Ахелозавр - Achelousaurus - Wikipedia
| Ахелозавр | |
|---|---|
 | |
| Череп образец голотипа MOR 485 (с реконструированными частями синего цвета), на Музей Скалистых гор, Монтана | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Заказ: | †Орнитишия |
| Семья: | †Ceratopsidae |
| Подсемейство: | †Centrosaurinae |
| Племя: | †Пахиринозаврини |
| Clade: | †Пахиростра |
| Род: | †Ахелозавр Сампсон, 1994 |
| Разновидность: | †A. horneri |
| Биномиальное имя | |
| †Ахелозавр горнери Сампсон, 1994 | |
Ахелозавр (/əˌkялoʊˈsɔːrəs/ или же: /ˌækɪˌлoʊəˈsɔːrəs/[1]) это род из центрозаврин цератопсид динозавр которые жили во время Поздний меловой период из того, что сейчас Северная Америка, около 74,2 миллиона лет назад. Первые окаменелости Ахелозавр были собраны в Монтана в 1987 году командой во главе с Джек Хорнер, больше находок было сделано в 1989 г. В 1994 г. Ахелозавр горнери был описан и назван Скотт Д. Сэмпсон; общее название означает "Ахелой ящерица ", по отношению к греческому богу Ахелой, а конкретное имя относится к Хорнеру. Род известен по нескольким экземплярам, состоящим в основном из черепного материала отдельных особей, от молодых до взрослых.
Большой центрозаврин, Ахелозавр предположительно был около 6 м (20 футов) в длину и весил около 3 т (3,3 коротких тонны). Как цератопс, он ходил на четвереньках, имел короткий хвост и большую голову с крючковатым клювом. На задней части черепа у него была костистая оборка на шее с парой длинных шипов, загибающихся наружу. Взрослый Ахелозавр имели грубые выступы (округлые выступы) над глазами и на морде, тогда как у других центрозавринов рога часто располагались в том же положении. Эти боссы были покрыты толстым слоем кератин, но их точная форма в жизни неизвестна. По мнению некоторых ученых, боссов использовали в драках, когда животные бодали друг друга головами, а также для демонстрации.
В рамках Цератопсия, Ахелозавр лежит в пределах клады Пахиростра (или «толстые рыла»). Было высказано предположение, что это был прямой потомок аналогичного рода Эйниозавр (который имел шипы, но не имел боссов) и прямой предок Пахиринозавр (у которых были более крупные боссы). Первые два рода будут переходные формы, развиваясь через анагенез из Стиракозавр. Об этой теории велись споры, и более поздние открытия показали, что Ахелозавр тесно связан с Пахиринозавр в группе Пахиринозаврини. Ахелозавр известно из Формирование двух лекарств и жил на островном континенте Ларамидия. Как цератопсианец, Ахелозавр был бы травоядным и, похоже, имел высокий скорость метаболизма, хотя и ниже, чем у современных млекопитающих и птиц.
История открытия
Экспедиции Хорнера на оползень-бьютт

Все известные Ахелозавр образцы были извлечены из Формирование двух лекарств в Glacier County, Монтана во время раскопок, проведенных Музей Скалистых гор, в котором до сих пор хранятся образцы. Открытия произошли в результате случайной цепочки событий.[2] Весной 1985 г. палеонтолог Джон «Джек» Р. Хорнер был проинформирован, что ему больше не будет разрешено использовать Willow Creek сайт, где он изучал Майасаура Яичная гора гнездовая колония на шесть лет.[3] Он уже тщательно подготовился к новому полевому сезону и внезапно был вынужден искать альтернативное место. Хорнера всегда интересовали полевые дневники Чарльз Уитни Гилмор которые сообщили об открытии яиц динозавров в Лэндслайд Бьютт в 1928 году, но никогда не публиковали их.[4] В этой местности Гилмор нанял Джордж Фрайер Штернберг раскопать скелеты рогатых динозавров Брахицератопс и Стиракозавр оватус.[5] Тем летом Хорнер получил разрешение Совет племен черноногих индейцев поискать окаменелости на Лендслайд Бьютт, который является частью Резервация черноногих индейцев; это было первое палеонтологическое исследование здесь с 1920-х годов. В августе 1985 года соратник Хорнера Боб Макела обнаружил богатый участок окаменелостей на земле фермера Рики Рейгана, который назывался «Карьер динозавров» и содержал окаменелости рогатых динозавров.[6] 20 июня 1986 года Хорнер и Макела вернулись в резервацию индейцев черноногих и возобновили работы на карьере Dinosaur Ridge.[7] который, как оказалось, содержал, помимо яиц, более дюжины скелетов рогатого динозавра, позже названного Эйниозавр. В августе 1986 года на соседнем участке - Костяном ложе каньона на земле Глории Сандквист, к востоку от Молочная река - Команда Хорнера обнаружила еще один Эйниозавр костное ложе. Частью открытий, сделанных по этому поводу, был дополнительный череп рогатого динозавра, образец MOR 492, который позже будет отнесен (то есть официально назначен) Рубеозавр, название рода в 2010 г. Стиракозавр оватус.[8][9]
