Гликогенфосфорилаза - Glycogen phosphorylase
эта статья нужны дополнительные цитаты для проверка. (Январь 2009 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |
| Фосфорилаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
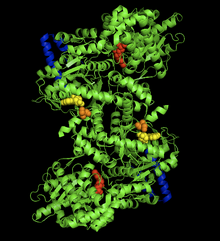 Кристаллическая структура комплекса гликогенфосфорилаза-АМФ в мышцах кролика. Аллостерический сайт AMP (желтый), фосфорилированный Ser14 (оранжевый), сайт связывания гликогена (синий), каталитический сайт (красный).[1] | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 2.4.1.1 | ||||||||
| Количество CAS | 9035-74-9 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
Гликогенфосфорилаза один из фосфорилаза ферменты (EC 2.4.1.1 ). Гликогенфосфорилаза катализирует лимитирующую стадию в гликогенолиз у животных путем выпуска глюкозо-1-фосфат от концевой альфа-1,4-гликозидной связи. Гликогенфосфорилаза также изучается как модельный белок, регулируемый как обратимыми, так и обратимыми фосфорилирование и аллостерический эффекты.
Механизм
Гликогенфосфорилаза расщепляется гликоген в глюкоза подразделения (см. также рисунок ниже):
(цепь гликогена α-1,4)п + Пи ⇌ (цепь гликогена α-1,4)п-1 + α-D-глюкозо-1-фосфат.[2]
Гликогена остается на одну глюкозу меньше молекула, а свободная молекула глюкозы имеет вид глюкозо-1-фосфат. Чтобы использовать для метаболизм, он должен быть преобразован в глюкозо-6-фосфат ферментом фосфоглюкомутаза.
Хотя реакция обратимая in vitro, внутри клетки фермент работает только в прямом направлении, как показано ниже, потому что концентрация неорганический фосфат намного выше, чем у глюкозо-1-фосфата.[2]
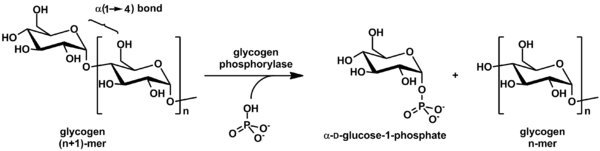
Гликогенфосфорилаза может действовать только на линейный цепи гликогена (α1-4 гликозидная связь). Его работа немедленно остановится в четырех остатках от α1-6. ветвь (которые чрезвычайно распространены в гликогене). В этих ситуациях расщепляющий фермент необходимо, чтобы распрямить цепь в этой области. Кроме того, фермент трансфераза сдвигает блок из 3 глюкозильных остатков с внешней ветви на другой конец, а затем α1-6 глюкозидаза фермент требуется, чтобы разорвать оставшийся (единственный глюкозный) остаток α1-6, который остается в новой линейной цепи. После того, как все это будет сделано, гликогенфосфорилаза может продолжаться. Фермент специфичен для α1-4 цепей, так как молекула содержит щель длиной 30 ангстрем с таким же радиусом, что и спираль, образованная цепью гликогена; это вмещает 4-5 глюкозильных остатков, но слишком мало для разветвлений. Эта щель соединяет сайт хранения гликогена с активным каталитическим сайтом.
Гликогенфосфорилаза имеет пиридоксальфосфат (PLP, полученный из Витамин B6 ) на каждом каталитическом сайте. Пиридоксальфосфат связывается с основными остатками (в данном случае Lys680) и ковалентно образует База Шиффа. После образования связи основания Шиффа, удерживающего молекулу PLP в активном центре, фосфатная группа на PLP легко отдает протон молекуле неорганического фосфата, позволяя неорганическому фосфату, в свою очередь, депротонироваться кислородом, образующим α-1 , 4 гликозидная связь. PLP легко депротонируется, потому что его отрицательный заряд не только стабилизируется внутри фосфатной группы, но также в пиридиновом кольце, таким образом, основание конъюгата, образующееся в результате депротонирования PLP, довольно стабильно. Протонированный кислород теперь представляет собой хороший уходящая группа, и цепь гликогена отделена от концевого гликогена в SN1 мода, приводящая к образованию молекулы глюкозы с вторичным карбокатионом в положении 1. Наконец, депротонированный неорганический фосфат действует как нуклеофил и связывается с карбокатионом, что приводит к образованию глюкозо-1-фосфата и цепи гликогена, укороченной на одну молекулу глюкозы.
