Изоцитратдегидрогеназа - Isocitrate dehydrogenase
| Изоцитратдегидрогеназа | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Кристаллографическая структура Кишечная палочка изоцитратдегидрогеназа.[1] Есть три активных сайта. Три изоцитрата, один изоцитрат в сайте связывания NADP+. | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 1.1.1.42 | ||||||||
| Количество CAS | 9028-48-2 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
| изоцитратдегидрогеназа (НАД+) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Номер ЕС | 1.1.1.41 | ||||||||
| Количество CAS | 9001-58-5 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
| Мономерная изоцитратдегидрогеназа | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 кристаллическая структура мономерной изоцитратдегидрогеназы в комплексе с изоцитратом и mn | |||||||||
| Идентификаторы | |||||||||
| Символ | IDH | ||||||||
| Pfam | PF03971 | ||||||||
| Pfam клан | CL0270 | ||||||||
| ИнтерПро | IPR004436 | ||||||||
| SCOP2 | 1фг / Объем / СУПФАМ | ||||||||
| |||||||||
Изоцитратдегидрогеназа (IDH) (EC 1.1.1.42 ) и (EC 1.1.1.41 ) является фермент который катализирует окислительное декарбоксилирование изоцитрат, производя альфа-кетоглутарат (α-кетоглутарат) и CO2. Это двухэтапный процесс, который включает окисление изоцитрат (вторичный алкоголь ) к оксалосукцинат (а кетон ) с последующим декарбоксилированием бета-карбоксильной группы до кетона с образованием альфа-кетоглутарата. У человека IDH существует в трех изоформах: IDH3 катализирует третью стадию цикл лимонной кислоты при преобразовании НАД+ к НАДН в митохондрии. Изоформы IDH1 и IDH2 катализировать ту же реакцию вне контекста цикла лимонной кислоты и использовать НАДФ+ в качестве кофактора вместо NAD+. Они локализуются как в цитозоле, так и в митохондрия и пероксисома.[2]
Изоферменты
Ниже приводится список изоферментов изоцитратдегидрогеназы человека:
НАДФ+ зависимый
Каждый НАДП+-зависимый изофермент действует как гомодимер:
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
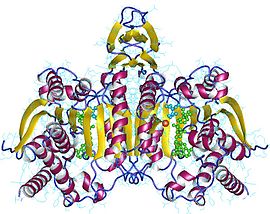
НАД+ зависимый
Изоцитратдегидрогеназа 3 представляет собой гетеротетрамер, который состоит из двух альфа-субъединиц, одной бета-субъединицы и одной гамма-субъединицы:
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Структура

NAD-IDH состоит из 3 субъединиц, регулируется аллостерически и требует интегрированного Mg2+ или Mn2+ ион. Ближайшим гомологом с известной структурой является Кишечная палочка НАДФ-зависимая IDH, которая имеет только 2 субъединицы и 13% идентичности и 29% сходства на основе аминокислотных последовательностей, что делает ее непохожей на IDH человека и не подходящей для близкого сравнения. Все известные НАДФ-ИДГ являются гомодимерами.
Большинство изоцитратдегидрогеназ являются димерами, а именно гомодимерами (две идентичные мономерные субъединицы, образующие одно димерное звено). При сравнении C. glutamicum и Кишечная палочка,[4] мономер и димер, соответственно, оба фермента «эффективно катализируют идентичные реакции». Тем не мение, C. glutamicum был зарегистрирован как имеющий в десять раз большую активность, чем Кишечная палочка и в семь раз больше аффинитивных / специфичных для НАДФ. C. glutamicum одобрил НАДФ+ над NAD+. Что касается стабильности в зависимости от температуры, оба фермента имели одинаковую Tm или температуру плавления примерно от 55 ° C до 60 ° C. Однако мономер C. glutamicum показал более стабильную стабильность при более высоких температурах, чего и следовало ожидать. Димер Кишечная палочка показал стабильность при более высокой температуре, чем обычно, из-за взаимодействий между двумя мономерными субъединицами.
