Долгосрочное потенцирование - Long-term potentiation

В нейробиология, долгосрочное потенцирование (LTP) - стойкое усиление синапсы на основе недавних моделей активности. Это паттерны синаптической активности, которые производят длительное усиление передачи сигнала между двумя нейроны.[2] Противоположностью LTP является длительная депрессия, что приводит к длительному снижению синаптической силы.
Это одно из нескольких явлений, лежащих в основе синаптическая пластичность, способность химические синапсы изменить свою силу. Считается, что воспоминания кодируются путем модификации синаптическая сила,[3] LTP широко считается одним из основных клеточных механизмов, лежащих в основе обучение и объем памяти.[2][3]
LTP был обнаружен у кролика гиппокамп от Терье Лёмо в 1966 году и с тех пор остается популярным объектом исследований. Многие современные исследования LTP стремятся лучше понять его основную биологию, в то время как другие стремятся установить причинную связь между LTP и поведенческим обучением. Третьи пытаются разработать фармакологические или иные методы усиления ДП для улучшения обучения и памяти. LTP также является предметом клинические исследования, например, в областях Болезнь Альцгеймера и лекарство от наркозависимости.
История
Ранние теории обучения

В конце XIX века ученые в целом признали, что количество нейронов в мозге взрослого человека (примерно 100 миллиардов[4]) не увеличивалась значительно с возрастом, что дает нейробиологам веские основания полагать, что воспоминания, как правило, не были результатом производства новых нейронов.[5] С осознанием этого пришла необходимость объяснить, как воспоминания могут формироваться в отсутствие новых нейронов.
В испанский нейроанатом Сантьяго Рамон-и-Кахаль был одним из первых, кто предложил механизм обучения, не требующий образования новых нейронов. В его 1894 г. Кроунская лекция он предположил, что вместо этого воспоминания могут быть сформированы путем усиления связей между существующими нейронами для повышения эффективности их коммуникации.[5] Хеббийская теория, представлен Дональд Хебб в 1949 году повторил идеи Рамона-и-Кахала, предположив, что клетки могут создавать новые связи или претерпевать метаболические и синаптические изменения, которые повышают их способность общаться и создавать нейронную сеть опыта.[6]:
Предположим, что постоянство или повторение реверберационной активности (или «следа») имеет тенденцию вызывать длительные клеточные изменения, которые добавляют к ее стабильности ... Когда аксон клетки А находится достаточно близко, чтобы возбуждать клетку В и многократно или постоянно участвует в его возбуждении, в одной или обеих клетках происходят некоторые процессы роста или метаболические изменения, так что эффективность A как одной из клеток, запускающих B, увеличивается.[7]
Эрик Кандел (1964) и его сотрудники были одними из первых исследователей, открывших долгосрочное потенцирование во время работы с морскими слизнями Aplysia. Они попытались применить поведенческое кондиционирование к различным клеткам нейронной сети слизня. Их результаты показали изменения синаптической силы, и исследователи предположили, что это может быть связано с базовой формой обучения, происходящей внутри слизняка.[8][9]
Хотя эти теории формирования памяти в настоящее время хорошо известны, они были дальновидными для своего времени: нейробиологи и психологи конца 19 - начала 20 веков не были оснащены нейрофизиологический методы, необходимые для выяснения биологических основ обучения у животных. Эти навыки появятся только во второй половине 20 века, примерно в то же время, что и открытие долгосрочного потенцирования.
Открытие
LTP впервые наблюдал Терье Лёмо в 1966 г. в Осло, Норвегия, лаборатория Пер Андерсен.[10][11] Там Лёмо провел серию нейрофизиологический эксперименты на под наркозом кроликов, чтобы изучить роль гиппокампа в краткосрочная память.
Эксперименты Лёмо были сосредоточены на связях или синапсах из перфорантный путь к зубчатые извилины. Эти эксперименты проводились путем стимуляции пресинаптических волокон перфорантного пути и регистрации ответов от набора постсинаптических клеток зубчатой извилины. Как и ожидалось, одиночный импульс электростимуляции волокон перфорантного пути вызвал возбуждающие постсинаптические потенциалы (ВПСП) в клетках зубчатой извилины. Неожиданно Лемо заметил, что реакция постсинаптических клеток на эти одноимпульсные стимулы может быть усилена в течение длительного периода времени, если он сначала доставит высокочастотная последовательность раздражителей к пресинаптическим волокнам. Когда применялась такая последовательность стимулов, последующие одноимпульсные стимулы вызывали более сильные и продолжительные ВПСП в популяции постсинаптических клеток. Этот феномен, при котором высокочастотный стимул может вызвать долгосрочное усиление реакции постсинаптических клеток на последующие одноимпульсные стимулы, первоначально был назван «длительной потенциацией».[12][13]
Тимоти Блисс, который присоединился к лаборатории Андерсена в 1968 году,[10] сотрудничал с Лемо, и в 1973 году они опубликовали первую характеристику длительного потенцирования в кролик гиппокамп.[12] Блисс и Тони Гарднер-Медвин опубликовали аналогичный отчет о длительном потенцировании у бодрствующего животного, который появился в том же выпуске, что и отчет Блисс и Лёмо.[13] В 1975 году Дуглас и Годдард предложили «долгосрочное потенцирование» в качестве нового названия для феномена долговременного потенцирования.[14][15] Андерсен предположил, что авторы выбрали «долгосрочное потенцирование», возможно, из-за легко произносимого акронима «LTP».[16]
Модели и теория


Физический и биологический механизм LTP до сих пор не изучен, но были разработаны некоторые успешные модели.[1] Исследования дендритные шипы, выступающие структуры на дендритах, которые физически растут и втягиваются в течение минут или часов, предположили связь между электрическое сопротивление позвоночника и эффективной прочности синапсов из-за их взаимосвязи с переходными процессами внутриклеточного кальция. Математические модели, такие как BCM Теория, который зависит также от внутриклеточного кальция по отношению к Рецептор NMDA ворота напряжения, были разработаны с 1980-х годов и модифицируют традиционные априори Hebbian обучение модель с биологическим и экспериментальным обоснованием. Третьи предложили перестроить или синхронизировать отношения между регуляцией рецепторов, LTP и синаптической силой.[17]
Типы
С момента своего первоначального открытия в гиппокампе кроликов, LTP наблюдали во множестве других нервных структур, включая кора головного мозга,[18] мозжечок,[19] миндалина,[20] и многие другие. Роберт Маленка, известный исследователь LTP, предположил, что LTP может даже встречаться во всех возбуждающих синапсах в мозге млекопитающих.[21]