Существует также альтернативный предложенный механизм с участием положительно заряженного кислорода в конформации полукресла.[3]

Структура
Мономер гликогенфосфорилазы - это большой белок, состоящий из 842 аминокислот с массой 97,434 кДа в мышечных клетках. Хотя фермент может существовать в виде неактивного мономера или тетрамера, он биологически активен как димер из двух одинаковых субъединиц.[4]

У млекопитающих основная изоферменты гликогенфосфорилазы обнаруживаются в мышцах, печени и головном мозге. Тип мозга преобладает во взрослом мозге и эмбриональных тканях, тогда как печень и мышечный тип преобладают в печени и скелетных мышцах взрослых, соответственно.[5]
Димер гликогенфосфорилазы имеет много областей биологического значения, включая каталитический сайты, сайты связывания гликогена, аллостерический сайтов и обратимо фосфорилированный остаток серина. Во-первых, каталитические центры относительно скрыты, на расстоянии 15 Å от поверхности белка и от границы раздела субъединиц.[6] Это отсутствие легкого доступа каталитического сайта к поверхности является значительным, поскольку оно делает активность белка очень чувствительной к регуляции, поскольку небольшие аллостерические эффекты могут значительно увеличить относительный доступ гликогена к сайту.
Пожалуй, самый важный регулирующий сайт это Ser14, сайт обратимого фосфорилирование очень близко к интерфейсу субблока. Структурное изменение, связанное с фосфорилированием и превращением фосфорилазы b в фосфорилазу a, представляет собой расположение первоначально неупорядоченных остатков с 10 по 22 в α-спирали. Это изменение увеличивает активность фосфорилазы до 25% даже в отсутствие AMP и дополнительно усиливает активацию AMP.[7]
Аллостерический сайт AMP Связывание с мышечными изоформами гликогенфосфорилазы близко к границе раздела субъединиц, как и Ser14. Связывание AMP на этом сайте, соответствующее изменению из состояния T фермента в состояние R, приводит к небольшим изменениям в третичной структуре на границе раздела субъединиц, что приводит к большим изменениям в четвертичной структуре.[8] Связывание AMP поворачивает башни-спирали (остатки 262-278) двух субъединиц на 50 ° относительно друг друга за счет большей организации и межсубъединичных взаимодействий. Это вращение башенных спиралей приводит к повороту двух субъединиц на 10 ° относительно друг друга и, что более важно, нарушает остатки 282-286 (петля 280s), которые блокируют доступ к каталитическому сайту в Т-состоянии, но не в состояние R.[6]
Последний, возможно, самый любопытный участок белка гликогенфосфорилазы - это так называемый участок хранения гликогена. Остатки 397-437 образуют эту структуру, которая позволяет белку ковалентно связываться с цепью гликогена на полных 30 Å от каталитического сайта. Этот сайт, скорее всего, является сайтом, в котором фермент связывается с гранулами гликогена перед инициацией расщепления концевых молекул глюкозы. Фактически, 70% димерной фосфорилазы в клетке существует как связанная с гранулами гликогена, а не свободно плавающая.[9]
Клиническое значение
| фосфорилаза, гликоген; мышца (синдром Макардла, болезнь накопления гликогена V типа) | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Символ | PYGM | ||||||
| Ген NCBI | 5837 | ||||||
| HGNC | 9726 | ||||||
| OMIM | 608455 | ||||||
| RefSeq | NM_005609 | ||||||
| UniProt | P11217 | ||||||
| Прочие данные | |||||||
| Номер ЕС | 2.4.1.1 | ||||||
| Locus | Chr. 11 q12-q13.2 | ||||||
| |||||||
| фосфорилаза, гликоген; печень (болезнь Херса, болезнь накопления гликогена VI типа) | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Символ | PYGL | ||||||