Структура Микобактерии туберкулеза (Mtb) ICDH-1, связанный с NADPH и Mn (2+), был определен с помощью рентгеновской кристаллографии. Это гомодимер, в котором каждая субъединица имеет складку Россмана и общую верхнюю область взаимосвязанных β-листов. Mtb ICDH-1 наиболее структурно сходен с мутантным R132H ICDH человека, обнаруженным в глиобластомах. Подобно человеческому R132H ICDH, Mtb ICDH-1 также катализирует образование α-гидроксиглутарата.[5]
Регулирование
Стадия IDH цикла лимонной кислоты часто (но не всегда) является необратимой реакцией в цикле лимонной кислоты из-за ее большого отрицательного изменения свободной энергии. Следовательно, его необходимо тщательно регулировать, чтобы избежать истощения изоцитрата (и, следовательно, накопления альфа-кетоглутарата). Реакция стимулируется простыми механизмами доступности субстрата (изоцитрат, НАД+ или же НАДФ+, Mg2+ / Mn2+ ), ингибирование продукта (по НАДН (или же НАДФН вне цикла лимонной кислоты) и альфа-кетоглутарата), а также ингибирование конкурентной обратной связи (путем АТФ ).[6]
Каталитические механизмы
Изоцитратдегидрогеназа катализирует химические реакции:
- Изоцитрат + НАД+ 2-оксоглутарат + CO2 + НАДН + Н+
- Изоцитрат + НАДФ+ 2-оксоглутарат + CO2 + НАДФН + Н+[7][8][9]

Общая свободная энергия для этой реакции составляет -8,4 кДж / моль.[10]

Шаги
В рамках цикл лимонной кислоты, изоцитрат, образующийся при изомеризации цитрата, подвергается как окисление и декарбоксилирование. С помощью фермента изоцитратдегидрогеназа (IDH) изоцитрат удерживается в своем активный сайт окружая аргинин, тирозин, аспарагин, серин, треонин, и аспарагиновая кислота аминокислоты. Первое поле показывает общую реакцию изоцитратдегидрогеназы. Реагентами, необходимыми для работы этого ферментного механизма, являются изоцитрат, НАД+ /НАДФ+, а Mn2+ или Mg2+. Продукты реакции: альфа-кетоглутарат, углекислый газ, и НАДН + H+/НАДФН + H+.[8] Молекулы воды используются для депротонирования кислорода (O3) изоцитрата.
Второе поле - это этап 1, который представляет собой окисление альфа-C (C # 2).[7][8] Окисление - это первая стадия, через которую проходит изоцитрат. В этом процессе[7] то алкоголь группа от альфа-углерод (C # 2) депротонируется, и электроны текут к альфа-C, образуя кетон группа и удаление гидрид выключен C # 2 с помощью NAD+/ НАДФ+ как электрон, принимающий кофактор. Окисление альфа-C позволяет создать положение, в котором электроны (на следующем этапе) будут выходить из карбоксил группа и толкает электроны (создавая кислород с двойной связью) обратно на кислород или захватывая ближайший протон у ближайшего Лизин аминокислота.
Третье поле - это шаг 2, который представляет собой декарбоксилирование оксалосукцинат. На этом этапе[7][8] кислород карбоксильной группы депротонируется соседним Тирозин аминокислоты, и эти электроны стекают к углероду 2. Двуокись углерода покидает бета-углерод изоцитрата в качестве уходящей группы с электронами, протекающими к кислороду кетона от альфа-C, создавая отрицательный заряд на кислороде альфа-C и образуя ненасыщенную двойную связь альфа-бета между атомами углерода 2 и 3. одинокая пара на альфа-С кислород улавливает протон из ближайшей аминокислоты лизина.
