Генипавирус - Henipavirus
| Генипавирус | |
|---|---|
 | |
| Цветные трансмиссионная электронная микрофотография из Hendra henipavirus вирион (длина около 300 нм) | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область: | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Негарнавирикота |
| Учебный класс: | Monjiviricetes |
| Заказ: | Mononegavirales |
| Семья: | Paramyxoviridae |
| Подсемейство: | Orthoparamyxovirinae |
| Род: | Генипавирус |
| Типовой вид | |
| Hendra henipavirus | |
| Разновидность | |
Генипавирус это род из вирусы с отрицательной цепью РНК в семье Paramyxoviridae, порядок Mononegavirales содержащий пять видов.[1][2] Генипавирусы естественным образом питают птеропид фруктовые летучие мыши (летучие лисицы) и микробаты нескольких видов.[3] Хенипавирусы характеризуются длительным геномы и широкий выбор хостов. Их недавнее появление как зоонозный патогены, способные вызвать болезнь и смерть у домашние животные и люди вызывают беспокойство.[4][5]
В 2009 году последовательности РНК трех новых вирусов в филогенетическом родстве с известными вирусами генипавируса были обнаружены у плодовых летучих мышей африканского соломенного цвета (Eidolon helvum ) в Гана. Обнаружение этих новых вирусов генипавируса за пределами Австралии и Азии указывает на то, что регион потенциальной эндемичности вирусов генипавируса может быть во всем мире.[6] Эти африканские вирусы генипавируса медленно исследуются.[7]
Структура


Генипавирионы являются плеоморфный (различной формы), размером от 40 до 600 нм в диаметре.[8] Они обладают липид мембрана, покрывающая оболочку вирусного матричный белок. В основе лежит одна спиральная нить геномного РНК тесно связана с N (нуклеокапсид ) белок и связанный с белками L (большой) и P (фосфопротеин), которые обеспечивают РНК-полимераза активность во время репликации.
В липидную мембрану встроены шипы тримеров F (слитого) белка и тетрамеров G (присоединения). Функция белка G (за исключением MojV-G) заключается в прикреплении вируса к поверхности клетки-хозяина через Эфрин B1, B2 или B3, семья очень консервированный белки млекопитающих.[9][10][11] Структура прикрепительного гликопротеина была определена с помощью рентгеновской кристаллографии.[12] Белок F сливает вирусную мембрану с мембраной клетки-хозяина, высвобождая содержимое вириона в клетку. Это также заставляет инфицированные клетки сливаться с соседними клетками, образуя большие многоядерные синцития.
Геном
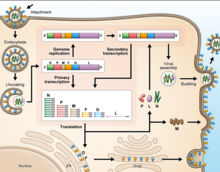
Как и все мононегавирусные геномы, геномы вируса Хендра и вируса Нипах несегментированы, одноцепочечные. отрицательный смысл РНК. Оба генома имеют длину 18,2 т.п.н. и содержат шесть гены соответствующих шести структурным белкам.[13]
Как и другие участники Paramyxoviridae семья, количество нуклеотиды в геноме вируса генипавируса кратно шести, что соответствует так называемому "правило шести '.[14] Отклонение от правила шести из-за мутации или неполного синтеза генома приводит к неэффективной репликации вируса, вероятно, из-за структурных ограничений, налагаемых связыванием между РНК и N-белком.
Генипавирусы используют необычный процесс, называемый Редактирование РНК для создания нескольких белков из одного гена. Специфический процесс у вирусов генипавируса включает введение дополнительных гуанозин остатки в гене P мРНК до перевод. Количество добавленных остатков определяет, синтезируются ли белки P, V C или W. Функции белков V и W неизвестны, но они могут участвовать в нарушении антивирусных механизмов хозяина.
Причины появления
Появление вирусов генипавируса происходит параллельно с появлением других зоонозный вирусы в последние десятилетия. SARS коронавирус, Лиссавирус австралийских летучих мышей, Вирус Менангле и вероятно Вирус Эбола и Марбургский вирус также являются укрытием для летучих мышей и способны заразить множество других видов. Появление каждого из этих вирусов было связано с увеличением контактов между летучими мышами и людьми, иногда с участием промежуточных домашних животных-хозяев. Усиление контактов обусловлено как вторжением человека на территорию летучих мышей (в случае Нипах, в частности, свиные загоны на указанной территории), так и перемещением летучих мышей к человеческому населению из-за изменений в распределении пищи и утраты среды обитания.
Есть свидетельства того, что утрата местообитаний летучих лисиц как в Южной Азии, так и в Австралии (особенно вдоль восточного побережья), а также вторжение человеческих жилищ и сельскохозяйственных угодий в оставшиеся места обитания приводит к большему дублированию распространения людей и летучих лисиц.
Таксономия
| Род | Разновидность | Вирус (аббревиатура) |
| Генипавирус | Кедровый генипавирус | Кедровый вирус (CedV) |
| Ганский летучая мышь henipavirus | Кумаси вирус (КВ) | |
| Hendra henipavirus * | Вирус Хендры (HeV) | |
| Хенипавирус Mojiang | Вирус Mòjiāng (MojV) | |
| Нипах генипавирус | Вирус нипах (NiV) | |
| Легенда таблицы: "*" обозначает типовые виды. | ||
Смотрите также
Рекомендации
- ^ Рима, Б; Балкема-Бушманн, А; Дандон, WG; Duprex, P; Истон, А; Фушье, Р. Курат, G; Lamb, R; Ли, Б; Рота, П; Ван, L; Консорциум отчетов ICTV (декабрь 2019 г.). "Профиль таксономии вирусов ICTV: Paramyxoviridae". Журнал общей вирусологии. 100 (12): 1593–1594. Дои:10.1099 / jgv.0.001328. PMID 31609197.