| Ген NCBI | 5836 | ||||||
| HGNC | 9725 | ||||||
| OMIM | 232700 | ||||||
| RefSeq | NM_002863 | ||||||
| UniProt | P06737 | ||||||
| Прочие данные | |||||||
| Номер ЕС | 2.4.1.1 | ||||||
| Locus | Chr. 14 q11.2-24.3 | ||||||
| |||||||
| фосфорилаза, гликоген; мозг | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Символ | PYGB | ||||||
| Ген NCBI | 5834 | ||||||
| HGNC | 9723 | ||||||
| OMIM | 138550 | ||||||
| RefSeq | NM_002862 | ||||||
| UniProt | P11216 | ||||||
| Прочие данные | |||||||
| Номер ЕС | 2.4.1.1 | ||||||
| Locus | Chr. 20 п11.2-п11.1 | ||||||
| |||||||
Ингибирование гликогенфосфорилазы было предложено в качестве одного из методов лечения диабет 2 типа.[10] Поскольку было показано, что производство глюкозы в печени увеличивается у пациентов с диабетом 2 типа,[11] подавление высвобождения глюкозы из запасов гликогена в печени, по-видимому, является действенным подходом. Клонирование гликогенфосфорилазы печени человека (HLGP) выявило новый аллостерический сайт связывания рядом с интерфейсом субъединиц, который не присутствует в гликогенфосфорилазе мышц кролика (RMGP), обычно используемой в исследованиях. Этот сайт не был чувствителен к тем же ингибиторам, что и аллостерический сайт AMP,[12] и наибольший успех был достигнут при синтезе новых ингибиторов, имитирующих структуру глюкозы, поскольку глюкозо-6-фосфат является известным ингибитором HLGP и стабилизирует менее активное Т-состояние.[13] Эти производные глюкозы имели некоторый успех в ингибировании HLGP с прогнозируемыми значениями Ki всего 0,016 мМ.[14]
Мутации в мышечной изоформе гликогенфосфорилазы (PYGM) связаны с болезнь накопления гликогена типа V (GSD V, болезнь Макардла). На сегодняшний день идентифицировано более 65 мутаций в гене PYGM, которые приводят к болезни Макардла.[15][16] Симптомы болезни Макардла включают мышечную слабость, миалгия и недостаток выносливости, все проистекающие из низкого уровня глюкозы в мышечной ткани.[17]
Мутации в изоформе гликогенфосфорилазы (PYGL) печени связаны с Ее болезнь (болезнь накопления гликогена типа VI ).[18][19] Ее болезнь часто сопровождается легкими симптомами, обычно ограниченными гипогликемия, и иногда его трудно диагностировать из-за остаточной активности ферментов.[20]
Изоформа гликогенфосфорилазы (PYGB) мозга была предложена в качестве биомаркер для рак желудка.[21]
Регулирование
Гликогенфосфорилаза регулируется через аллостерический контроль и через фосфорилирование. Каждая из фосфорилазы а и фосфорилазы b существует в двух формах: Т (напряженное) неактивное состояние и R (расслабленное) состояние. Фосфорилаза b обычно находится в состоянии T, неактивна из-за физиологического присутствия АТФ и глюкозо-6 фосфата, а фосфорилаза a обычно находится в состоянии R (активна). Изофермент гликогенфосфорилазы существует в печени, чувствительной к концентрации глюкозы, так как печень действует как экспортер глюкозы. По сути, фосфорилаза печени реагирует на глюкозу, что вызывает очень чувствительный переход от R к T-форме, инактивируя ее; кроме того, фосфорилаза печени нечувствительна к АМФ.
Гормоны, такие как адреналин, инсулин и глюкагон регулируют гликогенфосфорилазу с помощью систем амплификации второго мессенджера, связанных с G белки. Глюкагон активирует аденилатциклазу через G-белок-связанный рецептор (GPCR) в сочетании с гs что, в свою очередь, активирует аденилатциклаза для увеличения внутриклеточной концентрации цАМФ. цАМФ связывается и активирует протеинкиназа А (ПКА). Фосфорилаты PKA фосфорилаза киназа, которая, в свою очередь, фосфорилирует гликогенфосфорилазу b по Ser14, превращая ее в активную гликогенфосфорилазу a.