В разных областях мозга обнаруживаются разные формы ДП. Конкретный тип LTP, проявляемый между нейронами, зависит от ряда факторов. Одним из таких факторов является возраст организма, когда наблюдается ДП. Напр., Молекулярные механизмы LTP в незрелом гиппокампе отличаются от тех механизмов, которые лежат в основе LTP гиппокампа взрослых.[22] Сигнальные пути, используемые конкретной клеткой, также вносят вклад в конкретный тип присутствующего LTP. Например, некоторые типы ДП гиппокампа зависят от Рецептор NMDA, другие могут зависеть от метаботропный рецептор глутамата (mGluR), тогда как другие полностью зависят от другой молекулы.[21] Разнообразие сигнальных путей, которые вносят вклад в LTP, и широкое распространение этих различных путей в головном мозге являются причинами того, что тип LTP, проявляемый между нейронами, лишь частично зависит от анатомического местоположения, в котором наблюдается LTP. Например, LTP в Обеспечение Schaffer путь гиппокампа зависит от рецептора NMDA - это было доказано применением AP5, антагонист рецептора NMDA, который предотвращает LTP на этом пути.[23] И наоборот, LTP в моховое волокно Путь не зависит от рецептора NMDA, хотя оба пути находятся в гиппокампе.[24]
Пре- и постсинаптическая активность, необходимая для индукции LTP, - это другие критерии, по которым классифицируется LTP. В целом это позволяет классифицировать LTP на хеббийский, не-хеббийский и анти-хеббийский механизмы. Заимствовав свое название от Постулат Хебба, резюмируемый изречением, что «клетки, которые срабатывают вместе, соединяются вместе», Hebbian LTP требует одновременной пре- и постсинаптической деполяризации для его индукции.[25] Non-Hebbian LTP тип LTP, не требующий такой одновременной деполяризации пре- и постсинаптических клеток; Пример этого происходит в мшистых волокнах гиппокампа.[26] Частный случай нехеббийского LTP, антихеббийский LTP явно требует одновременной пресинаптической деполяризации и относительной постсинаптической гиперполяризации для ее индукции.[27]
Благодаря своей предсказуемой организации и легко индуцируемой LTP, гиппокамп CA1 стал прототипом исследования LTP у млекопитающих. В частности, зависимый от рецептора NMDA LTP в гиппокампе взрослого CA1 является наиболее широко изученным типом LTP,[21] и поэтому является целью данной статьи.
Свойства
LTP, зависимый от рецептора NMDA, проявляет несколько свойств, в том числе специфичность входа, ассоциативность, кооперативность и устойчивость.
- Специфика ввода
- После индуцирования LTP в одном синапсе не распространяется на другие синапсы; скорее LTP конкретный ввод. Долгосрочная потенциация распространяется только на эти синапсы в соответствии с правилами ассоциативности и кооперативности. Однако специфичность входа LTP может быть неполной на коротких расстояниях.[нужна цитата ] Одна модель, объясняющая специфичность входных данных LTP, была представлена Фреем и Моррисом в 1997 году и называется синаптическая маркировка и захват гипотеза.[нужна цитата ]
- Ассоциативность
- Ассоциативность относится к наблюдению, что, когда слабая стимуляция одного пути недостаточна для индукции LTP, одновременная сильная стимуляция другого пути будет индуцировать LTP на обоих путях.[нужна цитата ]
- Сотрудничество
- ДП может быть вызвана либо сильным тетаническая стимуляция единственного пути к синапсу, или совместно через более слабую стимуляцию многих. Когда один путь в синапс стимулируется слабо, он вызывает недостаточную постсинаптическую деполяризацию для индукции LTP. Напротив, когда слабые стимулы применяются ко многим путям, которые сходятся на единственном участке постсинаптической мембраны, генерируемые индивидуальные постсинаптические деполяризации могут коллективно деполяризовать постсинаптическую клетку в достаточной степени, чтобы совместно индуцировать LTP. Синаптическая маркировка, обсуждаемая позже, может быть общим механизмом, лежащим в основе ассоциативности и кооперативности. Брюс МакНотон утверждает, что любое различие между ассоциативностью и кооперативностью строго семантическое.[28] Эксперименты, проведенные путем стимуляции множества отдельных дендритных шипов, показали, что синаптическая кооперативность всего лишь двух соседних дендритных шипов предотвращает длительная депрессия (LTD) разрешает только LTP.[29]
- Упорство
- LTP - это стойкийдлится от нескольких минут до многих месяцев, и именно эта настойчивость отличает LTP от других форм синаптическая пластичность.[30]
Ранняя фаза
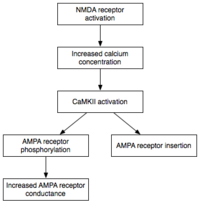

Обслуживание
В то время как индукция влечет за собой преходящий активация CaMKII и PKC, поддержание E-LTP (ранняя форма LTP) характеризуется их стойкий активация. На этом этапе ПКМз (Протеинкиназа Mζ ), не зависящий от кальция, становятся автономно активными. Следовательно, они способны осуществлять события фосфорилирования, лежащие в основе экспрессии E-LTP.[32]
Выражение
Фосфорилирование это химическая реакция, в которой небольшой фосфат группа добавляется к другой молекуле, чтобы изменить активность этой молекулы. Автономно активные CaMKII и PKC используют фосфорилирование для выполнения двух основных механизмов, лежащих в основе экспрессии E-LTP. Во-первых, и самое главное, они фосфорилируют существующие Рецепторы AMPA увеличить свою активность.[21] Во-вторых, они опосредуют или модулируют внедрение дополнительных рецепторов AMPA в постсинаптическую мембрану.[21] Важно отметить, что доставка рецепторов AMPA к синапсу во время E-LTP не зависит от синтез белка. Это достигается за счет наличия несинаптического пула рецепторов AMPA, прилегающего к постсинаптической мембране. Когда приходит соответствующий стимул, индуцирующий LTP, несинаптические рецепторы AMPA быстро попадают в постсинаптическую мембрану под влиянием протеинкиназ.[33] Как упоминалось ранее, рецепторы AMPA являются наиболее многочисленными рецепторами глутамата в головном мозге и опосредуют большую часть его возбуждающей активности. Повышая эффективность и количество рецепторов AMPA в синапсе, будущие возбуждающие стимулы генерируют более сильные постсинаптические ответы.
Хотя вышеприведенная модель E-LTP полностью описывает постсинаптические механизмы индукции, поддержания и экспрессии, дополнительный компонент экспрессии может происходить пресинаптически.[34] Одна из гипотез этого пресинаптического облегчения состоит в том, что постоянная активность CaMKII в постсинаптической клетке во время E-LTP может приводить к синтезу «ретроградного мессенджера», что обсуждается позже. Согласно этой гипотезе, вновь синтезированный мессенджер проходит через синаптическую щель от постсинаптической к пресинаптической клетке, приводя к цепочке событий, которые облегчают пресинаптическую реакцию на последующие стимулы. Такие события могут включать увеличение числа везикул нейромедиатора, вероятность высвобождения везикул или и то, и другое. В дополнение к ретроградному посланнику, лежащему в основе пресинаптического выражения в ранний LTP ретроградный мессенджер также может играть роль в экспрессии позднего LTP.
