маньчжурский, Мяо, Tujia, Xibe, Яо, и Чжуан из Китай и среди нескольких меньшинств Сычуань и Юньнань что говорят Тибето-бирманские языки и проживают в непосредственной близости от тибетцев, таких как Джингпо, Джино, Мосуо, Наси, Пуми, Цян, и Йи.[11]
Гаплогруппа D также встречается в популяциях собственно Китая и Кореи, но гораздо реже, чем в популяциях Тибета и Японии. Исследование, опубликованное в 2011 году, обнаружило D-M174 у 2,49% (43/1729) китайских мужчин хань, причем частота этой гаплогруппы, как правило, выше средней по направлению к северу и к западу страны (5/56 = 8,9%). D-M174 Shaanxi Han, 13/221 = 5,9% D-M174 Gansu Han, 6/136 = 4,4% D-M174 Yunnan Han, 1/27 = 3,7% D-M174 Guangxi Han, 2/61 = 3,3% D- M174 Hunan Han, 2/62 = 3,2% D-M174 Sichuan Han).[12] В другом исследовании Y-ДНК китайцев хань, опубликованном в 2011 году, гаплогруппа D-M174 наблюдалась у 1,94% (7/361) выборки неродственных китайских добровольцев-ханьцев в Университете Фудань в Шанхае, откуда происходит большинство добровольцев. прослеживается до Восточного Китая (Цзянсу, Чжэцзян, Шанхай и Аньхой).[13] В Корее гаплогруппа D-M174 наблюдалась в 3,8% (5/133) образца из Тэджона,[14] 3/85 = 3,5% выборки из Сеула,[15] 3,3% (3/90) образца из Чоллы,[16] 2,4% (2/84) образца из Кёнсана,[16] 2,3% (13/573) другого образца из Сеула,[14] 1,4% (1/72) образца из Чхунчхона,[16] 1,1% (1/87) выборки из Чеджу,[16] и 0,9% (1/110) третьей пробы из Сеула-Кёнги.[16] В других исследованиях гаплогруппа D-M174 наблюдалась у 6,7% (3/45)[17] и 4,0% (3/75)[18] образцов из Кореи без каких-либо дополнительных указаний области отбора образцов. Было опубликовано мало данных с высоким разрешением относительно филогенетического положения ханьских китайцев и корейских членов гаплогруппы D Y-ДНК, но имеющиеся данные предполагают, что большинство ханьских китайских членов гаплогруппы D должны принадлежать к кладам, часто встречающимся среди тибетцев (и особенно D-M15 clade, также встречается среди носителей некоторых языков Lolo-Burmese и Hmong-Mien), тогда как большинство корейских членов гаплогруппы D должны принадлежать к Clade D-M55, которая часто встречается среди Айны, Рюкюань, и Японский люди.[18][16][3]
Y-ДНК гаплогруппы D была обнаружена (хотя и с низкой частотой) среди современных популяций Евразийская степь, Такие как Южные Алтайцы (6/96 = 6,3% D-M174 (xM15),[19] 6/120 = 5,0% D-P47[20]), Казахи (1/54 = 1,9% D-M174,[17] 6/1294 = 0,5% D[21]), Ногайцы (4/76 = 5,3% D-M174 Кара ногайский,[22] 1/87 = 1,1% D-M174 Кубанские ногайцы[22]), Халхас (1/24 = 4,2% D-M174,[17] 3/85 = 3,5% D-M174,[15] 2/149 D-M15 + 2/149 D-P47 = 4/149 = 2,7% D-M174 всего[18]), Захчин (2/60 = 3,3% D-M174[15]), Урянхай (1/60 = 1,7% D-M174[15]), и Калмыки (5/426 = 1,2% D-M174[23]). Он также был обнаружен среди лингвистически похожих (Тюркский - или же Монгольский -говоря) современные популяции пустынь и пояса оазисов к югу от степи, такие как Югуры, Баоань, Monguors, Уйгуры, и Узбеки. При коммерческом тестировании члены были обнаружены на западе до Румыния в Европе и Ирак в Западной Азии.[24]
В отличие от гаплогруппы C-M217, гаплогруппа D-M174 не встречается в Новом Свете; он не присутствует ни в каких современных популяциях коренных американцев (северных, центральных или южных). Хотя возможно, что он попал в Новый Свет, как гаплогруппа C-M217, эти линии, по-видимому, вымерли.