В печени, глюкагон также активирует другой GPCR, который запускает другой каскад, что приводит к активации фосфолипазы C (PLC). PLC косвенно вызывает высвобождение кальция из эндоплазматического ретикулума гепатоцитов в цитозоль. Повышенная доступность кальция связывается с кальмодулин субъединица и активирует киназу гликогенфосфорилазы. Киназа гликогенфосфорилазы активирует гликогенфосфорилазу таким же образом, как упоминалось ранее.
Гликогенфосфорилаза b не всегда неактивна в мышцах, поскольку она может аллостерически активироваться AMP. Повышение концентрации АМФ во время физических упражнений сигнализирует о потребности в энергии. АМФ активирует гликогенфосфорилазу b, изменяя ее конформацию с напряженной на расслабленную. Эта расслабленная форма имеет такие же ферментативные свойства, что и фосфорилированный фермент. Повышение концентрации АТФ препятствует этой активации, вытесняя АМФ из сайта связывания нуклеотидов, что указывает на наличие достаточных запасов энергии.
После еды происходит высвобождение инсулин, сигнализирующий о наличии глюкозы в крови. Инсулин косвенно активирует протеинфосфатаза 1 (PP1) и фосфодиэстераза через каскад передачи сигнала. PP1 дефосфорилирует гликогенфосфорилазу a, реформируя неактивную гликогенфосфорилазу b. Фосфодиэстераза превращает цАМФ в АМФ. Вместе они снижают концентрацию цАМФ и ингибируют ПКА. В результате PKA больше не может инициировать каскад фосфорилирования, который заканчивается образованием (активной) гликогенфосфорилазы a. В целом, передача сигналов инсулина снижает гликогенолиз, чтобы сохранить запасы гликогена в клетке и запускает гликогенез.[22]
Историческое значение
Гликогенфосфорилаза была первым открытым аллостерическим ферментом.[8] Он был изолирован, а его деятельность подробно охарактеризована Карл Ф. Кори, Герхард Шмидт и Герти Т. Кори.[23][24]Арда Грин и Герти Кори кристаллизовал его впервые в 1943 г. [25] и проиллюстрировал, что гликогенфосфорилаза существует либо в форме a, либо в форме b в зависимости от ее состояния фосфорилирования, а также в состояниях R или T в зависимости от присутствия AMP.[26]
Смотрите также
использованная литература
- ^ PDB: 3E3N
- ^ а б Ливанова Н.Б., Чеботарева Н.А., Еронина Т.Б., Курганов Б.И. (октябрь 2002 г.). «Пиридоксаль-5'-фосфат как каталитический и конформационный кофактор мышечной гликогенфосфорилазы B». Биохимия. Биохимия. 67 (10): 1089–98. Дои:10.1023 / А: 1020978825802. PMID 12460107. S2CID 12036788.
- ^ Palm D, Klein HW, Schinzel R, Buehner M, Helmreich EJ (февраль 1990 г.). «Роль пиридоксаль-5'-фосфата в катализе гликогенфосфорилазы». Биохимия. 29 (5): 1099–107. Дои:10.1021 / bi00457a001. PMID 2182117.
- ^ Браунер М.Ф., Флеттерик Р.Дж. (февраль 1992 г.). «Фосфорилаза: биологический преобразователь». Тенденции в биохимических науках. 17 (2): 66–71. Дои:10.1016/0968-0004(92)90504-3. PMID 1566331.
- ^ Дэвид ES, Crerar MM (январь 1986 г.). «Количественное определение мРНК мышечной гликогенфосфорилазы и количества ферментов в тканях взрослых крыс». Biochimica et Biophysica Acta (BBA) - Общие предметы. 880 (1): 78–90. Дои:10.1016/0304-4165(86)90122-4. PMID 3510670.
- ^ а б Джонсон Л.Н. (март 1992 г.). «Гликогенфосфорилаза: контроль за счет фосфорилирования и аллостерических эффекторов». Журнал FASEB. 6 (6): 2274–82. Дои:10.1096 / fasebj.6.6.1544539. PMID 1544539. S2CID 25954545.