Четвертое поле - это этап 3, который представляет собой насыщение альфа-бета-ненасыщенной двойной связи между атомами углерода 2 и 3. На этом этапе реакции[7][8] Лизин депротонирует кислород с альфа-углерода, и неподеленная пара электронов на кислороде альфа-углерода опускается вниз, реформируя двойную связь кетона и отталкивая неподеленную пару (образуя двойную связь между альфа- и бета-углеродом), собирая протон из соседней аминокислоты тирозин.[12] Эта реакция приводит к образованию альфа-кетоглутарата, НАДН + Н.+/ НАДФН + Н+, и CO2.
Подробный механизм
Два аспартат аминокислотные остатки (внизу слева) взаимодействуют с двумя соседними молекулами воды (w6 и w8) в Mn2+ изоцитрат свиного IDH комплекса для депротонирования спирта с альфа-углеродного атома. Окисление альфа-C также происходит на этой картинке, где НАД+ принимает гидрид с образованием оксалосукцината. Вместе с зр3 к зр2 стереохимический изменение вокруг альфа-С, есть кетонная группа, которая образуется из спиртовой группы. Образование этой двойной связи кетона позволяет иметь место резонансу, когда электроны, идущие от уходящей карбоксилатной группы, движутся к кетону.
Декарбоксилирование оксалосукцината (внизу в центре) является ключевым этапом в образовании альфа-кетоглутарата. В этой реакции неподеленная пара на соседнем гидроксиле тирозина отрывает протон от карбоксильной группы.[12] Эта карбоксильная группа также называется бета-субъединицей в молекуле изоцитрата. Депротонирование карбоксильной группы заставляет неподеленную пару электронов двигаться вниз, образуя диоксид углерода и отделяясь от оксалосукцината. Электроны продолжают двигаться к альфа-углероду, выталкивая электроны с двойной связью (образуя кетон), чтобы оторвать протон от соседнего остатка лизина. Альфа-бета-ненасыщенная двойная связь образуется между углеродом 2 и 3. Как вы можете видеть на картинке, зеленый ион представляет собой либо Mg2+ или Mn2+, который является кофактором, необходимым для протекания этой реакции. Ион металла образует небольшой комплекс за счет ионного взаимодействия с атомами кислорода на четвертом и пятом атомах углерода (также известный как гамма-субъединица изоцитрата).
После отделения диоксида углерода от оксалосукцината на стадии декарбоксилирования (внизу справа) энол будет таутомеризоваться в кето из. Образование двойной связи кетона начинается с депротонирование кислорода из альфа-углерода (C # 2) тем же лизином, который протонировал кислород в первую очередь.[12] Неподеленная пара электронов движется вниз, отталкивая неподеленные пары, образующие двойную связь. Эта неподеленная пара электронов отрывает протон от тирозина, который депротонировал карбоксильную группу на стадии декарбоксилирования. Причина, по которой мы можем сказать, что остатки Lys и Tyr будут такими же, как и на предыдущем этапе, заключается в том, что они помогают удерживать молекулу изоцитрата в активном центре фермента. Эти два остатка будут способны образовывать водородные связи взад и вперед, пока они находятся достаточно близко к субстрат.[4]
 Стадия оксидоредуктазы, где НАД+ используется для принятия гидрида.[12] |  Декарбоксилирование оксалосукцината.[12] |  Насыщение альфа-бета-ненасыщенной двойной связи.[12] |
Фермент изоцитратдегидрогеназа, как указано выше, производит альфа-кетоглутарат, диоксид углерода и NADH + H.+/ НАДФН + Н+. За время реакции произошло три изменения. Окисление углерода 2, декарбоксилирование (потеря диоксида углерода) углерода 3 и образование кетонной группы со стереохимическим изменением от sp3 к sp2.[12]
 |  Митохондриальный НАДФ свиньи+-зависимая изоцитратдегидрогеназа в комплексе с Mn2+ и изоцитрат. Вид поверхности активный сайт карман, в котором изоцитрат ограничен полярными аминокислотами.[3] |  Митохондриальный НАДФ свиньи+-зависимая изоцитратдегидрогеназа в комплексе с Mn2+ и изоцитрат.[3] |  Свиной ферментный комплекс; Изоцитрат активного сайта и соседний A.A.[3] |