Во время полевого сезона 1987 года (начало июля) волонтер Сидни М. Хостеттер обнаружил еще один череп рогатого динозавра возле костного ложа каньона, образец MOR 485.[10] К концу августа он был закреплен и перегонялся на зерновоз в Музей Скалистых гор в Бозман.[11] 23 июня 1988 г. поблизости было обнаружено еще одно место - Северный Блэктейл-Крик.[12] Летом 1989 г. аспирант Скотт Д. Сэмпсон присоединился к команде, желая изучить функцию витражей у рогатых динозавров.[13] В конце июня 1989 года Хорнер, его сын Джейсон и его главный специалист по подготовке Кэрри Анселл обнаружили образец рогатого динозавра MOR 591 недалеко от Блэктейл-Крик.[14]
Интерпретация собранных окаменелостей

Первоначально предполагалось, что весь материал рогатых динозавров, обнаруженный экспедициями, можно отнести к одному виду «стиракозавров», отличному от Стиракозавр альбертенз, поскольку окаменелости представляют собой ограниченный геологический период времени, который тогда оценивался примерно в полмиллиона лет.[15] Раймонд Роберт Роджерс, который изучал стратиграфия костного ложа, называемого Стиракозавр sp. (неопределенных видов) в 1989 г.[16] Стиракозавр оватус - хотя иногда считается недействительным номен дубиум[17] - уже был обнаружен в этом районе Г. Ф. Штернбергом и являлся очевидным кандидатом.[15] Но также была принята во внимание возможность того, что находки относятся к новому для науки виду. Этот вид был неофициально назван «Styracosaurus makeli» в честь Боба Макелы, который погиб в дорожно-транспортном происшествии всего за несколько дней до открытия экземпляра MOR 485.[18] В 1990 году это имя, как недействительное. nomen nudum,[19] появляется в подписи к фотографии в книге Стивена Черкаса.[20]
Хорнер, эксперт по Hadrosauridae семья, менее близка к другим видам динозавров.[15] В 1987 и 1989 годах специалист по рогатым динозаврам. Питер Додсон был приглашен исследовать новый цератопсий находит.[15] В 1990 году ископаемый материал рассматривался Додсоном как усиление аргументов в пользу обоснованности отдельного исследования. Стиракозавр оватус, чтобы отличаться от Стиракозавр альбертенз.[21]
Тем временем Хорнер пришел к более сложному взгляду на ситуацию. Он по-прежнему считал, что ископаемый материал был частью одной популяции, но пришел к выводу, что со временем это сформировалось как хроновиды превращаясь в серию последующих таксоны. В 1992 году Хорнер, Дэвид Варриккио и Марк Гудвин опубликовали статью в Природа на основе шестилетних полевых исследований отложений и динозавров в Монтане. Они предположили, что экспедиции обнаружили три "переходные таксоны "преодолевая разрыв между уже известными Стиракозавр и Пахиринозавр. На данный момент они отказались назвать эти таксоны. Самая старая форма была обозначена как «Переходный таксон А», в основном представленная черепом MOR 492. Затем появился «Таксон В» - многочисленные скелеты из карьера хребта динозавров и костного ложа каньона. Самым молодым был «Таксон C», представленный черепом MOR 485 и окаменелостями рогатого динозавра из Блэктейл-Крик.[22][23]
Имена Сэмпсона Ахелозавр

Сэмпсон продолжал изучение материала с 1989 года. В 1994 году во время выступления на ежегодном собрании Общество палеонтологии позвоночных, он назвал «Таксон C» новым родом и видом, Ахелозавр горнери. Хотя Абстрактные был опубликован с достаточным описанием, не идентифицировал голотип, именной экземпляр.