Поздняя фаза
Поздний LTP (L-LTP) является естественным продолжением E-LTP. В отличие от E-LTP, который не зависит от синтеза белка, L-LTP требует транскрипция гена[35] и синтез белка[36] в постсинаптической клетке. Существуют две фазы L-LTP: первая зависит от синтеза белка, а вторая зависит как от транскрипции гена, так и от синтеза белка.[31] Эти фазы иногда называют LTP2 и LTP3 соответственно, а E-LTP в рамках этой номенклатуры обозначается как LTP1.
Индукция
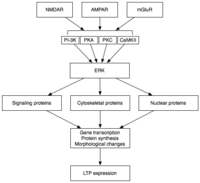
Поздняя ДП вызвана изменениями в экспрессия гена и синтез белка вызванный постоянной активацией протеинкиназ, активируемых во время E-LTP, таких как MAPK.[31][32][37] Фактически, MAPK, а именно киназа, регулируемая внеклеточными сигналами (ERK) подсемейство MAPK - может быть молекулярной связью между E-LTP и L-LTP, поскольку многие сигнальные каскады, участвующие в E-LTP, включая CaMKII и PKC, могут сходиться на ERK.[37] Недавние исследования показали, что индукция L-LTP может зависеть от совпадающих молекулярных событий, а именно активации PKA и притока кальция, которые сходятся на CRTC1 (TORC1), мощном коактиваторе транскрипции для белок, связывающий элемент ответа цАМФ (CREB).[38] Это требование молекулярного совпадения полностью объясняет ассоциативную природу LTP и, по-видимому, способность обучения.
Обслуживание
После активации ERK может фосфорилировать ряд цитоплазматических и ядерных молекул, что в конечном итоге приводит к синтезу белка и морфологическим изменениям, наблюдаемым в L-LTP.[31] Эти цитоплазматические и ядерные молекулы могут включать: факторы транскрипции такие как CREB.[32] ERK-опосредованные изменения активности факторов транскрипции могут запускать синтез белков, которые лежат в основе поддержания L-LTP. Одна такая молекула может быть протеинкиназа Mζ (PKMζ), постоянно активная киназа, синтез которой увеличивается после индукции LTP.[39][40] PKMζ является атипичной изоформой PKC, в которой отсутствует регуляторная субъединица, и поэтому она остается конститутивно активной.[39] В отличие от других киназ, которые опосредуют LTP, PKMζ активен не только в первые 30 минут после индукции LTP; скорее, PKMζ становится требованием для поддержания LTP только во время поздней фазы LTP.[39] PKMζ, таким образом, оказывается важным для сохранения памяти и, как ожидается, будет важен для поддержания Долгосрочная память. Действительно, введение ингибитора PKMζ в гиппокамп крысы приводит к ретроградная амнезия с неповрежденным краткосрочная память; PKMζ не играет роли в установлении кратковременной памяти.[40] Недавно было показано, что PKMζ лежит в основе поддержания L-LTP.[39][40] управляя перемещением и реорганизацией белков в синаптических каркасах, которые лежат в основе экспрессии L-LTP.[39] Даже совсем недавно трансгенные мыши, лишенные PKMζ, демонстрируют нормальный LTP, что ставит под сомнение необходимость PKMζ.[41]
Долговременная стабилизация синаптических изменений также определяется параллельным увеличением пре- и постсинаптических структур, таких как аксональный бутон, дендритный позвоночник и постсинаптическая плотность.[42]На молекулярном уровне увеличение количества белков постсинаптического каркаса. PSD-95 и Homer1c было показано, что коррелирует со стабилизацией расширения синапсов.[42]
Выражение
Идентичность только нескольких белков, синтезируемых во время L-LTP, известна. Независимо от их личности, считается, что они способствуют увеличению дендритный позвоночник количество, площадь поверхности и постсинаптическая чувствительность к нейротрансмиттеру, связанному с экспрессией L-LTP.[31] Последнее может быть частично вызвано усиленным синтезом рецепторов AMPA во время L-LTP.[31] Поздняя ДП также связана с пресинаптическим синтезом синаптотагмин и увеличение синаптический пузырек число, предполагающее, что L-LTP индуцирует синтез белка не только в постсинаптических клетках, но также и в пресинаптических клетках.[31] Как упоминалось ранее, для того, чтобы индукция постсинаптического LTP приводила к пресинаптическому синтезу белка, должна быть связь от постсинаптической к пресинаптической клетке. Это может происходить через синтез ретроградного посланника, который обсуждается позже.
Даже в исследованиях, ограниченных постсинаптическими событиями, исследователи не определили место синтеза белка, лежащего в основе L-LTP. В частности, неясно, происходит ли синтез белка в постсинаптическом Тело клетки или в его дендриты.[37] Несмотря на то, что наблюдал рибосомы (основные компоненты механизма синтеза белка) в дендритах еще в 1960-х годах преобладала точка зрения, что тело клетки является основным местом синтеза белка в нейронах.[37] Это рассуждение не подвергалось серьезному сомнению до 1980-х годов, когда исследователи сообщили о наблюдении синтеза белка в дендритах, связь которых с их клеточным телом была прервана.[37] Совсем недавно исследователи продемонстрировали, что этот тип локального синтеза белка необходим для некоторых типов LTP.[43][44]
Одна из причин популярности гипотезы локального синтеза белка заключается в том, что она обеспечивает возможный механизм специфичности, связанной с LTP.[37] В частности, если в основе L-LTP действительно лежит локальный синтез белка, только дендритные шипы, получающие LTP-индуцирующие стимулы, будут подвергаться LTP; потенциация не будет распространяться на соседние синапсы. Напротив, глобальный синтез белка, который происходит в теле клетки, требует, чтобы белки были доставлены во все области клетки, включая синапсы, которые не получили стимулы, индуцирующие LTP. В то время как локальный синтез белка обеспечивает механизм специфичности, глобальный синтез белка, по-видимому, напрямую его нарушает. Однако, как обсуждается позже, гипотеза синаптического тегирования успешно согласовывает глобальный синтез белка, специфичность синапсов и ассоциативность.