Активный сайт

Структура фермента изоцитратдегидрогеназа (IDH) в кишечная палочка была первой структурой, которая была выяснена и понята.[12] С тех пор кишечная палочка Структура IDH использовалась большинством исследователей для сравнения с другими ферментами изоцитратдегидрогеназы. Об этом бактериальном ферменте известно много подробностей, и было обнаружено, что большинство изоцитратдегидрогеназ схожи по структуре и, следовательно, по функциям. Это сходство структуры и функции дает основание полагать, что структуры консервативны так же, как и аминокислоты.[9] Следовательно, активные центры среди большинства прокариотических ферментов изоцитратдегидрогеназы также должны быть сохранены, что наблюдается во многих исследованиях, проведенных на прокариотических ферментах. С другой стороны, эукариотические ферменты изоцитратдегидрогеназы еще не полностью открыты. Каждый димер IDH имеет два активных центра.[12] Каждый активный сайт связывает NAD+/ НАДФ+ молекула и ион двухвалентного металла (Mg2+, Mn2+). В общем, каждый активный сайт имеет консервативную последовательность аминокислот для каждого специфического сайта связывания. В Десульфоталея психрофила (ДпIDH)[12] и свиной (ПКIDH)[3] с активным сайтом связаны три субстрата.
- Изоцитрат связывается в активном сайте с консервативной последовательностью примерно из восьми аминокислот посредством водородных связей. Эти кислоты включают (могут различаться по остатку, но с аналогичными свойствами) тирозин, серин, аспарагин, аргинин, аргинин, аргинин, тирозин и лизин. Их положение на позвоночнике различается, но все они находятся в близком диапазоне (т.е. Arg131 DpIDH и Arg133 PcIDH, Tyr138 DpIDH и Tyr140 PcIDH).[12]
- Ион металла (Mg2+, Mn2+) связывается с тремя консервативными аминокислотами через водородные связи. Эти аминокислоты включают три остатка аспартата.[12]
- НАД+ и НАДФ+ связываются в активном центре в четырех областях с аналогичными свойствами среди ферментов IDH. Эти регионы различаются, но составляют около [250–260], [280–290], [300–330] и [365–380]. Снова регионы различаются, но близость регионов сохраняется.[12]
Клиническое значение

Специфические мутации в гене изоцитратдегидрогеназы IDH1 были обнаружены в нескольких опухолях головного мозга, включая астроцитома, олигодендроглиома и мультиформная глиобластома, с мутациями, обнаруженными почти во всех случаях вторичных глиобластом, которые развиваются из глиом более низкой степени злокачественности, но редко в первичных глиобластомах высокой степени злокачественности. мультиформная глиобластома.[14] Пациенты, у которых опухоль имела мутацию IDH1, выживали дольше.[15][16] Более того, мутации IDH2 и IDH1 были обнаружены почти в 20% цитогенетически нормальных острый миелоидный лейкоз (AML).[17][18] Известно, что эти мутации производят (D) -2-гидроксиглутарат из альфа-кетоглутарата.[19] (D) -2-гидроксиглутарат накапливается до очень высоких концентраций, которые подавляют функцию ферментов, которые зависят от альфа-кетоглутарата.[20] Это приводит к гиперметилированному состоянию ДНК и гистонов, что приводит к различной экспрессии генов, которые могут активировать онкогены и инактивировать гены-супрессоры опухолей. В конечном итоге это может привести к типам рака, описанным выше.[21] Соматические мозаичные мутации этого гена также были связаны с Болезнь Оллье и Синдром Маффуччи.[22] Однако недавние исследования также показали, что (D) -2-гидроксиглутарат может превращаться обратно в альфа-кетоглутарат ферментативно или неферментативно.[23][24] Необходимы дальнейшие исследования, чтобы полностью понять роль мутации IDH1 (и (D) -2-гидроксиглутарата) при раке.