Гаплогруппа D-M174 также примечательна своей довольно экстремальной географической дифференциацией, с отдельным подмножеством хромосом гаплогруппы D-M174, обнаруживаемым исключительно в каждой из популяций, которые содержат большой процент людей, чьи Y-хромосомы принадлежат к гаплогруппе D-M174: Гаплогруппа D-M15 среди Тибетцы (а также среди других Восточная Азия и Юго восточный азиат популяции, которые демонстрируют низкие частоты Y-хромосом гаплогруппы D-M174), Гаплогруппа D-M55 среди различных популяций Японского архипелага и редко среди корейцы, Гаплогруппа D-P99 у жителей Тибета и некоторых других частей центральной Евразии (например Монголия[25] и Алтай[18][19][20]). D-M174 * без положительных результатов тестирования субкладов D-M15 или D-M55 обнаруживается на высоких частотах среди Жители Андаманских островов а недавно обнаружено, что андаманский субклад D-Y34637 (D1a2b).[26] Другой тип (или типы) парагруппы D-M174 * без тестированных положительных субкладов D-M15, D-P47 или D-M55 встречается с очень низкой частотой среди Тюркский и Монгольский население Центральная Азия, в сумме не более 1%. Эта очевидно древняя диверсификация гаплогруппы D-M174 предполагает, что ее, возможно, лучше охарактеризовать как «супергаплогруппу» или «макро-гаплогруппу».
В одном исследовании частота гаплогруппы D-M174 без тестируемых положительных субкладов среди тайцев составила 10%.[2] Вс и другие. (2000) обнаружили DE-YAP / DYS287 (xM15) в 11,1% (5/45) из набора из трех образцов из Таиланда (включая 20% (4/20) северных тайцев, 20% (1/5) Так, и 0% (0/20) северо-восточного Таиланда) и в 16,7% (1/6) выборки из Гуам.[27] Между тем, авторы обнаружили D-M15 в 15% из пары образцов Яо (включая 30% (3/10) Яо Jinxiu и 0% (0/10) Яо Нандан ), 14,3% (2/14) выборки Йи, 3,8% (1/26) выборки Камбоджийцы, и 3,6% (1/28) выборки Чжуан.[27] Донг и другие. (2002) обнаружили Y-хромосомы DE-YAP в 12,5% (2/16) выборки Джингпо из Luxi City, Юньнань, 10,0% (2/20) выборки Дай из Luxi City, Юньнань, и 1,82% (1/55) выборки Nu из Гуншань и Fugong округов Юньнань.[28]
Распределение
Y-хромосомы гаплогруппы D-M174, которые встречаются среди Тибето-бирманский популяции, а также люди Японского архипелага (гаплогруппы D1a2b, D1a2a и D1a1). D-M55 (D1a2a) являются особенно отличительными, несущими комплекс, по меньшей мере, пяти отдельных мутаций вдоль внутренней ветви гаплогруппы D-M174. филогения, тем самым четко отличая их от других хромосом гаплогруппы D-M174, которые встречаются у тибетцев и жителей Андаманских островов, и предоставляя доказательства того, что гаплогруппа D-M55 Y-хромосомы была модальной гаплогруппой в наследственном населении, которое развило доисторическую эпоху. Jōmon культура на японских островах.
Предполагается, что большинство носителей Y-хромосомы D-M174 мигрировали из Центральная Азия к Восточная Азия. Одна группа переехала в Андаманские острова, таким образом формируя или помогая сформировать Андаманский люди. Другая группа осталась в современном Тибете и на юге Китая (сегодня Тибето-бирманский народов), а другая группа мигрировала в Японию, возможно, через Корейский полуостров (предварительноJmon люди ).[2][18]
Субклады
D-Z27276 (D1a1)
Гаплогруппа D-Z27276 является общим предком D-M15 и D-P99.