Ретроградная сигнализация
Ретроградная передача сигналов - это гипотеза, которая пытается объяснить, что, хотя LTP индуцируется и экспрессируется постсинаптически, некоторые данные свидетельствуют о том, что он экспрессируется также пресинаптически.[21][34][45] Гипотеза получила свое название потому, что нормальная синаптическая передача является направленным и идет от пресинаптической клетки к постсинаптической. Чтобы индукция происходила постсинаптически и частично выражалась пресинаптически, сообщение должно пройти от постсинаптической клетки к пресинаптической клетке в ретроградный (обратное направление. Оказавшись там, сообщение предположительно инициирует каскад событий, который приводит к пресинаптическому компоненту выражения, например, к повышенной вероятности везикула нейротрансмиттера выпуск.[46]
Ретроградная передача сигналов в настоящее время является предметом споров, поскольку некоторые исследователи не верят, что пресинаптические клетки вообще вносят вклад в экспрессию LTP.[21] Даже среди сторонников гипотезы есть разногласия по поводу личности посланника. Первые мысли были сосредоточены на оксид азота, в то время как самые последние свидетельства указывают на клеточная адгезия белки.[21]
Синаптическая маркировка
До того, как гипотеза локального синтеза белка получила значительную поддержку, существовало общее мнение, что синтез белка, лежащего в основе L-LTP, происходит в теле клетки. Кроме того, считалось, что продукты этого синтеза неспецифическим образом доставляются по всей клетке. Таким образом, возникла необходимость объяснить, как синтез белка может происходить в теле клетки без ущерба для специфичности входа LTP. Гипотеза синаптической маркировки пытается решить сложную проблему синтеза белков в теле клетки, но гарантирует, что они достигают только синапсов, которые получили стимулы, индуцирующие LTP.
Гипотеза синаптической маркировки предполагает, что «синаптическая метка» синтезируется в синапсах, которые получили LTP-индуцирующие стимулы, и что эта синаптическая метка может служить для захвата связанных с пластичностью белков, доставляемых по всей клетке из тела клетки.[47] Исследования LTP в морская улитка Аплизия калифорнийская подразумевают синаптическую маркировку как механизм специфичности входа LTP.[48][49] Есть некоторые свидетельства того, что при двух широко разделенных синапсах стимул, индуцирующий LTP в одном синапсе, запускает несколько сигнальных каскадов (описанных ранее), которые инициируют экспрессию генов в ядре клетки. В том же синапсе (но не в нестимулированном синапсе) локальный синтез белка создает недолговечную (менее трех часов) синаптическую метку. Продукты экспрессии генов распространяются по всей клетке по всему миру, но захватываются только синапсами, которые экспрессируют синаптический тег. Таким образом, потенцируется только синапс, получающий стимулы, индуцирующие LTP, демонстрируя специфичность входа LTP.
Гипотеза синаптических тегов может также учитывать ассоциативность и кооперативность LTP. Ассоциативность (увидеть Свойства ) наблюдается, когда один синапс возбуждается LTP-индуцирующей стимуляцией, тогда как отдельный синапс стимулируется слабо. Принимая во внимание, что можно было ожидать, что только сильно стимулированный синапс подвергнется LTP (поскольку одной слабой стимуляции недостаточно для индукции LTP в любом синапсе), и то и другое на самом деле синапсы подвергаются ДП. Хотя слабые стимулы не могут вызвать синтез белка в теле клетки, они могут стимулировать синтез синаптической метки. Одновременная сильная стимуляция отдельного пути, способного индуцировать синтез белков тела клетки, затем может вызвать выработку белков, связанных с пластичностью, которые доставляются по всей клетке. Когда оба синапса экспрессируют синаптическую метку, оба будут захватывать белковые продукты, что приводит к экспрессии LTP как в сильно стимулированном, так и в слабостимулированном путях.
Кооперативность наблюдается, когда два синапса активируются слабыми стимулами, неспособными индуцировать LTP при индивидуальной стимуляции. Но при одновременной слабой стимуляции оба синапса кооперативно подвергаются ДП. Синаптическая маркировка не объясняет, как множественные слабые стимулы могут привести к коллективному стимулу, достаточному для индукции ДП (это объясняется постсинаптическим суммированием ВПСП, описанным ранее). Скорее, синаптическая маркировка объясняет способность слабо стимулированных синапсов, ни один из которых не способен независимо генерировать LTP, получать продукты синтеза белка, инициированного коллективно. Как и раньше, это может быть достигнуто посредством синтеза локальной синаптической метки после слабой синаптической стимуляции.
Модуляция
| Модулятор | Цель |
|---|---|
| β-адренорецептор | цАМФ, усиление MAPK |
| Синтаза оксида азота | Гуанилилциклаза, PKG, NMDAR |
| Рецепторы допамина | цАМФ, усиление MAPK |
| Метаботропный рецептор глутамата | PKC, усиление MAPK |
Как описано ранее, молекулы, лежащие в основе LTP, можно классифицировать как медиаторы или модуляторы. Медиатор LTP представляет собой молекулу, такую как рецептор NMDA или кальций, присутствие и активность которой необходимы для генерации LTP почти во всех условиях. Напротив, модулятор - это молекула, которая может изменять LTP, но не важна для его образования или экспрессии.[21]
В дополнение к сигнальным путям, описанным выше, LTP в гиппокампе может изменяться множеством модуляторов. Например, стероидный гормон эстрадиол может усиливать LTP за счет фосфорилирования CREB и последующего дендритный позвоночник рост.[50] Дополнительно, β-адренорецептор агонисты, такие как норэпинефрин может изменять зависимую от синтеза белка позднюю фазу LTP.[51] Синтаза оксида азота активность может также привести к последующей активации гуанилилциклазы и PKG.[52] Аналогичным образом активация дофаминовые рецепторы может усиливать LTP через сигнальный путь цАМФ / PKA.[53][54]
Отношение к поведенческой памяти
Хотя долгосрочное усиление синапсов в клеточной культуре, по-видимому, обеспечивает элегантный субстрат для обучения и памяти, вклад LTP в поведенческое обучение, то есть обучение на уровне всего организма, нельзя просто экстраполировать из in vitro исследования. По этой причине значительные усилия были направлены на то, чтобы установить, является ли LTP требованием для обучения и памяти у живых животных. Из-за этого LTP также играет решающую роль в обработка страха.
Пространственная память

В 1986 году Ричард Моррис предоставил одни из первых доказательств того, что LTP действительно требуется для формирования воспоминаний. in vivo.[55] Он проверил пространственная память крыс путем фармакологической модификации их гиппокампа, структуры мозга, роль которой в пространственном обучении хорошо известна. Крыс дрессировали на Водный лабиринт Морриса, задача пространственной памяти, в которой крысы плавают в бассейне с мутной водой, пока не обнаружат платформу, скрытую под ее поверхностью. Ожидается, что во время этого упражнения нормальные крысы будут ассоциировать расположение скрытой платформы с заметными сигналами, размещенными в определенных местах по окружности лабиринта. После тренировки гиппокамп одной группы крыс был купан в блокаторе рецепторов NMDA. APV, а другая группа служила контроль. Затем обе группы были подвергнуты задаче на пространственную память в водном лабиринте. Крысы в контрольной группе смогли найти платформу и покинуть бассейн, в то время как производительность крыс, получавших APV, была значительно снижена. Более того, когда срезы гиппокампа были взяты у обеих групп, LTP легко индуцировался в контроле, но не мог быть индуцирован в мозге крыс, получавших APV. Это дало ранние доказательства того, что рецептор NMDA - и, в более широком смысле, LTP - необходим по крайней мере для некоторых типов обучения и памяти.