- ^ "Отчет ICTV Paramyxoviridae".
- ^ Ли, У; Ван, Дж; Хики, AC; Zhang, Y; Ли, У; Wu, Y; Чжан, Хуацзюнь; и другие. (Декабрь 2008 г.). «Антитела к Nipah или Nipah-подобным вирусам у летучих мышей, Китай [письмо]». Возникающие инфекционные заболевания. 14 (12): 1974–6. Дои:10.3201 / eid1412.080359. ЧВК 2634619. PMID 19046545.
- ^ Саватский (2008). «Хендра и Нипах Вирус». Вирусы животных: молекулярная биология. Caister Academic Press. ISBN 978-1-904455-22-6.
- ^ «Нипах еще не подтвержден, 86 под наблюдением: Шайладжа». OnManorama. Получено 4 июн 2019.
- ^ Дрекслер Дж. Ф., Корман В. М., Глоза-Рауш Ф., Зеебенс А., Аннан А. (2009). Маркоттер У (ред.). «РНК генипавируса африканских летучих мышей». PLOS One. 4 (7): e6367. Bibcode:2009PLoSO ... 4.6367D. Дои:10.1371 / journal.pone.0006367. ЧВК 2712088. PMID 19636378.
- ^ Дрекслер Дж. Ф., Корман В. М.; и другие. (2012). «Летучие мыши являются носителями основных парамиксовирусов млекопитающих». Nat Commun. 3: 796. Дои:10.1038 / ncomms1796.
- ^ Hyatt AD, Zaki SR, Goldsmith CS, Wise TG, Hengstberger SG (2001). «Ультраструктура вируса Хендра и вируса Нипах в культивируемых клетках и животных-хозяевах». Микробы и инфекции. 3 (4): 297–306. Дои:10.1016 / S1286-4579 (01) 01383-1. PMID 11334747.
- ^ Бонапарт, М; Димитров А; Боссарт, К. (2005). «Лиганд эфрина-B2 является функциональным рецептором вируса Хендра и вируса Нипах». Труды Национальной академии наук. 102 (30): 10652–7. Bibcode:2005ПНАС..10210652Б. Дои:10.1073 / pnas.0504887102. ЧВК 1169237. PMID 15998730.
- ^ Негрете О.А., Левруни Е.Л., Агилар ХК (2005). «EphrinB2 является рецептором входа для вируса Nipah, возникающего смертельного парамиксовируса». Природа. 436 (7049): 401–5. Bibcode:2005Натура.436..401Н. Дои:10.1038 / природа03838. PMID 16007075.
- ^ Bowden, Thomas A .; Криспин, Макс; Джонс, Э. Ивонн; Стюарт, Дэвид I. (1 октября 2010 г.). «Общая архитектура парамиксовирусных гликопротеинов адаптирована для различных стратегий прикрепления». Сделки Биохимического Общества. 38 (5): 1349–1355. Дои:10.1042 / BST0381349. ЧВК 3433257. PMID 20863312.
- ^ Bowden, Thomas A .; Криспин, Макс; Харви, Дэвид Дж .; Арическу, А. Раду; Граймс, Джонатан М .; Джонс, Э. Ивонн; Стюарт, Дэвид I. (1 декабря 2008 г.). «Кристаллическая структура и анализ углеводов гликопротеина прикрепления вируса Нипах: шаблон для разработки противовирусных препаратов и вакцин». Журнал вирусологии. 82 (23): 11628–11636. Дои:10.1128 / JVI.01344-08. ЧВК 2583688. PMID 18815311.
- ^ Ван Л., Харкорт Б. Х., Ю. М. (2001). «Молекулярная биология вирусов Хендра и Нипах». Микробы и инфекции. 3 (4): 279–87. Дои:10.1016 / S1286-4579 (01) 01381-8. PMID 11334745.
- ^ Колаковский, Д; Пелет, Т; Гарсин, Д; Hausmann, S; Курран, Дж; Ру, Л. (февраль 1998 г.). «Синтез РНК парамиксовируса и требования к длине гексамерного генома: пересмотренное правило шести». Журнал вирусологии. 72 (2): 891–9. Дои:10.1128 / JVI.72.2.891-899.1998. ЧВК 124558. PMID 9444980.
- ^ Amarasinghe, Gaya K .; Бао, Иминь; Basler, Christopher F .; Бавари, Сина; Пиво, Мартин; Бежерман, Николас; Blasdell, Kim R .; Бохновски, Алиса; Бризе, Томас (7 апреля 2017 г.). «Таксономия отряда Mononegavirales: обновление 2017 г.». Архив вирусологии. 162 (8): 2493–2504. Дои:10.1007 / s00705-017-3311-7. ISSN 1432-8798. ЧВК 5831667. PMID 28389807.
внешняя ссылка
- Отчет ICTV: Paramyxoviridae
- Карта болезни
- ViralZone: генипавирус
- Генипавирус - Группа исследования экологии генипавируса (HERG) INFO
- Ресурс базы данных и анализа вирусных патогенов (ViPR): Paramyxoviridae