Смотрите также
Рекомендации
- ^ PDB: 1CW7; Cherbavaz DB, Lee ME, Stroud RM, Koshland DE (январь 2000 г.). «Молекулы воды с активным центром, обнаруженные в разрешающей структуре 2.1 A сайт-направленного мутанта изоцитратдегидрогеназы». J. Mol. Биол. 295 (3): 377–85. Дои:10.1006 / jmbi.1999.3195. PMID 10623532.
- ^ Corpas FJ, Barroso JB, Sandalio LM, Palma JM, Lupiáñez JA, del Río LA (1999). "Пероксисомальная НАДФ-зависимая изоцитратдегидрогеназа. Характеристика и регулирование активности при естественном старении". Физиология растений. 121 (3): 921–928. Дои:10.1104 / стр.121.3.921. ЧВК 59455. PMID 10557241.
- ^ а б c d е ж PDB: 1LWD; Чеккарелли С., Нил Б. (2002). «Кристаллическая структура митохондриального НАДФ свиньи.+-зависимая изоцитратдегидрогеназа в комплексе с Mn2+ и изоцитрат ». Журнал биологической химии. 277 (45): 43454–43462. Дои:10.1074 / jbc.M207306200. PMID 12207025.
- ^ а б Чен Р., Ян Х (ноябрь 2000 г.). "Высокоспецифичная мономерная изоцитратдегидрогеназа из Коринебактерии глутамикум". Arch. Biochem. Биофизы. 383 (2): 238–45. Дои:10.1006 / abbi.2000.2082. PMID 11185559.
- ^ Quartararo CE, Hazra S, Hadi T, Blanchard JS (2013). «Структурный, кинетический и химический механизм изоцитратдегидрогеназы-1 из Mycobacterium tuberculosis». Биохимия. 52 (10): 1765–75. Дои:10.1021 / bi400037w. ЧВК 3706558. PMID 23409873.
- ^ Мэйтинг, Инес; Шмидт, Георг; Sahm, Hermann; Стахманн, К.-Петер (2000). «Роль пероксисомальной НАДФ-специфической изоцитратдегидрогеназы в метаболизме избыточного продуцента рибофлавина Ashbya gossypii». Журнал молекулярного катализа B: ферментативный. 10 (1–3): 335–343. Дои:10.1016 / S1381-1177 (00) 00135-1.
- ^ а б c d е Тадг П. Бегли; Макмерри, Джон (2005). Органическая химия биологических путей. Робертс и Ко. Издатели. С. 189–190. ISBN 0-9747077-1-6.
- ^ а б c d е Кокс, Майкл; Нельсон, Дэвид Р .; Ленингер, Альберт Л (2005). Принципы биохимии Ленингера. Сан-Франциско: W.H. Фримен. стр.609–611. ISBN 0-7167-4339-6.
- ^ а б Ясутакэ Й, Ватанабэ С., Яо М., Такада Й, Фукунага Н., Танака И. (2003). «Кристаллическая структура мономерной изоцитратдегидрогеназы в присутствии НАДФ.+". Журнал биологической химии. 278 (38): 36897–36904. Дои:10.1074 / jbc.M304091200. PMID 12855708.
- ^ Гаррет, Реджинальд; Гришэм, Чарльз М. (2012). Биохимия. Cengage Learning. п. 621. ISBN 978-1133106296.
- ^ Аошима М., Игараси Ю. (март 2008 г.). «Недекарбоксилирующие и декарбоксилирующие изоцитратдегидрогеназы: оксалосукцинатредуктаза как предковая форма изоцитратдегидрогеназы». Журнал бактериологии. 190 (6): 2050–5. Дои:10.1128 / JB.01799-07. ЧВК 2258884. PMID 18203822.