Так же, Сусуму Тонегава продемонстрировали в 1996 г., что область CA1 гиппокампа имеет решающее значение для формирования пространственной памяти у живых мышей.[56] Так называемые разместить клетки расположенные в этой области, становятся активными только тогда, когда крыса находится в определенном месте, называемом разместить поле - в окружающей среде. Поскольку эти поля мест распределены по окружающей среде, одна из интерпретаций состоит в том, что группы клеток мест формируют карты в гиппокампе. Точность этих карт определяет, насколько хорошо крыса узнает об окружающей среде и, следовательно, насколько хорошо она может ориентироваться в ней. Тонегава обнаружил, что за счет нарушения рецептора NMDA, в частности путем генетического удаления субъединицы NR1 в области CA1, генерируемые поля мест были значительно менее специфичными, чем поля контроля. То есть мыши создавали неправильные пространственные карты, когда их рецепторы NMDA были повреждены. Как и ожидалось, эти мыши очень плохо справлялись с пространственными задачами по сравнению с контрольной группой, что дополнительно подтверждает роль LTP в пространственном обучении.
Также было показано, что повышенная активность рецептора NMDA в гиппокампе приводит к усилению LTP и общему улучшению пространственного обучения. В 1999 году Тан и другие. получили линию мышей с усиленной функцией рецептора NMDA за счет сверхэкспрессии NR2B субъединица в гиппокампе.[57][58] В результате появились умные мыши, прозванные «мышками Дуги» в честь вымышленного чудо-доктора. Дуги Хаузер, имели более крупный LTP и преуспели в задачах пространственного обучения, что подчеркивало важность LTP в формировании зависимых от гиппокампа воспоминаний.
Тормозное избегание
В 2006 г. Джонатан Уитлок и его коллеги сообщили о серии экспериментов, которые предоставили, возможно, наиболее убедительные доказательства роли LTP в поведенческой памяти, утверждая, что для вывода о том, что LTP лежит в основе поведенческого обучения, два процесса должны имитировать и перекрывать друг друга.[59] Используя парадигму обучения тормозящего избегания, исследователи тренировали крыс в двухкамерном аппарате со светлой и темной камерами, последняя была оснащена устройством, которое наносило крысе удар ногой при входе. Анализ синапсов гиппокампа CA1 показал, что тренировка тормозящего избегания индуцирует in vivo Фосфорилирование рецептора AMPA того же типа, что и у LTP in vitro; то есть тренировка ингибирующего избегания имитировала LTP. Кроме того, синапсы, потенцированные во время тренировки, не могли быть дополнительно усилены экспериментальными манипуляциями, которые в противном случае индуцировали бы LTP; то есть тренировка с ингибирующим избеганием блокировала LTP. В ответ на статью Тимоти Блисс и его коллеги отметили, что эти и связанные с ними эксперименты «существенно продвигают аргументы в пользу LTP как нейронного механизма памяти».[60]
Клиническое значение
Роль LTP в заболевании менее ясна, чем его роль в основных механизмах синаптическая пластичность. Однако изменения в LTP могут способствовать ряду неврологические заболевания, в том числе депрессия, болезнь Паркинсона, эпилепсия, и невропатическая боль.[61] Нарушение LTP также может играть роль в Болезнь Альцгеймера и наркотическая зависимость.
Болезнь Альцгеймера

LTP привлекла большое внимание среди тех, кто учится Болезнь Альцгеймера (AD), а нейродегенеративное заболевание что вызывает заметное снижение когнитивных функций и слабоумие. Большая часть этого ухудшения происходит в связи с дегенеративными изменениями гиппокампа и других медиальная височная доля конструкции. Из-за хорошо известной роли гиппокампа в LTP, некоторые предположили, что снижение когнитивных функций, наблюдаемое у людей с AD, может быть результатом нарушения LTP.
В обзоре литературы за 2003 год Роуэн и другие. предложил одну модель того, как LTP может быть затронут в AD.[62] AD является результатом, по крайней мере частично, неправильной обработки белок-предшественник амилоида (ПРИЛОЖЕНИЕ). Результатом этой ненормальной обработки является накопление фрагментов этого белка, называемых амилоид β (Aβ). Aβ существует как в растворимой, так и в фибриллярной формах. Неправильная обработка APP приводит к накоплению растворимого Aβ, который, согласно гипотезе Роуэна, нарушает LTP в гиппокампе и может привести к когнитивному снижению, наблюдаемому на ранних этапах AD.
AD также может нарушать LTP посредством механизмов, отличных от Aβ. Например, одно исследование показало, что фермент PKMζ накапливается в нейрофибриллярные сплетения, которые являются патологическим маркером БА. PKMζ - это фермент, имеющий решающее значение для поддержание позднего ДП.[63]
Наркотическая зависимость
Исследования в области лекарство от наркозависимости также недавно обратила свое внимание на LTP из-за гипотезы о том, что наркотическая зависимость представляет собой мощную форму обучения и памяти.[64] Зависимость - это сложное нейроповеденческое явление, затрагивающее различные части мозга, такие как вентральная тегментальная область (VTA) и прилежащее ядро (NAc). Исследования показали, что синапсы VTA и NAc способны подвергаться LTP.[64] и что этот ДП может быть ответственным за поведение, характеризующее зависимость.[65]
Смотрите также
- Нейропластичность
- Ремоделирование актина нейронов
- Транскраниальная стимуляция постоянным током
- Посттетаническое потенцирование
использованная литература
- ^ Парадизо М.А., Медведь М.Ф., Коннорс Б.В. (2007). Неврология: изучение мозга. Хагерствон, доктор медицины: Липпинкотт Уильямс и Уилкинс. п.718. ISBN 978-0-7817-6003-4.
- ^ а б Кук SF, Bliss TV (июль 2006 г.). «Пластичность центральной нервной системы человека». Мозг. 129 (Pt 7): 1659–73. Дои:10.1093 / мозг / awl082. PMID 16672292.
- ^ а б Bliss TV, Collingridge GL (январь 1993 г.). «Синаптическая модель памяти: долговременная потенциация в гиппокампе». Природа. 361 (6407): 31–9. Bibcode:1993Натура 361 ... 31Б. Дои:10.1038 / 361031a0. PMID 8421494. S2CID 4326182.
- ^ Уильямс Р. У., Херруп К. (1988). «Контроль числа нейронов». Ежегодный обзор нейробиологии. 11 (1): 423–53. Дои:10.1146 / annurev.ne.11.030188.002231. PMID 3284447.
- ^ а б Сантьяго Ри (1894). "Крооновская лекция: тонкая структура нервных центров". Труды Лондонского королевского общества. 55 (331–335): 444–468. Bibcode:1894RSPS ... 55..444C. Дои:10.1098 / rspl.1894.0063.
- ^ Хебб Д. (1949). Организация поведения: НЕЙРОПСИХОЛОГИЧЕСКАЯ ТЕОРИЯ. Нью-Йорк: ДЖОН ВИЛИ, если SONS, Inc. ISBN 978-0805843002.