D-M15 (D1a1a)
Впервые сообщалось, что D-M15 был обнаружен в образце из Камбоджи и Лаоса (1/18 = 5,6%) и в образце из Японии (1/23 = 4,3%) в предварительном всемирном исследовании вариации Y-ДНК в сохранившиеся человеческие популяции.[29]
Впоследствии Y-ДНК, принадлежащая гаплогруппе D-M15, часто обнаруживалась среди Тибето-бирманский - говорящее население Юго-Западный Китай (в том числе примерно 23% Цян,[2][30][31] примерно 12,5% Тибетцы,[2] и примерно 9% Йи[2][32]) и среди Яо люди населяющий северо-восток Гуанси (6/31 = 19,4% Яо низменности, 5/41 = 12,2% коренных жителей Миен, 3/41 = 7,3% Киммун низменности)[33] с умеренным распространением по всей Центральная Азия, Восточная Азия и континентальный Юго-Восточная Азия (Индокитай ).[2]
Исследование, опубликованное в 2011 году, показало, что D-M15 в 7,8% (4/51) выборки Хмонг Доу и в 3,4% (1/29) выборки Синьмуль с севера Лаос.[33]
D-P47 (D1a1b1)
Часто встречается среди Пуми,[2] Наси,[2] и Тибетцы,[34][2] с умеренным распространением в Центральная Азия.[2] Согласно одному исследованию, у тибетцев частота гаплогруппы D-P47 составляет около 41,31%.[2] Гаплогруппа D-P47 обнаружена в 9% Сиань-Хань.[35]
D-M55 (D1a2a)
Ранее известный как D-M55, D-M64.1 / Page44.1 (D1a2a) часто встречается среди Айны[36] и со средней частотой среди Японский[37] и Рюкюанцы.[37]
Ким и другие. (2011) обнаружили гаплогруппу D-M55 в 2,0% (1/51) образца Пекин Хан и в 1,6% (8/506) пула образцов из Южной Кореи, в том числе 3,3% (3/90) из региона Чолла, 2,4% (2/84) из региона Кёнсан, 1,4% (1/72) из региона Чхунчхон, 1,1% (1/87) из региона Чеджу, 0,9% (1/110) из региона Сеул-Кёнги и 0% (0/63) из региона Канвондо.[16] Молоток и другие. (2006) обнаружили гаплогруппу D-P37.1 в 4,0% (3/75) выборки из Южной Кореи.[18]
Низкие уровни D-M116.1 (субклада D-M55) среди мужчин в настоящее время Тимор (0,2% мужчин),[38] Встречается у 9,5% мужчин из Микронезия (Hammer et al. 2006 г.,[18] считается, что это недавняя примесь из Японии. То есть, D-M116.1 (D1a2a1) обычно считается первичным субкладом D-M64.1 (D1a2a), возможно, в результате Японская военная оккупация Юго-Восточной Азии в течение Вторая Мировая Война.
В соответствии с Мицуру Сакитани, Гаплогруппа D1 прибыла из Средней Азии на север Кюсю через Горный Алтай и Корейский полуостров более чем за 40 000 лет до настоящего времени, и гаплогруппа D-M55 (D1a2a) родилась на Японском архипелаге.[39]
Недавно было подтверждено, что японская ветвь гаплогруппы D-M55 отличается и изолирована от других D-ветвей более 53000 лет. Раскол между D1a, вероятно, произошел в Центральной Азии, в то время как некоторые другие предполагают мгновенный раскол во время происхождения самой гаплогруппы D, поскольку японская ветвь имеет пять уникальных мутаций, не обнаруженных ни в одной другой D-ветви.[8]
(D *) Парагруппа D1a2b (D34637)
D1a2b находится на высоких частотах среди Жители Андаманских островов[2] (особенно Онге (23/23 = 100%), Джарава (4/4 = 100%).[40][26]
D1a *
D1a * (xM15, P99, M55) встречается у некоторых тибетских племен меньшинств в Северо-Восточная Индия (среди которых ставки варьируются от нуля до 65%).[5][41][42][43]
D-M175 *
Базальный D-M174 * (xM15, P47, M55) был обнаружен примерно в 5% случаев Алтайцы.[18] Харьков и другие. обнаружили гаплогруппу D * (xD-M15) в 6,3% (6/96) пула выборок южных алтайцев из трех разных местностей, особенно в Куладе (5/46 = 10,9%) и Кош-Агаче (1/7 = 14%), хотя они не тестировали ни одного маркера субклада D-M55 или D-P99. Харьков и другие. также сообщили об обнаружении Y-ДНК гаплогруппы DE-M1 (xD-M174) у одного жителя Южного Алтая из Бешпельтира (1/43 = 2,3%).[19]
Филогенетика
Филогенетическая история