- ^ Ньюгард С.Б., Хван П.К., Флеттерик Р.Дж. (1989). «Семейство гликогенфосфорилаз: структура и функции». Критические обзоры по биохимии и молекулярной биологии. 24 (1): 69–99. Дои:10.3109/10409238909082552. PMID 2667896.
- ^ а б Джонсон Л.Н., Барфорд Д. (февраль 1990 г.). «Гликогенфосфорилаза. Структурная основа аллостерического ответа и сравнение с другими аллостерическими белками». Журнал биологической химии. 265 (5): 2409–12. PMID 2137445.
- ^ Мейер Ф., Хейльмейер Л. М., Хашке Р. Х., Фишер Э. Х. (декабрь 1970 г.). «Контроль активности фосфорилазы в частице мышечного гликогена. I. Выделение и характеристика комплекса белок-гликоген». Журнал биологической химии. 245 (24): 6642–8. PMID 4320610.
- ^ Сомсак Л., Нагья В., Хадади З., Докса Т., Гергей П. (2003). «Аналоги глюкозы, ингибиторы гликогенфосфорилаз как потенциальные противодиабетические средства: последние разработки». Текущий фармацевтический дизайн. 9 (15): 1177–89. Дои:10.2174/1381612033454919. PMID 12769745.
- ^ Моллер Д.Е. (декабрь 2001 г.). «Новые лекарственные мишени для лечения диабета 2 типа и метаболического синдрома». Природа. 414 (6865): 821–7. Bibcode:2001Натура.414..821М. Дои:10.1038 / 414821a. PMID 11742415. S2CID 4426975.
- ^ Coats WS, Browner MF, Fletterick RJ, Newgard CB (август 1991 г.). "Сконструированная гликогенфосфорилаза печени с аллостерической активацией AMP". Журнал биологической химии. 266 (24): 16113–9. PMID 1874749.
- ^ Oikonomakos NG, Kontou M, Zographos SE, Tsitoura HS, Johnson LN, Watson KA и др. (Июль 1994). «Дизайн потенциальных противодиабетических препаратов: экспериментальное исследование ряда ингибиторов гликогенфосфорилазы аналога бета-D-глюкозы». Европейский журнал метаболизма и фармакокинетики лекарственных средств. 19 (3): 185–92. Дои:10.1007 / BF03188920. PMID 7867660. S2CID 11168623.
- ^ Hopfinger AJ, Reaka A, Venkatarangan P, Duca JS, Wang S (сентябрь 1999 г.). «Прогнозирование свободной энергии связывания лиганд-рецептор с помощью анализа 4D-QSAR: применение к набору аналогов глюкозных ингибиторов гликогенфосфорилазы». Журнал химической информации и компьютерных наук. 39 (6): 1141–1150. Дои:10.1021 / ci9900332.
- ^ Ногалес-Гадеа Дж., Аренас Дж., Андреу А.Л. (январь 2007 г.). «Молекулярная генетика болезни Макардла». Текущие отчеты по неврологии и неврологии. 7 (1): 84–92. Дои:10.1007 / s11910-007-0026-2. PMID 17217859. S2CID 39626196.
- ^ Андреу А.Л., Ногалес-Гадеа Дж., Кассандрини Д., Аренас Дж., Бруно С. (июль 2007 г.). «Болезнь Макардла: обновление молекулярной генетики». Acta Myologica. 26 (1): 53–7. ЧВК 2949323. PMID 17915571.
- ^ Грюнфельд Дж. П., Ганеваль Д., Чанард Дж., Фардо М., Дрейфус Дж. К. (июнь 1972 г.). «Острая почечная недостаточность при болезни Макардла. Сообщение о двух случаях». Медицинский журнал Новой Англии. 286 (23): 1237–41. Дои:10.1056 / NEJM197206082862304. PMID 4502558.
- ^ Бурвинкель Б., Баккер Х.Д., Гершковиц Э., Мозес С.В., Шин Ю.С., Килиманн М.В. (апрель 1998 г.). «Мутации в гене гликогенфосфорилазы печени (PYGL), лежащие в основе гликогеноза VI типа». Американский журнал генетики человека. 62 (4): 785–91. Дои:10.1086/301790. ЧВК 1377030. PMID 9529348.