- ^ Хебб Д.О. (1949). Организация поведения: нейропсихологическая теория. Нью-Йорк: Джон Вили. ISBN 978-0-471-36727-7.
- ^ Кандел Э. Р., Тауц Л. (ноябрь 1965 г.). «Гетеросинаптическое облегчение нейронов брюшного ганглия Aplysia depilans». Журнал физиологии. 181 (1): 1–27. Дои:10.1113 / jphysiol.1965.sp007742. ЧВК 1357435. PMID 5866283.
- ^ Патихис Л. (октябрь 2018 г.). «Историческое значение открытия долгосрочного потенцирования: обзор и оценка для неспециалистов». Американский журнал психологии. 131 (3): 369–80. Дои:10.5406 / amerjpsyc.131.3.0369.
- ^ а б Лёмо Т. (апрель 2003 г.). «Открытие длительного потенцирования». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 358 (1432): 617–20. Дои:10.1098 / rstb.2002.1226. ЧВК 1693150. PMID 12740104.
- ^ Лемо Т. (1966). «Частотное потенцирование возбуждающей синаптической активности в зубчатой области гиппокампа». Acta Physiologica Scandinavica. 68 (Приложение 277): 128.
- ^ а б Bliss TV, Lomo T (июль 1973). «Длительное усиление синаптической передачи в зубчатой области анестезированного кролика после стимуляции перфорантного пути». Журнал физиологии. 232 (2): 331–56. Дои:10.1113 / jphysiol.1973.sp010273. ЧВК 1350458. PMID 4727084.
- ^ а б Bliss TV, Гарднер-Медвин AR (июль 1973 г.). «Длительное усиление синаптической передачи в зубчатой области кролика без анестезии после стимуляции перфорантного пути». Журнал физиологии. 232 (2): 357–74. Дои:10.1113 / jphysiol.1973.sp010274. ЧВК 1350459. PMID 4727085.
- ^ Хотя термин «долгосрочное потенцирование» однажды появился в оригинальной статье Блисса и Лёмо, он не был официально предложен для этого явления до работы Дугласа и Годдарда.
- ^ Дуглас Р.М., Годдард Г.В. (март 1975 г.). «Долгосрочное усиление синапса перфорантного пути гранулярных клеток в гиппокампе крысы». Исследование мозга. 86 (2): 205–15. Дои:10.1016/0006-8993(75)90697-6. PMID 163667. S2CID 43260928.
- ^ Андерсен П. (апрель 2003 г.). «Прелюдия к длительному потенцированию». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 358 (1432): 613–5. Дои:10.1098 / rstb.2002.1232. ЧВК 1693144. PMID 12740103.
- ^ McEachern JC, Shaw CA (июнь 1996 г.). «Альтернатива ортодоксальности LTP: модель континуума пластичности и патологии». Исследование мозга. Обзоры исследований мозга. 22 (1): 51–92. Дои:10.1016/0165-0173(96)00006-9. PMID 8871785. S2CID 41680613. 8871785.
- ^ Медведь М.Ф. (1996). «Синаптическая основа для хранения памяти в коре головного мозга». Труды Национальной академии наук. 93 (24): 13453–13459. Bibcode:1996PNAS ... 9313453B. Дои:10.1073 / пнас.93.24.13453. ЧВК 33630. PMID 8942956.
- ^ Уардуз М., Састри Б.Р. (2000). «Механизмы, лежащие в основе LTP ингибиторной синаптической передачи в глубоких ядрах мозжечка». Журнал нейрофизиологии. 84 (3): 1414–1421. Дои:10.1152 / jn.2000.84.3.1414. PMID 10980014.
- ^ Clugnet MC, LeDoux JE (август 1990 г.). «Синаптическая пластичность в схемах кондиционирования страха: индукция LTP в латеральном ядре миндалевидного тела путем стимуляции медиального коленчатого тела» (PDF). Журнал неврологии. 10 (8): 2818–24. Дои:10.1523 / JNEUROSCI.10-08-02818.1990. ЧВК 6570263. PMID 2388089.
- ^ а б c d е ж г час я Маленка Р.С., Медведь М.Ф. (сентябрь 2004 г.). «ЛТП и ООО: позор богатства». Нейрон. 44 (1): 5–21. Дои:10.1016 / j.neuron.2004.09.012. PMID 15450156. S2CID 79844.
- ^ Ясуда Х., Барт А.Л., Стеллваген Д., Маленка Р.К. (январь 2003 г.). «Переключатель развития в сигнальных каскадах для индукции LTP». Природа Неврология. 6 (1): 15–6. Дои:10.1038 / nn985. PMID 12469130. S2CID 28913342.
- ^ Collingridge GL, Kehl SJ, McLennan H (январь 1983 г.). «Возбуждающие аминокислоты в синаптической передаче в коллатерально-комиссуральном пути Шаффера гиппокампа крысы». Журнал физиологии. 334: 33–46. Дои:10.1113 / jphysiol.1983.sp014478. ЧВК 1197298. PMID 6306230.
- ^ Харрис EW, Котман CW (сентябрь 1986). «Долгосрочное усиление реакции мшистых волокон морских свинок не блокируется антагонистами N-метил-D-аспартата». Письма о неврологии. 70 (1): 132–7. Дои:10.1016/0304-3940(86)90451-9. PMID 3022192. S2CID 42647125.
- ^ Вигстрём Х, Густафссон Б. (1986). «Постсинаптический контроль долговременной потенциации гиппокампа». Journal de Physiologie. 81 (4): 228–36. PMID 2883309.
- ^ Городской Н.Н., Баррионуево Г (июль 1996 г.). «Индукция долгосрочной потенциации геббийских и негеббийских мшистых волокон с помощью различных паттернов высокочастотной стимуляции». Журнал неврологии. 16 (13): 4293–9. Дои:10.1523 / JNEUROSCI.16-13-04293.1996. ЧВК 6579001. PMID 8753890.
- ^ Куллманн Д.М., Ламса К. (март 2008 г.). «Роль различных глутаматных рецепторов в индукции долгосрочной потенциации против Хебба». Журнал физиологии. 586 (6): 1481–6. Дои:10.1113 / jphysiol.2007.148064. ЧВК 2375711. PMID 18187472.
- ^ Макнотон Б.Л. (апрель 2003 г.). «Долгосрочное потенцирование, кооперативность и сборки клеток Хебба: личная история». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 358 (1432): 629–34. Дои:10.1098 / rstb.2002.1231. ЧВК 1693161. PMID 12740107.
- ^ Tazerart S, Mitchell DE, Miranda-Rottmann S, Araya R (август 2020 г.). «Правило пластичности дендритных шипов, зависящее от времени спайков». Nature Communications. 11 (1): 4276. Bibcode:2020NatCo..11.4276T. Дои:10.1038 / s41467-020-17861-7. ЧВК 7449969. PMID 32848151.