[24] В 1995 году в последующей статье Сэмпсон указал на образец MOR 485 как на голотипный образец Ахелозавр горнери. Общее название состоит из слов Ахелой, имя Греческий мифологический фигура и савр, который Латинизированный греческий для ящерицы. Ахелой (Ἀχελῷος) - греческое речное божество и оборотень который смог превратиться во что угодно. Во время боя с Геркулес, мифический герой, Ахелай принял форму быка, но проиграл битву, когда один из его рогов был удален. Этот намек является отсылкой к предположительно переходным чертам динозавра и характерной потере рогов из-за онтогенетический и филогенетический развитие, и, следовательно, через индивидуальные изменения и эволюцию.[22] В 1996 году Додсон похвалил родовое имя за оригинальность и интеллект.[15] Это конкретное имя удостоено Джека Хорнера за его исследования динозавров формации Два лекарства в Монтане. Сэмпсон также назвал род "Таксон B" Эйниозавр в той же статье, где Ахелозавр был описан. Он сказал, что палеонтологи должны быть осторожны при назывании новых родов цератопсов, потому что их внутривидовая изменчивость (то есть вариация внутри вида) может быть ошибочно принята за межвидовые различия (между видами). До 1995 года был назван только один новый род динозавров центрозавринов. Пахиринозавр в 1950 году, а именно Авацератопс в 1986 г.[22] Ахелозавр таким образом, имеет особое значение как один из немногих родов цератопсид, названных в конце двадцатого века.[25]
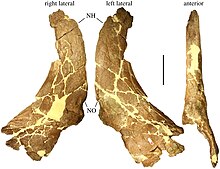
Образец голотипа MOR 485 был собран Хостеттером и Рэем Роджерсом.[26] от области оползня-Батт-Филд примерно в 40 км (25 миль) к северо-западу от Cut Bank. В 1995 году Сэмпсон описал его как частичный череп взрослого животного, включая носовой и надглазничный (область над глазницей) бугорки (вместо рогов округлые выступы) и теменные кости.[22] Кроме того, MOR 485 сохраняет некоторые кости задней и боковых сторон черепа, которые в 2009 году были перечислены Трейси Л. Форд как правильные. чешуйчатая кость, левый чешуйчатый, оба максиллы, обе слезные кости, обе квадратные кости, обе небные кости, то черепная коробка и базиально-затылочная кость.[27] В 2015 году Леонардо Майорино сообщил, что как часть того же образца фрагментарная нижняя челюсть была внесена в каталог как MOR 485-7-12-87-4.[28] Правая чешуйчатая кость другого взрослого человека была извлечена на том же участке костного ложа в каньоне, что и MOR 485 (и занесена в каталог под тем же номером), но сообщена только в 2010 году.[29] Два других экземпляра были собраны в Блэктейл-Крик, в 35 км (22 милях) к югу от Кат-Бэнк, и относились к ним. Ахелозавр Сэмпсона в 1995 году. Образец MOR 591 представляет собой частичный череп и примерно на 60% полный скелет субзрослого образца, который включает позвоночник, таз, крестец и бедренная кость.[22] Сюда также входят нижние челюсти, обозначенные как MOR 591-7-15-89-1.[30] И череп, и нижняя челюсть почти закончены, отсутствуют только черепная коробка и нижняя челюсть. затылочная область.[27] MOR 591 меньше голотипа с длиной основания черепа около 60 см (24 дюйма).[31] Образец MOR 571 включает частичный череп и нижнюю челюсть с соответствующими ребрами и позвонками взрослого человека.[22] Череп состоит только из теменных костей, а нижние челюсти ограничены их верхними задними костями, surangulars и суставы.[27] Пятый экземпляр - MOR 456.1, несовершеннолетний.[32] Ни один из особей не был пожилого индивидуального возраста.[33] По словам Эндрю Макдональда и его коллег, Ахелозавр находки представлены одиночными особями, а не костными пластами.[34]
Возможный Ахелозавр находит

В дополнение к окаменелостям, которые однозначно были отнесены к Ахелозавр, был найден другой материал, идентичность которого не установлена. А центрозаврин цератопсид с выступами из Формирование парка динозавров (экз. TMP 2002.76.1), обнаруженный в 1996 г., в 2006 г. было предложено отнести к новому таксону, но вместо этого может принадлежать к Ахелозавр или же Пахиринозавр. Поскольку в нем отсутствуют теменные кости, которые используются для диагностировать centrosaurines, уверенно отнести его к какому-либо роду невозможно.[33][35] В 2006 г. также было предложено Monoclonius lowei, сомнительный вид, основанный на черепе (образец CMN 8790) из формации парка динозавров, мог быть суб-взрослым экземпляром Стиракозавр, Ахелозавр или же Эйниозавр, с которым он примерно одновременно.[36] Кроме того, есть несколько неопределенных экземпляров из формации Два лекарства, например, фрагментарный череп MOR 464[37] или морда MOR 449 - может принадлежать Ахелозавр или два других примерно современных цератопсида Эйниозавр и Рубеозавр.[8]