До 2002 года в академической литературе существовало по крайней мере семь систем именования Филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть, прежде всего, своевременным. В приведенной ниже таблице собраны все эти работы на основе знакового Дерева YCC 2002 года. Это позволяет исследователю, просматривающему ранее опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (стенография) | (α) | (β) | (γ) | (δ) | (е) | (ζ) | (η) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ISOGG 2006 | ISOGG 2007 | ISOGG 2008 | ISOGG 2009 | ISOGG 2010 | ISOGG 2011 | ISOGG 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D-M174 | * | * | * | * | * | * | * | * | D | D | D | D | D | D | D | D | D | D |
| D-M15 | 4 | IV | 3G | 12 | Eu5 | H3 | B | D1 | D1 | D1 | D1 | D1 | D1 | D1 | D1 | D1 | D1 | D1 |
| D-M55 | * | * | * | * | * | * | * | * | D2 | D2 | D2 | D2 | D2 | D2 | D2 | D2 | D2 | D2 |
| D-P12 | 4 | IV | 3G | 11 | Eu5 | H2 | B | D2a | D2a | D2a1a1 | D2a1a1 | D2 | D2 | D2a1a1 | D2a1a1 | D2a1a1 | удаленный | удаленный |
| D-M116.1 | 4 | IV | 3G | 11 | Eu5 | H2 | B | D2b * | D2a | D2a | D2a | D2a | D2a | D2a | D2a | D2a | удаленный | удаленный |
| D-M125 | 4 | IV | 3G | 11 | Eu5 | H2 | B | D2b1 | D2a1 | D2a1 | D2a1 | D2a1 | D2a1 | D2a1 | D2a1 | D2a1 | D2a1 | D2a1 |
| D-M151 | 4 | IV | 3G | 11 | Eu5 | H2 | B | D2b2 | D2a1 | D2a2 | D2a2 | D2a2 | D2a2 | D2a2 | D2a2 | D2a2 | D2a2 | D2a2 |
Научные публикации
Следующие исследовательские группы в соответствии с их публикациями были представлены в создании дерева YCC.
Филогенетические деревья
По дереву ISOGG (Версия: 14.151).[44]
- DE (ЯП)
- D (CTS3946)
- D1 (M174 / страница 30, IMS-JST021355)
- D1a (CTS11577)
- D1a1 (F6251 / Z27276)
- D1a1a (M15)Материковый Китай, Тибет
- D1a1b (P99)Материковый Китай, Тибет, Монгол, Центральная Азия
- D1a2(Z3660)
- D1a2a (M64.1 / Страница 44.1, M55) Япония (Люди Ямато 、Народ рюкюань 、Айны )
- D1a2b (Y34637)Андаманские острова (Онге люди 、Люди Джарава )
- D1a1 (F6251 / Z27276)
- D1b (L1378) Филиппины[45]
- D1a (CTS11577)
- D2 (A5580.2)Нигерия, Саудовская Аравия, Сирия
- D1 (M174 / страница 30, IMS-JST021355)
- D (CTS3946)
Смотрите также
Генетика
Субклады Y-ДНК D-M174
Основное дерево Y-ДНК
| Филогенетическое древо человека Гаплогруппы ДНК Y-хромосомы [χ 1][χ 2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y-хромосомный Адам " | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| грамм | HIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | ЧАС | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| я | J | LT [χ 5] | K2 [χ 6] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| L | Т | K2a [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| K-M2313 [χ 10] | K2b1 [χ 11] | п [χ 12] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| НЕТ | S [χ 13] | M [χ 14] | P1 | P2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| N | О | Q | р | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Рекомендации
- ^ «Гаплогруппа Y-ДНК D-M174 и ее субклады - 2017».
- ^ а б c d е ж грамм час я j k л м п о п Ши Х, Чжун Х, Пэн И, Донг ИЛ, Ци ХБ, Чжан Ф, Лю Л.Ф., Тан С.Дж., Ма Р.З., Сяо СиДжей, Уэллс Р.С., Цзинь Л., Су Б. (октябрь 2008 г.). «Y-хромосома - свидетельство самого раннего современного человеческого поселения в Восточной Азии и множественного происхождения тибетского и японского населения». BMC Биология. 6: 45. Дои:10.1186/1741-7007-6-45. ЧВК 2605740. PMID 18959782.
- ^ а б c YFull Haplogroup YTree v6.02 от 2 апреля 2018 г. По состоянию на 7 июля 2018 г.
- ^ а б c Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF (май 2008 г.). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогруппы Y-хромосомы человека». Геномные исследования. 18 (5): 830–8. Дои:10.1101 / gr.7172008. ЧВК 2336805. PMID 18385274.