- ^ Чанг С., Розенберг М.Дж., Мортон Х., Франкомано, Калифорния, Бизекер Л.Г. (май 1998 г.). «Идентификация мутации гликогенфосфорилазы печени при болезни накопления гликогена VI типа». Молекулярная генетика человека. 7 (5): 865–70. Дои:10,1093 / чмг / 7,5,865. PMID 9536091.
- ^ Тан Н.Л., Хуэй Дж., Янг Э., Уортингтон В., То К.Ф., Чунг К.Л. и др. (Июнь 2003 г.). «Новая мутация (G233D) в гене гликогенфосфорилазы у пациента с болезнью накопления гликогена в печени и остаточной ферментативной активностью». Молекулярная генетика и метаболизм. 79 (2): 142–5. Дои:10.1016 / S1096-7192 (03) 00068-4. PMID 12809646.
- ^ Шимада С., Мацузаки Х., Маруцука Т., Сиомори К., Огава М. (июль 2001 г.). «Желудочные и кишечные фенотипы карциномы желудка со ссылкой на экспрессию гликогенфосфорилазы головного (фетального) типа». Журнал гастроэнтерологии. 36 (7): 457–64. Дои:10.1007 / s005350170068. PMID 11480789. S2CID 25602637.
- ^ Alemany S, Pelech S, Brierley CH, Cohen P (апрель 1986). «Протеиновые фосфатазы, участвующие в клеточной регуляции. Доказательства того, что дефосфорилирование гликогенфосфорилазы и гликогенсинтазы в гликогеновой и микросомальной фракциях печени крысы катализируется одним и тем же ферментом: протеинфосфатазой-1». Европейский журнал биохимии. 156 (1): 101–10. Дои:10.1111 / j.1432-1033.1986.tb09554.x. PMID 3007140.
- ^ Кори CF, Шмидт G, Кори GT (май 1939 г.). «Синтез полисахарида из глюкозо-1-фосфата в мышечном экстракте». Наука. 89 (2316): 464–5. Bibcode:1939Sci .... 89..464C. Дои:10.1126 / наука.89.2316.464. PMID 17731092.
- ^ Cori GT, Cori CF (июль 1940 г.). «Кинетика ферментативного синтеза гликогена из глюкозо-1-фосфата». Журнал биологической химии. 135: 733–756.
- ^ Грин AA, Cori GT (7 июля 1943 г.). «Кристаллическая мышечная фосфорилаза I. Получение, свойства и молекулярный вес». Журнал биологической химии. 151: 21–29.
- ^ Cori GT, Green AA (июль 1943 г.). «Кристаллическая мышечная фосфорилаза II простетической группы». Журнал биологической химии. 151 (1): 21–29.
дальнейшее чтение
- Voet JG, Voet D (1995). «Глава 17: Метаболизм гликогена». Биохимия (2-е изд.). Нью-Йорк: J. Wiley & Sons. ISBN 978-0-471-58651-7.
- Voet JG, Voet D (2004). «Глава 18: Метаболизм гликогена». Биохимия (3-е изд.). Нью-Йорк: J. Wiley & Sons. ISBN 978-0-471-19350-0.
- Гудселл Д.С. (01.12.2001). «Гликогенфосфорилаза». Молекула месяца. Банк данных белков RCSB. Получено 2009-01-10.
- Диван Дж. Дж. «Метаболизм гликогена». Молекулярная биохимия I. Политехнический институт Ренсселера. Архивировано из оригинал на 2009-01-25. Получено 2009-01-10.
внешние ссылки
- GeneReviews / NCBI / NIH / UW запись о болезни накопления гликогена типа VI - ее болезнь
- Гликоген + фосфорилаза в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- Обзор всей структурной информации, доступной в PDB для UniProt: P11217 (Человеческая мышечная гликогенфосфорилаза) на PDBe-KB.
- Обзор всей структурной информации, доступной в PDB для UniProt: P06737 (Гликогенфосфорилаза печени человека) на PDBe-KB.
- Обзор всей структурной информации, доступной в PDB для UniProt: P11216 (Гликогенфосфорилаза головного мозга человека) на PDBe-KB.