Описание
Общая сборка

Ахелозавр По оценкам, его длина составляла 6 м (20 футов), а вес - 3 т (3,3 коротких тонны).[38] Череп взрослого человека (голотип MOR 485), по оценкам, имел длину 1,62 м (5,3 фута). Это помещает его в тот же диапазон размеров, что и другие члены Centrosaurinae подгруппа цератопсы которые жили во время Кампанский возраст. Он был примерно таким же большим, как и его ближайший родственник. Эйниозавр, но с гораздо более тяжелой сборкой.[22] Ахелозавр приблизился к прочности одного из самых крупных и крепко сложенных рогатых динозавров - Трицератопс.[39]
Как цератопсид, Ахелозавр был бы четвероногий животное с копытными пальцами и укороченным, загнутым вниз хвостом. Его очень большая голова, которая могла бы покоиться на прямой шее, имела крючковатый верхний клюв, очень большие носовые отверстия и длинные ряды зубов, превращенные в зубные батареи, которые содержали сотни сжатых и уложенных друг на друга отдельных зубов.[22][38] В лунках зубов новые зубы росли под старыми, в каждой позиции располагался столбец зубов, расположенных друг над другом. Ахелозавр в каждой верхней челюсти (кость верхней челюсти) было от 25 до 28 таких позиций зубов.[40]
Отличительные черты
В 1995 году при описании вида Сэмпсон дал официальный список из четырех черты, которые отличают Ахелозавр от его родственников центрозавринов. Во-первых, у взрослых людей носовые кости с выступом наверху, относительно небольшим и тонким, сильно покрыты ямками; во-вторых, у взрослых особей над глазницами настоящих рогов нет, а есть относительно крупные бобышки с высокими гребнями; в-третьих, у еще не выросших особей или субзрелых есть настоящие роговые ядра (костная часть рогов) над глазницами с обращенной внутрь поверхностью вогнутый; в-четвертых, на теменных костях шейного щита есть одна пара изогнутых шипов, торчащих от заднего края кзади и снаружи.[22]
Помимо этих уникальных характеристик, Сэмпсон указал на дополнительные различия между двумя очень близкими формами. Шипы с оборками Ахелозавр больше ориентированы вовне, чем шипы Эйниозавр, которые изогнуты медиально; шипы Ахелозавр тем не менее, менее направлены вовне, чем сопоставимые шипы Пахиринозавр. Ахелозавр также отличается от Пахиринозавр в его меньшем носовом выступе, который не достигает лобных костей сзади. Кроме черепа, не известны какие-либо особенности скелета, позволяющие различать Ахелозавр от других представителей Centrosaurinae.[22]
Череп

Рогатые динозавры в основном отличаются друг от друга рогами, которые расположены на морде и над глазами, и большой оборкой черепа, которая щитом прикрывает шею. Ахелозавр выставил сборку полученный («продвинутые») центрозаврины, которые представлены короткими надбровными рогами или выступами в сочетании с замысловатыми шипами с оборками. Общие пропорции оборок обычно центрозавриновые, с широкой округлой чешуйчатой костью сбоку, которая расширяется к задней части.[41] Он также имеет типичную кривизну оборки с верхней поверхностью, которая выпуклый сбоку в сторону и вогнутая спереди назад.[42]
Взрослый Ахелозавр черепа имели морщинистый выступ с сильными ямками на морде или в области носа, тогда как у многих других цератопсидов был рог.[22] Такую бобышку часто называют «пахиостотической», т.е. состоящей из утолщенной кости.[43] Но описание его как толстого «выступа» может ввести в заблуждение: на самом деле он образует широкую впадину с тонким костным полом и неровными выемками, хотя и менее вдавлен, чем с Пахиринозавр.[33] Носовая бобышка закрывала примерно две трети верхней поверхности носовых костей.[8] Босс был похож на того, что видели в родственном роду Пахиринозаврхотя и уже, короче и менее высоки.[22] Она покрывала 27% общей длины черепа, была на 30% длиннее, чем расстояние от ноздри до глазницы, и примерно в два раза длиннее, чем глазница.[44] Его задний край не доходил до уровня глазницы.[33] Носовая бобышка выдвигалась вперед, где сливалась с носовой и предчелюстные кости (верхней челюсти) в передней части морды,[22] хотя сама носовая кость не срослась с предчелюстной костью.