- ^ а б Су Би, Сяо К., Дека Р., Сейелстад М. Т., Кангванпонг Д., Сяо Дж., Лу Д., Андерхилл П., Кавалли-Сфорца Л., Чакраборти Р., Джин Л. (декабрь 2000 г.). «Гаплотипы Y-хромосомы показывают доисторические миграции в Гималаи». Генетика человека. 107 (6): 582–90. Дои:10.1007 / s004390000406. PMID 11153912. S2CID 36788262.
- ^ Гайден, Тензин; Кадены, Алисия М .; Регейро, Мария; Сингх, Нанда Б.; Животовский, Лев А .; Андерхилл, Питер А .; Кавалли-Сфорца, Луиджи Л .; Эррера, Рене Дж. (Май 2007 г.). «Гималаи как направленный барьер для потока генов». Американский журнал генетики человека. 80 (5): 884–894. Дои:10.1086/516757. ЧВК 1852741. PMID 17436243.
- ^ Мацумото Х (февраль 2009 г.). «Происхождение японской расы на основе генетических маркеров иммуноглобулина G». Труды Японской академии. Серия B, Физические и биологические науки. 85 (2): 69–82. Bibcode:2009PJAB ... 85 ... 69M. Дои:10.2183 / pjab.85.69. ЧВК 3524296. PMID 19212099.
- ^ а б Мондаль, Маюх; Бергстрём, Андерс; Сюэ, Яли; Калафель, Франсеск; Лааюни, Хафид; Казальс, Ферран; Majumder, Partha P .; Тайлер-Смит, Крис; Бертранпетит, Жауме (01.05.2017). «Последовательности Y-хромосомы разнообразных индийских популяций и предков андаманцев». Генетика человека. 136 (5): 499–510. Дои:10.1007 / s00439-017-1800-0. HDL:10230/34399. ISSN 1432-1203. PMID 28444560. S2CID 3725426.
Напротив, у рианг (тибето-бирманскоязычных) и андаманцев есть свои ближайшие соседи в Восточной Азии. Джарава и Онге разделяли родословные гаплогруппы D друг с другом в течение последних ~ 7000 лет, но расходились с японскими Y-хромосомами гаплогруппы D ~ 53000 лет назад, скорее всего, из-за отделения от общего предкового населения.
- ^ Тайлер-Смит, Крис; Сюэ, Яли; Thomas, Mark G .; Ян, Хуаньминь; Арчиеро, Елена; Асан; Коннелл, Брюс А .; Джонс, Эбигейл Л .; Хабер, Марк (13.06.2019). «Редкая африканская гаплогруппа Y-хромосомы D0 с глубокими корнями и ее значение для распространения современного человека за пределы Африки». Генетика. 212 (4): 1421–1428. Дои:10.1534 / генетика.119.302368. ISSN 0016-6731. ЧВК 6707464. PMID 31196864.
- ^ Эстес, Роберта (21.06.2019). "Захватывающие новые открытия гаплогруппы D ДНК Y!". DNAeXplained - Генетическая генеалогия. Получено 2019-07-08.
- ^ Y 染色体 单 倍 群 D 在 東亞 的 分布 及其 意義
- ^ Чжун Х, Ши Х, Ци ХБ, Дуань Цзы, Тан П.П., Цзинь Л., Су Б, Ма Р.З. (2011). «Расширенное исследование Y-хромосомы предполагает постледниковую миграцию современного человека в Восточную Азию по северному пути». Мол. Биол. Evol. 28 (1): 717–727. Дои:10.1093 / molbev / msq247. PMID 20837606.
- ^ Ян С., Ван СС, Ли Х, Ли С.Л., Джин Л. и др. (Генографический консорциум) (сентябрь 2011 г.). «Обновленное дерево Y-хромосомы гаплогруппы O и пересмотренные филогенетические позиции мутаций P164 и PK4». Европейский журнал генетики человека. 19 (9): 1013–5. Дои:10.1038 / ejhg.2011.64. ЧВК 3179364. PMID 21505448.
- ^ а б Пак MJ, Ли Х.Й., Ян В.И., Шин KJ (июль 2012 г.). «Понимание вариации Y-хромосомы в Корее - актуальность комбинированного анализа гаплогруппы и гаплотипа». Международный журнал судебной медицины. 126 (4): 589–99. Дои:10.1007 / s00414-012-0703-9. PMID 22569803. S2CID 27644576.