- ^ а б c d е ж грамм час я j k л м Fedøy AE, Yang N, Martinez A, Leiros HK, Steen IH (сентябрь 2007 г.). «Структурные и функциональные свойства изоцитратдегидрогеназы психрофильной бактерии. Десульфоталея психрофила выявить активный к холоду фермент с необычно высокой термостабильностью ». J. Mol. Биол. 372 (1): 130–49. Дои:10.1016 / j.jmb.2007.06.040. PMID 17632124.
- ^ Каппер Д., Центграф Х., Бальсс Дж., Хартманн С., фон Даймлинг А. (ноябрь 2009 г.). «Моноклональные антитела, специфичные к мутации IDH1 R132H». Acta Neuropathol. 118 (5): 599–601. Дои:10.1007 / s00401-009-0595-z. PMID 19798509. S2CID 36093146.
- ^ Бликер, ИП; Molenaar, RJ; Леенстра, S (май 2012 г.). «Последние достижения в молекулярном понимании глиобластомы». Журнал нейроонкологии. 108 (1): 11–27. Дои:10.1007 / s11060-011-0793-0. ЧВК 3337398. PMID 22270850.
- ^ Хартманн С., Хентшель Б., Вик В. и др. (Декабрь 2010 г.). «Пациенты с анапластическими астроцитомами IDH1 дикого типа имеют худший прогноз, чем глиобластомы с мутацией IDH1, а статус мутации IDH1 объясняет неблагоприятный прогностический эффект старшего возраста: значение для классификации глиом» (PDF). Acta Neuropathol. 120 (6): 707–18. Дои:10.1007 / s00401-010-0781-z. PMID 21088844. S2CID 7323032.
- ^ Molenaar, RJ; Verbaan, D; Ламба, S; Занон, К; Jeuken, JW; Ботинки-Шпренгер, SH; Wesseling, P; Hulsebos, TJ; Troost, D; ван Тилборг, AA; Leenstra, S; Вандертоп, WP; Барделли, А; ван Ноорден, CJ; Бликер, ИП (сентябрь 2014 г.). «Комбинация мутаций IDH1 и статуса метилирования MGMT предсказывает выживаемость при глиобластоме лучше, чем только IDH1 или MGMT». Нейроонкология. 16 (9): 1263–73. Дои:10.1093 / neuonc / nou005. ЧВК 4136888. PMID 24510240.
- ^ Уорд П.С., Патель Дж., Мудрый Д.Р. и др. (Март 2010 г.). «Общей чертой связанных с лейкемией мутаций IDH1 и IDH2 является активность неоморфного фермента, превращающего альфа-кетоглутарат в 2-гидроксиглутарат». Раковая клетка. 17 (3): 225–34. Дои:10.1016 / j.ccr.2010.01.020. ЧВК 2849316. PMID 20171147.
- ^ Ван И, Сяо М, Чен Х, Чен Л, Сюй И, Ур Л, Ван П, Ян Х, Ма С, Лин Х, Цзяо Б., Рен Р, Йе Д, Гуань К.Л., Сюн Й. (февраль 2015 г.). «WT1 рекрутирует TET2 для регулирования экспрессии своего целевого гена и подавления пролиферации лейкозных клеток». Молекулярная клетка. 57 (4): 662–73. Дои:10.1016 / j.molcel.2014.12.023. ЧВК 4336627. PMID 25601757.