[33] Бобышка образца MOR 485, кроме того, имела выемку (или полость) на переднем конце. Сердцевина рога, которая образовывала бобышку, могла развиться либо в результате изгиба (т. Е. Изгиба вперед) во время роста, как рог соответствующего рога. Эйниозавр, пока он не срастается с носовой костью; или от простого прямостоячего рога, который позже расширял свое основание вперед над областью морды, как в Пахиринозавр.[22] Носовая кость образовывала вершину большой костной ноздри. От заднего края этой ноздри вперед торчал острый отросток.[8] Морда была - по сравнению с Эйниозавр - относительно широкие на уровне задних ноздрей. Слезная кость перед глазницей была утолщена, в основном на внутренней поверхности, в то время как внешняя поверхность была безликой, если не считать кратерообразной выемки.[45]

У взрослых черепов также есть большие, морщинистые и овальные бобышки на надглазничной области над глазами, вместо рогов других цератопсид. Надглазничные бугорки отходили от заглазничная кость вперед, чтобы включить треугольник пальпебральный и префронтальные кости, и имел высокие поперечные гребни вокруг середины, которые были толстыми у основания и тонкими к вершине.[22] Глазные кости сильно выделялись, образуя «анторбитальный контрфорс». Сросшиеся префронталы не дошли до носовой бобышки,[22] образуют характерную поперечную седловидную канавку, отделяющую носовую бугорку от надглазничных бугорков.[46] Эта бороздка проходила назад, отделяя надглазничные выступы друг от друга и образуя Т-образную форму на виде сверху.[33] Эти боссы были похожи на боссов Пахиринозавр, но с более высокими гребнями и более выраженной шероховатостью. Длинные и низкие надглазничные роговые ядра суб-взрослой особи MOR 591 были аналогичны таковым суб-взрослой особи. Эйниозавр и Пахиринозавр. У них с внутренней стороны была вогнутая поверхность, как у Пахиринозавр; на заглазничных костях присутствовали гребни, которые могут указывать на начало перехода к бугоркам.[22]
Крыша черепа Ахелозавр имел полость по средней линии с отверстием наверху, называемым лобным родничок, особенность, обнаруженная у всех цератопсид, у которых есть «двойная» крыша черепа, образованная лобными костями, складывающимися навстречу друг другу между основаниями надбровных рогов. Эта полость образовала пазухи который проходил ниже надглазничных выступов, которые, следовательно, были относительно тонкими внутри, толщиной 25 мм (1 дюйм) снаружи до крыши полости. Эта полость, по-видимому, частично закрылась у пожилого животного, и только задняя часть родничка была открыта у взрослого экземпляра MOR 485.[22]

Как и у всех других цератопсид, череп Ахелозавр имел теменно-чешуйчатую оборку или «шейный щиток», который образовывался из теменных костей сзади и чешуек по бокам. Теменная кость является одной из основных костей, используемых для различения таксонов центрозавринов друг от друга и установления взаимосвязей между ними, тогда как чешуйчатая кость очень похожа для разных таксонов.[22] В Ахелозавр, чешуйчатая кость была значительно короче теменной. По своему внутреннему краю задняя часть образовывала ступеньку по отношению к передней части, при этом шов между чешуйчатой и теменной костью показывает изгиб сзади на уровне задней части. надвисочное окно, типичная черта центрозаврина.[8] Плоскоклеточная и скуловая кость, касаясь друг друга, исключили четырехъядерный от края бокового височное окно, то есть отверстие в задней части черепа.[47]
Оборка Ахелозавр имел два заметных больших шипа, направленных назад и загнутых в стороны друг от друга. В течение 1990-х годов все больше понималось, что такие шипы на теменных костях - это не случайные образования, а специфические черты, которые можно было бы использовать для определения эволюции рогатых динозавров, если бы только можно было проанализировать, как они соотносятся между видами. Сэмпсон в статье, описывающей Ахелозавр Поэтому в 1995 году была введена обобщенная система нумерации таких теменных отростков, считая их от средней линии до боковой стороны оборки.[22] Это было применено к центрозавринам в целом в 1997 г.