- ^ а б c d Като Т., Мункхбат Б., Тунаи К., Мано С., Андо Х, Оюнгерел Г., Чаэ Г. Т., Хан Х., Цзя Г.Дж., Токунага К., Мунхтувшин Н., Тамия Г., Иноко Х (февраль 2005 г.). «Генетические особенности монгольских этносов, выявленные с помощью анализа Y-хромосомы». Ген. 346: 63–70. Дои:10.1016 / j.gene.2004.10.023. PMID 15716011.
- ^ а б c d е ж грамм Ким, Сун-Хи; Ким, Ки-Чхоль; Шин, Донг-Джик; Джин, Хан-Джун; Квак, Кён-Дон; Хан, Мюн-Су; Сон, Джун-Мён; Ким, Вон; Ким, Ук (2011). «Высокие частоты линий гаплогруппы O2b-SRY465 Y-хромосомы в Корее: генетическая перспектива заселения Кореи». Следственная генетика. 2 (1): 10. Дои:10.1186/2041-2223-2-10. ЧВК 3087676. PMID 21463511.
- ^ а б c Уэллс Р.С., Юлдашева Н., Рузибакиев Р., Андерхилл П.А., Евсеева И., Блю-Смит Дж., Джин Л., Су Б., Питчаппан Р., Шанмугалакшми С., Балакришнан К., Рид М., Пирсон Н. М., Зерджал Т., Вебстер М. Т., Жолошвили И., Джамарджашвили Э., Гамбаров С., Никбин Б., Достиев А., Акназаров О., Заллоуа П., Цой И., Китаев М., Миррахимов М., Чариев А., Бодмер В. Ф. (август 2001 г.). «Сердце Евразии: континентальный взгляд на разнообразие Y-хромосомы». Труды Национальной академии наук Соединенных Штатов Америки. 98 (18): 10244–9. Bibcode:2001PNAS ... 9810244W. Дои:10.1073 / pnas.171305098. ЧВК 56946. PMID 11526236.
- ^ а б c d е ж грамм час Hammer MF, Karafet TM, Park H, Omoto K, Harihara S, Stoneking M, Horai S (2006). «Двойное происхождение японцев: общая основа для Y-хромосом охотников-собирателей и фермеров». Журнал генетики человека. 51 (1): 47–58. Дои:10.1007 / s10038-005-0322-0. PMID 16328082.
- ^ а б c Харьков В.Н., Степанов В.А., Медведева О.Ф., Спиридонова М.Г., Воевода М.И., Тадинова В.Н., Пузырев В.П. (май 2007 г.). «[Различия генофонда северных и южных алтайцев, выведенные из данных о гаплогруппах Y-хромосомы]». Генетика (на русском). 43 (5): 675–87. Дои:10.1134 / S1022795407050110. PMID 17633562. S2CID 566825.
- ^ а б Дулик М.С., Жаданов С.И., Осипова Л.П., Аскапули А., Гау Л., Гоккумен О., Рубинштейн С., Шурр Т.Г. (февраль 2012 г.). «Митохондриальная ДНК и вариации Y-хромосомы свидетельствуют о недавнем общем происхождении коренных американцев и коренных алтайцев». Американский журнал генетики человека. 90 (2): 229–46. Дои:10.1016 / j.ajhg.2011.12.014. ЧВК 3276666. PMID 22281367.
- ^ Е.Е. Аширбеков, Д.М. Ботбаев, А.М. Белкожаев, А.О. Абайлдаев, А.С. Неупокоева, Ю.Е. Мухатаев, Б. Альжанулы, Д.А. Шарафутдинова, Д.Д. Мукушкина, М.Б. Рахымгожин, А.К. Хансеитова, С.А. Лимборска, Н.А. Айтхожожина. Казахи из Южно-Казахстанской, Жамбылской и Алматинской областей ». Доклады Национальной академии наук Республики Казахстан, ISSN 2224-5227, Volume 6, Number 316 (2017), 85 - 95.
- ^ а б Юнусбаев Б., Мецпалу М., Ярве М., Кутуев И., Роотси С., Мецпалу Е., Бехар Д.М., Варенди К., Саакян Х., Хусаинова Р., Епископосян Л., Хуснутдинова Е.К., Андерхилл П.А., Кивисилд Т., Виллемс Р. (январь 2012 г.). «Кавказ как асимметричный полупроницаемый барьер для древних миграций человека». Молекулярная биология и эволюция. 29 (1): 359–65. Дои:10.1093 / molbev / msr221. PMID 21917723.