- ^ Авраам WC (апрель 2003 г.). "Как долго продлится долгосрочное потенцирование?". Философские труды Лондонского королевского общества. Серия B, Биологические науки. 358 (1432): 735–44. Дои:10.1098 / rstb.2002.1222. ЧВК 1693170. PMID 12740120.
- ^ а б c d е ж г час Линч М.А. (январь 2004 г.). «Долговременное потенцирование и память». Физиологические обзоры. 84 (1): 87–136. Дои:10.1152 / физрев.00014.2003. PMID 14715912.
- ^ а б c d Свэтт Дж. Д. (1999). «К молекулярному объяснению долгосрочного потенцирования». Обучение и память. 6 (5): 399–416. Дои:10,1101 / лм. 6.5.399. PMID 10541462.
- ^ Малинов Р. (апрель 2003 г.). «Торговля рецепторами AMPA и долгосрочное потенцирование». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 358 (1432): 707–14. Дои:10.1098 / rstb.2002.1233. ЧВК 1693162. PMID 12740116.
- ^ а б Emptage NJ, Reid CA, Fine A, Bliss TV (июнь 2003 г.). «Оптический квантовый анализ выявляет пресинаптический компонент LTP в гиппокампе-ассоциативных синапсах Шаффера». Нейрон. 38 (5): 797–804. Дои:10.1016 / S0896-6273 (03) 00325-8. PMID 12797963. S2CID 13629691.
- ^ Frey U, Frey S, Schollmeier F, Krug M (февраль 1996 г.). «Влияние актиномицина D, ингибитора синтеза РНК, на долгосрочное потенцирование нейронов гиппокампа крысы in vivo и in vitro». Журнал физиологии. 490. 490 (Pt 3) (Pt 3): 703–11. Дои:10.1113 / jphysiol.1996.sp021179. ЧВК 1158708. PMID 8683469.
- ^ Фрей У, Круг М., Рейманн К.Г., Маттис Х. (июнь 1988 г.). «Анизомицин, ингибитор синтеза белка, блокирует поздние фазы LTP-феноменов в области CA1 гиппокампа in vitro». Исследование мозга. 452 (1–2): 57–65. Дои:10.1016 / 0006-8993 (88) 90008-X. PMID 3401749. S2CID 39245231.
- ^ а б c d е ж Келлехер Р.Дж., Говиндараджан А., Тонегава С. (сентябрь 2004 г.). «Трансляционные регуляторные механизмы в стойких формах синаптической пластичности». Нейрон. 44 (1): 59–73. Дои:10.1016 / j.neuron.2004.09.013. PMID 15450160. S2CID 1511103.
- ^ Ковач К.А., Стуллет П., Стейнманн М., До К.К., Магистретти П.Дж., Халфон О., Кардино-младший (март 2007 г.). «TORC1 - это кальций- и цАМФ-чувствительный детектор совпадений, участвующий в долговременной синаптической пластичности гиппокампа». Труды Национальной академии наук Соединенных Штатов Америки. 104 (11): 4700–5. Bibcode:2007ПНАС..104.4700К. Дои:10.1073 / pnas.0607524104. ЧВК 1838663. PMID 17360587.
- ^ а б c d е Серрано П., Яо Й, Сактор Т.С. (февраль 2005 г.). «Устойчивое фосфорилирование протеинкиназой Mzeta поддерживает долгосрочное усиление на поздней фазе». Журнал неврологии. 25 (8): 1979–84. Дои:10.1523 / JNEUROSCI.5132-04.2005. ЧВК 6726070. PMID 15728837.
- ^ а б c Пасталкова Е., Серрано П., Пинхасова Д., Уоллес Е., Фентон А.А., Sacktor TC (август 2006 г.). «Хранение пространственной информации механизмом обслуживания LTP». Наука. 313 (5790): 1141–4. Bibcode:2006Научный ... 313.1141П. CiteSeerX 10.1.1.453.2136. Дои:10.1126 / science.1128657. PMID 16931766. S2CID 7260010.
- ^ Volk LJ, Bachman JL, Johnson R, Yu Y, Huganir RL (январь 2013 г.). «PKM-ζ не требуется для синаптической пластичности гиппокампа, обучения и памяти». Природа. 493 (7432): 420–3. Bibcode:2013Натура.493..420В. Дои:10.1038 / природа11802. ЧВК 3830948. PMID 23283174.
- ^ а б Мейер Д., Бонхёффер Т., Шойс В. (апрель 2014 г.). «Равновесие и стабильность синаптических структур при синаптической пластичности». Нейрон. 82 (2): 430–43. Дои:10.1016 / j.neuron.2014.02.031. PMID 24742464.
- ^ Канг Х., Шуман Э.М. (сентябрь 1996 г.). «Необходимость в локальном синтезе белка в нейротрофин-индуцированной синаптической пластичности гиппокампа». Наука. 273 (5280): 1402–6. Bibcode:1996Научный ... 273.1402K. Дои:10.1126 / science.273.5280.1402. PMID 8703078. S2CID 38648558.
- ^ Стюард О., Уорли П.Ф. (июнь 2001 г.). «Клеточный механизм для нацеливания вновь синтезированных мРНК на синаптические сайты на дендритах». Труды Национальной академии наук Соединенных Штатов Америки. 98 (13): 7062–8. Bibcode:2001PNAS ... 98.7062S. Дои:10.1073 / pnas.131146398. ЧВК 34623. PMID 11416188.
- ^ Павлидис П., Монтгомери Дж., Мэдисон Д.В. (июнь 2000 г.). «Активность пресинаптической протеинкиназы поддерживает долгосрочную потенциацию в синапсах между отдельными нейронами гиппокампа». Журнал неврологии. 20 (12): 4497–505. Дои:10.1523 / JNEUROSCI.20-12-04497.2000. ЧВК 6772468. PMID 10844019.
- ^ Захаренко С.С., Паттерсон С.Л., Драгацис И., Цейтлин С.О., Зигельбаум С.А., Кандел Е.Р., Морозов А. (сентябрь 2003 г.). «Пресинаптический BDNF необходим для пресинаптического, но не постсинаптического компонента LTP в синапсах CA1-CA3 гиппокампа». Нейрон. 39 (6): 975–90. Дои:10.1016 / S0896-6273 (03) 00543-9. PMID 12971897. S2CID 7117772.
- ^ Фрей У., Моррис Р.Г. (февраль 1997 г.). «Синаптическая маркировка и долгосрочное потенцирование». Природа. 385 (6616): 533–6. Bibcode:1997Натура.385..533F. Дои:10.1038 / 385533a0. PMID 9020359. S2CID 4339789.