[48] Большие шипы Ахелозавр соответствуют шипам «Процесса 3» других центрозавринов и аналогичны шипам у Эйниозавр, хотя и изогнутые в стороны, похожие на Пахиринозавр.[22] Они были короче и тоньше, чем соответствующие шипы Стиракозавр.[49] Между этими шипами, по обе стороны от центральной вырезки, были два небольших язычковых выступа («Процесс 2»), которые были направлены к средней линии.[22] Внутренние пики «Процесса 1», присутствующие в Центрозавр, отсутствуют с Ахелозавр.[49] В оборке было два больших парных отверстия - теменные отверстия, между которыми располагалась срединная теменная перемычка. Линейный ряд округлых вздутий проходил вдоль верхней части теменной перемычки, которая может быть гомологичный к шипам и рогам в той же области некоторых Пахиринозавр образцы. Ряд относительно небольших отростков проходил по краю теменного щита от выступов «отростка 3» наружу,[22] всего на каждую сторону семь. Они были в основном одинакового размера, поэтому процесс P4 был меньше, чем P3.[50] Эти нижние отростки, по-видимому, были закрыты эпокципиталами - костями, выстилающими оборки цератопсид.[22] В Ахелозавр эти эпокципиталы, которые начинаются как отдельная кожа окостенения или же остеодермы, соединяются с подлежащей оборкой, образуя шипы,[51] хотя бы на третьей позиции.[52] В 2020 году отрицалось, что эти процессы были отдельными окостенениями. У наиболее зрелых особей передние отростки P6 и P7 будут менее членистыми относительно друг друга, повернутыми вокруг своих продольных осей.[50]
Кератиновые оболочки

Боссы на черепе Ахелозавр мог быть охвачен ороговевший ножны в жизни, но их форма у живого животного неясна.[22] В 2009 году палеонтолог Тобин Л. Иеронимус и его коллеги изучили корреляции между морфологией черепа, рогом и особенностями кожи современных рогатых животных и исследовали череп центрозавриновых динозавров на предмет тех же коррелятов. Они предложили морщинистым боссам Ахелозавр и Пахиринозавр были покрыты толстыми подушечками ороговевшей (или ороговевшей) кожи, как у босса современных овцебык (Ovibos moschatus). Носовой рог взрослого Ахелозавр имел наклон вверх, а его верхняя поверхность имела корреляты для толстого эпидермальный (внешний слой кожи) подушечка, которая соотносится с ороговевшей оболочкой по бокам. Толстая подушечка эпидермиса могла вырасти из V-образной выемки на кончике рогового ядра носа. Направление роста подушечки носа было бы вперед. Надглазничные бугорки могли иметь толстую подушечку эпидермиса, которая росла под углом, похожим на изогнутые роговые ядра Коронозавр, на что указывает ориентация «плавников» или выступов на выступах. Что надглазничным бугоркам не хватало борозда (или борозда) у их оснований указывает на то, что их подушечки рога останавливались на морщинистых краях выступов. Ямка может указывать на более мягкий растущий слой, соединяющий твердую внутреннюю кость с твердым рогом. Кроме того, корреляты для ростральная шкала перед носовой бугоркой и чешуйчатыми рядами по срединной линии теменной и надглазнично-чешуйчатой области.[53]
Эволюция
Гипотеза Хорнера об анагенезе

В 1992 году исследование Хорнера и другие. пытались объяснить тот факт, что в течение ограниченного геологического периода времени (около полумиллиона лет) произошла быстрая преемственность сообществ животных в верхней формации Два Медицины. Обычно это интерпретируется как серия вторжений, когда новые типы животных заменяют старые. Но Хорнер заметил, что новые формы часто сильно похожи на предыдущие. Это наводило его на мысль, что он открыл редкое доказательство эволюции в действии: более поздний фауна был в основном старым, но на более развитой стадии. Различные обнаруженные типы не были отдельными видами, но переходные формы разработан в процессе анагенез. Это соответствовало распространенному в то время предположению, что вид должен существовать около двух-трех миллионов лет. Еще одним признаком, по словам Хорнера, была неспособность идентифицировать истинное аутапоморфии - уникальные признаки, доказывающие, что таксон является отдельным видом. Вместо этого окаменелости показали постепенное изменение от базальный (или предков) на более производные символы.[23]