- ^ Малярчук Б., Деренко М., Денисова Г., Хойт С., Возняк М., Гжибовски Т., Захаров И. (декабрь 2013 г.). «Разнообразие Y-хромосомы у калмыков на этническом и племенном уровнях». Журнал генетики человека. 58 (12): 804–11. Дои:10.1038 / jhg.2013.108. PMID 24132124.
- ^ Y-ДНК D Проект гаплогруппы в генеалогическом древе ДНК
- ^ Ди Кристофаро Дж., Пеннарун Э., Мазьер С., Майрес Н.М., Лин А.А., Темори С.А., Мецпалу М., Мецпалу Э., Витцель М., Кинг Р.Дж., Андерхилл ПА, Виллемс Р., Кьярони Дж. (2013). «Афганский Гиндукуш: там, где сходятся генные потоки Евразийского субконтинента». PLOS ONE. 8 (10): e76748. Bibcode:2013PLoSO ... 876748D. Дои:10.1371 / journal.pone.0076748. ЧВК 3799995. PMID 24204668.
- ^ а б Y-полный
- ^ а б Су, Бинг; Джин, Ли; Андерхилл, Питер; Мартинсон, Джереми; Саха, Нилмани; МакГарви, Стивен Т .; Шрайвер, Марк Д .; Чу, Цзяю; Эфнер, Питер; Чакраборти, Ранаджит; Дека, Ранджан (18 июля 2000 г.). «Полинезийское происхождение: выводы из Y-хромосомы». Труды Национальной академии наук Соединенных Штатов Америки. 97 (15): 8225–8228. Bibcode:2000PNAS ... 97.8225S. Дои:10.1073 / пнас.97.15.8225. ЧВК 26928. PMID 10899994.
- ^ ДОНГ Юнли, ШИ Хун, Ли Вэйсян, Ян Цзе, Цзэн Вэйминь, Ли Кайюань и Сяо Чунцзе, «Изучение полиморфизма в локусе YAP в семи популяциях этнических меньшинств Юньнани в большом ущелье реки Салуин и в нижнем течении» (оригинал название на китайском языке: "怒江 大 峡谷 及 下游 地区 7 个 云南 少数民族 YAP 位 点 的 多态性 研究"), Acta Anthropologica Sinica, Vol. 21, № 3 (август 2002 г.). http://www.ivpp.cas.cn/cbw/rlxxb/xbwzxz/201203/t20120320_3512811.html
- ^ Андерхилл П.А., Шен П., Линь А.А., Джин Л., Пассарино Дж., Ян У.Х., Кауфман Э, Бонне-Тамир Б., Бертранпетит Дж., Франкалаччи П., Ибрагим М., Дженкинс Т., Кидд Дж. Р., Мехди С. К., Зайелстад М. Т., Уэллс Р. Пьяцца А, Дэвис Р.В., Фельдман М.В., Кавалли-Сфорца Л.Л., Эфнер П.Дж. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история популяций человека». Природа Генетика. 26 (3): 358–61. Дои:10.1038/81685. PMID 11062480. S2CID 12893406.
- ^ Сюэ И, Зерджал Т., Бао В., Чжу С., Шу Кью, Сюй Дж, Ду Р, Фу С., Ли П, Хёрлс М.Э., Ян Х., Тайлер-Смит К. (апрель 2006 г.). «Демография мужчин в Восточной Азии: контраст между севером и югом во временах роста населения». Генетика. 172 (4): 2431–9. Дои:10.1534 / генетика.105.054270. ЧВК 1456369. PMID 16489223.
- ^ Ван СС, Ван LX, Шреста Р., Чжан М., Хуанг XY, Ху К., Цзинь Л., Ли Х (2014). «Генетическая структура популяций Qiangic, проживающих в западном коридоре Сычуань». PLOS ONE. 9 (8): e103772. Bibcode:2014PLoSO ... 9j3772W. Дои:10.1371 / journal.pone.0103772. ЧВК 4121179. PMID 25090432.
- ^ Вэнь Би, Се Х, Гао С., Ли Х, Ши Х, Сон Х, Цянь Т, Сяо С., Джин Дж, Су Би, Лу Ди, Чакраборти Р., Джин Л. (май 2004 г.). «Анализ генетической структуры тибето-бирманских популяций выявляет гендерно-предвзятую примесь южных тибето-бирманцев». Американский журнал генетики человека. 74 (5): 856–65. Дои:10.1086/386292. ЧВК 1181980. PMID 15042512.