- ^ Мартин К.С., Касадио А., Чжу Х., Япин Э., Роуз Дж. К., Чен М., Бейли С.Х., Кандел ER (декабрь 1997 г.). «Синапс-специфичное, долгосрочное облегчение сенсорной аплизии моторных синапсов: функция локального синтеза белка в памяти». Ячейка. 91 (7): 927–38. Дои:10.1016 / S0092-8674 (00) 80484-5. PMID 9428516. S2CID 16423304.
- ^ Casadio A, Martin KC, Giustetto M, Zhu H, Chen M, Bartsch D, Bailey CH, Kandel ER (октябрь 1999 г.). «Временная форма CREB-опосредованного долгосрочного облегчения, охватывающая все нейроны, может быть стабилизирована в конкретных синапсах путем локального синтеза белка». Ячейка. 99 (2): 221–37. Дои:10.1016 / S0092-8674 (00) 81653-0. PMID 10535740.
- ^ Сигал М., Мерфи Д.Д. (1999). «Активация CREB опосредует пластичность культивируемых нейронов гиппокампа». Нейронная пластичность. 6 (3): 1–7. Дои:10.1155 / NP.1998.1. ЧВК 2565317. PMID 9920677.
- ^ Straube T, Фрей JU (2003). «Вовлечение бета-адренорецепторов в зависимую от синтеза белка позднюю долгосрочную потенциацию (LTP) в зубчатой извилине свободно движущихся крыс: критическая роль силы индукции LTP». Неврология. 119 (2): 473–9. Дои:10.1016 / S0306-4522 (03) 00151-9. PMID 12770561. S2CID 23436714.
- ^ Лу Ю.Ф., Кандел Э.Р., Хокинс Р.Д. (декабрь 1999 г.). «Передача сигналов оксида азота способствует поздней фазе фосфорилирования LTP и CREB в гиппокампе». Журнал неврологии. 19 (23): 10250–61. Дои:10.1523 / JNEUROSCI.19-23-10250.1999. ЧВК 6782403. PMID 10575022.
- ^ Фрей Ю., Мэттис Х., Рейманн К.Г., Мэттис Х. (август 1991 г.). «Влияние блокады дофаминергического рецептора D1 во время тетанизации на экспрессию долговременной потенциации в области СА1 крысы in vitro». Письма о неврологии. 129 (1): 111–4. Дои:10.1016/0304-3940(91)90732-9. PMID 1833673. S2CID 45084596.
- ^ Отмахова Н.А., Лисман Ю.Е. (декабрь 1996 г.). «Активация дофаминового рецептора D1 / D5 увеличивает величину ранней долгосрочной потенциации в синапсах СА1 гиппокампа». Журнал неврологии. 16 (23): 7478–86. Дои:10.1523 / JNEUROSCI.16-23-07478.1996. ЧВК 6579102. PMID 8922403.
- ^ Моррис Р.Г., Андерсон Э., Линч Г.С., Бодри М. (1986). «Избирательное нарушение обучения и блокада долгосрочного потенцирования антагонистом рецептора N-метил-D-аспартата, AP5». Природа. 319 (6056): 774–6. Bibcode:1986Натура.319..774М. Дои:10.1038 / 319774a0. PMID 2869411. S2CID 4356601.
- ^ McHugh TJ, Blum KI, Tsien JZ, Tonegawa S, Wilson MA (декабрь 1996 г.). «Нарушение представления пространства в гиппокампе у CA1-специфичных мышей с нокаутом NMDAR1». Ячейка. 87 (7): 1339–49. Дои:10.1016 / S0092-8674 (00) 81828-0. PMID 8980239. S2CID 5131226.
- ^ Тан Ю.П., Симидзу Э., Дубе Г.Р., Рэмпон С., Керчнер Г.А., Чжуо М., Лю Дж., Цзянь Дж. З. (1999). «Генетическое улучшение обучения и памяти у мышей». Природа. 401 (6748): 63–69. Bibcode:1999 Натур 401 ... 63 т. Дои:10.1038/43432. PMID 10485705. S2CID 481884.
- ^ Тан И, Ван Х, Фэн Р, Чжин М., Цзянь Дж (2001). «Дифференциальные эффекты обогащения на обучение и функцию памяти у трансгенных мышей NR2B». Нейрофармакология. 41 (6): 779–90. Дои:10.1016 / S0028-3908 (01) 00122-8. PMID 11640933. S2CID 23602265.
- ^ Whitlock JR, Heynen AJ, Shuler MG, Bear MF (август 2006 г.). «Обучение вызывает долгосрочное потенцирование в гиппокампе». Наука. 313 (5790): 1093–7. Bibcode:2006Sci ... 313.1093W. Дои:10.1126 / science.1128134. PMID 16931756. S2CID 612352.
- ^ Bliss TV, Collingridge GL, Laroche S (август 2006 г.). «Неврология. ZAP и ZIP, история, которую нужно забыть». Наука. 313 (5790): 1058–9. Дои:10.1126 / наука.1132538. PMID 16931746. S2CID 27735098.
- ^ Кук SF, Bliss TV (июль 2006 г.). «Пластичность центральной нервной системы человека». Мозг. 129 (Pt 7): 1659–73. Дои:10.1093 / мозг / awl082. PMID 16672292.
- ^ а б Роуэн М.Дж., Клюбин И., Каллен В.К., Анвил Р. (апрель 2003 г.). «Синаптическая пластичность в животных моделях ранней болезни Альцгеймера». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 358 (1432): 821–8. Дои:10.1098 / rstb.2002.1240. ЧВК 1693153. PMID 12740129.
- ^ Crary JF, Shao CY, Mirra SS, Hernandez AI, Sacktor TC (апрель 2006 г.). «Атипичная протеинкиназа С при нейродегенеративном заболевании I: PKMzeta объединяется с лимбическими нейрофибриллярными клубками и рецепторами AMPA при болезни Альцгеймера». Журнал невропатологии и экспериментальной неврологии. 65 (4): 319–26. Дои:10.1097 / 01.jnen.0000218442.07664.04. PMID 16691113.
- ^ а б Кауэр Я.А., Маленка RC (ноябрь 2007 г.). «Синаптическая пластичность и зависимость». Обзоры природы. Неврология. 8 (11): 844–58. Дои:10.1038 / номер 2234. PMID 17948030. S2CID 38811195.
- ^ Wolf ME (август 2003 г.). «LTP может вызвать зависимость». Молекулярные вмешательства. 3 (5): 248–52. Дои:10.1124 / mi 3.5.248. PMID 14993438.
дальнейшее чтение
- Блисс Т., Коллингридж Г., Моррис Р. (2004). Долгосрочное потенцирование: совершенствование нейробиологии на 30 лет. Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-853030-5.
внешние ссылки
- Исследователи предоставляют первые доказательства механизма обучения, а PhysOrg.com отчет об исследовании 2006 г., проведенном Bear и коллегами.
- Короткий документальный видеофильм о мышах Дуги. (Реальный игрок формат)
- «Умная мышь», телесериал Quantum ABC о мышах Doogie.
- Долгосрочный + потенциал в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)