- ^ Dang L, White DW, Gross S, Bennett BD, Bittinger MA, Driggers EM, Fantin VR, Jang HG, Jin S, Keenan MC, Marks KM, Prins RM, Ward PS, Yen KE, Liau LM, Rabinowitz JD, Cantley LC , Thompson CB, Vander Heiden MG, Su SM (июнь 2010 г.). «Связанные с раком мутации IDH1 продуцируют 2-гидроксиглутарат». Природа. 465 (7300): 966. Bibcode:2010Натура.465..966D. Дои:10.1038 / природа09132. ЧВК 3766976. PMID 20559394.CS1 maint: использует параметр авторов (связь)
- ^ Чоудхури Р., Йео К.К., Тиан Ю.М., Хиллрингхаус Л., Багг Е.А., Роуз Н.Р., Люнг И.К., Ли XS, Вун ЕС, Ян М., Макдонау М.А., Кинг О.Н., Клифтон И.Дж., Клозе Р.Дж., Кларидж ТДВ, Рэтклифф П.Дж., Шофилд С.Дж. , Кавамура А. (май 2011 г.). «Онкометаболит 2-гидроксиглутарат ингибирует гистоновые лизиндеметилазы». Отчеты EMBO. 12 (5): 463–9. Дои:10.1038 / embor.2011.43. ЧВК 3090014. PMID 21460794.CS1 maint: использует параметр авторов (связь)
- ^ Molenaar, RJ; Радивоевич, Т; Maciejewski, JP; van Noorden, CJ; Бликер, ИП (28 мая 2014 г.). «Драйвер и пассажир эффекты мутаций изоцитратдегидрогеназы 1 и 2 в онкогенезе и продлении жизни». Biochimica et Biophysica Acta (BBA) - Обзоры на рак. 1846 (2): 326–341. Дои:10.1016 / j.bbcan.2014.05.004. PMID 24880135.
- ^ Amary MF, Damato S, Halai D, Eskandarpour M, Berisha F, Bonar F, McCarthy S, Fantin VR, Straley KS, Lobo S, Aston W, Green CL, Gale RE, Tirabosco R, Futreal A, Campbell P, Presneau N , Фланаган AM (2011). «Болезнь Олли и синдром Маффуччи вызваны соматическими мозаичными мутациями IDH1 и IDH2». Nat. Genet. 43 (12): 1262–5. Дои:10,1038 / нг.994. PMID 22057236. S2CID 5592593.
- ^ Tarhonskaya H, Rydzik AM, Leung IKH, Loik ND, Chan MC, Kawamura A, McCullagh JS, Claridge TDW, Flashman E, Schofield CJ (март 2014 г.). «Неферментативная химия делает возможной активацию 2-оксоглутарат оксигеназ, опосредованную 2-гидроксиглутаратом». Nature Communications. 5: 3423. Bibcode:2014 НатКо ... 5,3423 т. Дои:10.1038 / ncomms4423. ЧВК 3959194. PMID 24594748.CS1 maint: использует параметр авторов (связь)
- ^ Койвунен П., Ли С., Дункан К. Г., Лопес Дж., Лу Дж., Рамкиссун С., Лосман Дж. А., Йоэнсуу П., Бергманн Ю., Гросс С., Травинс Дж., Вайс С., Лупер Р., Лигон К. Л., Верхаак Р. Г., Ян Х., Келин В. Г. Jr (февраль 2012 г.). «Трансформация (R) -энантиомером 2-гидроксиглутарата, связанная с активацией EGLN» (PDF). Природа. 483 (7390): 484–8. Bibcode:2012Натура.483..484K. Дои:10.1038 / природа10898. ЧВК 3656605. PMID 22343896.CS1 maint: использует параметр авторов (связь)
внешняя ссылка
- Изоцитратдегидрогеназа: молекула месяца RCSB PDB
- Обзор всей структурной информации, доступной в PDB за UniProt: O75874 (Изоцитратдегидрогеназа [НАДФ] цитоплазма) в PDBe-KB.
- Обзор всей структурной информации, доступной в PDB за UniProt: P48735 (Изоцитратдегидрогеназа [НАДФ], митохондриальная) в PDBe-KB.
- Обзор всей структурной информации, доступной в PDB за UniProt: P50213 (Субъединица альфа изоцитратдегидрогеназы [НАД], митохондриальная) на PDBe-KB.
- Обзор всей структурной информации, доступной в PDB за UniProt: O43837 (Бета-субъединица изоцитратдегидрогеназы [НАД], митохондриальная) на PDBe-KB.
- Обзор всей структурной информации, доступной в PDB за UniProt: P51553 (Гамма-субъединица изоцитратдегидрогеназы [НАД], митохондриальная) на PDBe-KB.