Рогатые динозавры, обнаруженные Хорнером, являются примером этого явления. В самых нижних слоях формации Two Medicine, на 60 м (200 футов) ниже перекрывающего слоя. Формация Медвежьей Лапы «Переходный таксон А» присутствовал. Это было похоже на Стиракозавр альбертенз, отличающийся от него лишь наличием одной пары теменных шипов.Средние слои, находящиеся на 45 м (150 футов) ниже Медвежьей лапы, содержали «переходный таксон B», который также имел одну пару шипов, но отличался формой носового рога, который загибался вперед над передними ветвями носовых костей. В верхних слоях, на 20 м (65 футов) ниже Медвежьей Лапы, был раскопан «переходный таксон C». У него тоже была пара шипов, но теперь носовой рог сросся с передними ветвями. Верхняя поверхность рога была приподнятой и очень шероховатой. На орбитальных рогах видны грубые гребни. Впоследствии «Таксон А» был назван Стеллазавр,[9][50] «Таксон B» стал Эйниозавр, а "Таксон C" стал Ахелозавр.[23] В 1992 году Хорнер и другие. не назвал их видами по той простой причине, что вся эволюционная последовательность рассматривалась как представляющая ступень переходных цератопсов между Стиракозавр альбертенз, известный из Формация реки Джудит, а производные, безрогие Пахиринозавр от Формация каньона Подкова с парой шипов и бобышками на носу и над глазами, а также с дополнительным орнаментом из оборок.[23] В 1997 году Хорнер назвал эти три таксона «центрозаврином 1», «центрозаврином 2» и «центрозаврином 3 ..».[54]
Хорнер думал, что он нашел механизм, приводящий в движение эту эволюцию, развивая идеи, которые он разработал еще до того, как исследовал Landslide Butte.[55] Животные жили на узкой полосе на восточном побережье Ларамидии, граничащей с Западный внутренний морской путь и ограничена на западе высотой от 3 до 4 километров (от 2 до 2,5 миль). прото-Скалистые горы. Во время Трансгрессии Медвежьей Лапы уровень моря повышался, неуклонно сокращая ширину их прибрежной среды обитания примерно с 300 км (200 миль) до 30 км (20 миль).[56] Это привело к усилению давление отбора,[23] самый суровый для Ахелозавр которые жили в ту фазу, когда прибрежная полоса была самой узкой.[57] Меньшее количество особей, которое могла выдержать меньшая среда обитания, составляло узкое место населения, делая возможной быструю эволюцию.[15] Повысился половой отбор вызвало бы изменения в половом орнаменте, таком как шипы, рога и выпуклости.[23] С другой стороны, снижение экологического стресса из-за более низкого уровня моря будет характеризоваться адаптивное излучение. То, что половой отбор действительно был основным механизмом, будет доказано тем фактом, что молодые особи всех трех популяций были очень похожи: все они имели два шипа-оборки, небольшой носовой рог, указывающий вперед, и орбитальные рога в форме слегка приподнятых шипов. повышенные ручки. Только во взрослой фазе они стали отличаться. По словам Хорнера, это также показало, что популяции были очень тесно связаны.[58]

Хорнер не выполнил точного кладистический анализ определение отношений между тремя популяциями. Такой анализ вычисляет, какое эволюционное дерево подразумевает наименьшее количество эволюционных изменений и, следовательно, является наиболее вероятным. Он предположил, что это приведет к дереву, в котором типы будут последовательными ветвями. Такое дерево, как следствие используемого метода, никогда не будет показывать прямую связь предок-потомок. Многие ученые считали, что такую связь невозможно доказать. Хорнер не согласился: он рассматривал постепенные морфологические изменения как явное доказательство того, что в этом случае эволюцию одного таксона в другой без разделения популяций можно наблюдать непосредственно. Эволюционисты в целом не решились бы это признать.[59] Такой переход называется анагенезом; он утверждал, что в противном случае кладогенез, не могло быть доказано, ученый был свободен предположить анагенетический процесс.[23]