- ^ а б Цай X, Цинь З., Вэнь Б., Сюй С., Ван И, Лу И, Вэй Л., Ван С., Ли С., Хуанг Х, Цзинь Л., Ли Х (2011). «Миграция людей через узкие места из Юго-Восточной Азии в Восточную Азию во время последнего ледникового максимума, выявленного Y-хромосомами». PLOS ONE. 6 (8): e24282. Bibcode:2011PLoSO ... 624282C. Дои:10.1371 / journal.pone.0024282. ЧВК 3164178. PMID 21904623.
- ^ Дуншэн Лу и другие. Предки и генетическая история тибетских горцев, 25 августа 2016 г.
- ^ Ким 2011
- ^ Таджима А., Хаями М., Токунага К., Джуджи Т., Мацуо М., Марзуки С., Омото К., Хораи С. (2004). «Генетическое происхождение айнов установлено на основе комбинированного анализа ДНК материнской и отцовской линий». Журнал генетики человека. 49 (4): 187–93. Дои:10.1007 / s10038-004-0131-x. PMID 14997363.
- ^ а б ЮИЧИ САТО, ТОСИКАЦУ ШИНКА, АШРАФ А. ЭВИС, АЙКО ЯМАУТИ, ТЕРУАКИ ИВАМОТО, ЮТАКА НАКАХОРИ Обзор генетической изменчивости Y-хромосомы современных японских мужчин.
- ^ Тумонггор, Мерианн К.; Карафет, Татьяна М; Дауни, Шон; Лансинг, Дж. Стивен; Норквест, Питер; Судойо, Херавати; Хаммер, Майкл Ф; Кокс, Мюррей П. (сентябрь 2014 г.). «Изоляция, контакты и социальное поведение сформировали генетическое разнообразие в Западном Тиморе». Журнал генетики человека. 59 (9): 494–503. Дои:10.1038 / jhg.2014.62. ЧВК 4521296. PMID 25078354.
- ^ 崎 谷 満 『DNA ・ 考古 ・ 言語 の 学 が 示 す 新 ・ 日本 列島』 (勉 誠 出 2009) (на японском языке)
- ^ Thangaraj K, Singh L, Reddy AG, Rao VR, Sehgal SC, Underhill PA, Pierson M, Frame IG, Hagelberg E (январь 2003 г.). «Генетическая близость жителей Андаманских островов, исчезающая человеческая популяция». Текущая биология. 13 (2): 86–93. Дои:10.1016 / S0960-9822 (02) 01336-2. PMID 12546781. S2CID 12155496.
- ^ Cordaux R, Weiss G, Saha N, Stoneking M (август 2004 г.). «Северо-восточный индийский коридор: барьер или коридор для миграции людей?». Молекулярная биология и эволюция. 21 (8): 1525–33. Дои:10.1093 / молбев / мш151. PMID 15128876.
- ^ Чандрасекар А., Сахеб С.Ю., Гангопадья П., Гангопадья С., Мукерджи А., Басу Д., Лакшми Г.Р., Сахани А.К., Дас Б., Баттачарья С., Кумар С., Хавиур Д., Сан Д., Рао В.Р. (2007). «Подпись для вставки YAP в Южной Азии». Анналы биологии человека. 34 (5): 582–6. Дои:10.1080/03014460701556262. PMID 17786594. S2CID 11860142.
- ^ Редди Б.М., Лангстих Б.Т., Кумар В., Нагараджа Т., Редди А.Н., Мека А., Редди А.Г., Тангарадж К., Сингх Л. (ноябрь 2007 г.). «Австро-азиатские племена Северо-Восточной Индии обеспечивают до сих пор отсутствующую генетическую связь между Южной и Юго-Восточной Азией». PLOS ONE. 2 (11): e1141. Bibcode:2007PLoSO ... 2.1141R. Дои:10.1371 / journal.pone.0001141. ЧВК 2065843. PMID 17989774.
- ^ Y-ДНК Гаплогруппа D и ее субклады - 2019
- ^ Y-ДНК Гаплогруппа D и ее субклады - 2014
- Андерхилл ПА, Кивисилд Т. (2007). «Использование структуры популяции y-хромосомы и митохондриальной ДНК в отслеживании миграции людей». Ежегодный обзор генетики. 41: 539–64. Дои:10.1146 / annurev.genet.41.110306.130407. PMID 18076332. S2CID 24904955.
внешняя ссылка
- Атлас пути человека: генетические маркеры, гаплогруппа D-M174 (M174), из Генографический проект на Национальная география
- Знаменитая ДНК Японии