Основываясь на уточненных данных, Сэмпсон в 1995 году подсчитал, что исследованные слои представляли собой более длительный период времени, чем первоначально предполагалось 500000 лет: после осаждения Гилмора. Брахицератопс карьера, прошло бы 860 000 лет, а после Эйниозавр кровати 640 000 лет, до максимальной степени трансгрессии Bearpaw. Он не принял гипотезу Хорнера об анагенезе, но предположил, что видообразование произошел с разделением популяций. Эти временные интервалы все еще были достаточно короткими, чтобы указывать на то, что скорость видообразования должна была быть высокой, что могло быть верно для всех центрозавринов позднего кампана.[22]

В 1996 году Додсон выдвинул два возражения против гипотезы Хорнера. Во-первых, наличие только одной пары главных шипов казалось более базальным, чем наличие трех пар, как в случае с Стиракозавр альбертенз. Это подсказало ему, что Эйниозавр–Ахелозавр родословная была отдельной ветвью в пределах Centrosaurinae. Во-вторых, он был обеспокоен тем, что Эйниозавр и Ахелозавр были случай половой диморфизм, один тип - мужчины, другой - женщины. Об этом может свидетельствовать короткий геологический интервал времени между слоями, в которых были обнаружены их окаменелости, который, по его оценке, составлял около 250 000 лет. Но если бы гипотеза верна, это был бы, пожалуй, лучший пример быстрой эволюции динозавров.[15]
В 2010 году Хорнер признал, что образец TMP 2002.76.1, казалось, указывает на то, что Ахелозавр не был потомком Эйниозавр, так как он предшествовал обоим по возрасту, но при этом имел носовой выступ. Но он подчеркнул, что даже если бы линии разделились, его предок мог походить на Эйниозавр. Более того, возможно, что Эйниозавр был прямым потомком Рубеозавр. Кроме того, по его мнению, процесс быстрого перемещения и вымирания видов все еще можно элегантно объяснить расширением моря Медвежьей Лапы на запад.[8]
Процесс анагенеза был подтвержден Джоном Уилсоном и Джеком Сканнеллой в 2016 году, которые изучали онтогенетические изменения у рогатых динозавров. Они сравнили небольшой Эйниозавр образец, MOR 456 8-8-87-1, с Ахелозавр образец MOR 591. Оба оказались очень похожими, с основными различиями в более длинном лице в MOR 456 8-8-87-1 и более остром надглазничном роговом ядре в MOR 591. Они пришли к выводу, что Ахелозавр вероятно был прямым потомком Эйниозавр. Более взрослый Эйниозавр люди обратились к Ахелозавр морфология. Различия между двумя таксонами были бы вызваны гетерохрония - дифференциальные изменения в скорости различных черт, развившихся в течение жизни человека.[31] Поскольку в 2020 году Уилсон и его коллеги обнаружили, что Стеллазавр («Таксон А» Хорнера) занимал промежуточное положение между Стиракозавр и Эйниозавр в морфологии и стратиграфии они не могли сбрасывать со счетов, что это был переходный таксон в пределах анагенетической линии.[50]
Классификация
В 1995 году Сэмпсон официально разместил Ахелозавр в Ceratopsidae, точнее Centrosaurinae.[22] Во всех анализах Эйниозавр и Ахелозавр являются частью клады Pachyrhinosaurini. По определению, Ахелозавр является членом клада Пахиростра (или «толстые рыла»), в которых он соединен с Пахиринозавр.[60] В 2010, Грегори С. Пол назначенный A. horneri к роду Центрозавр, так как C. horneri.[38] Это не нашло признания среди других исследователей, с последующим таксономический оценки, неизменно сохраняя родовое название Ахелозавр.[61][62][60][63]
Филогения

В 1995 г. Сэмпсон считал, что недостаточно доказательств, чтобы сделать вывод о том, что Ахелозавр был прямым потомком Эйниозавр. В отличие от Хорнера, он решил провести кладистический анализ, чтобы установить филогения. Это показало эволюционное древо, в котором Ахелозавр разделены между Эйниозавр и Пахиринозавр, как и предсказывал Хорнер. Вопреки утверждению Хорнера, Стиракозавр альбертенз не мог быть прямым предком, так как это был родственные виды из Центрозавр в анализе Сэмпсона.[22]
Последующие исследования были направлены на определение точных взаимоотношений в этой части эволюционного древа, с противоречивыми результатами относительно вопроса о том, является ли Стиракозавр альбертенз или же Эйниозавр мог быть на прямой линии восхождения к Ахелозавр. В 2005 году анализ Майкла Райана и Энтони Рассела показал, что Стиракозавр более тесно связанный с Ахелозавр чем Центрозавр.[64] Это подтвердили анализы, проведенные Райаном в 2007 году.[65] Николас Лонгрич в 2010 году,[66] и Xu et al. в 2010.[67] В том же году Хорнер и Эндрю Т. Макдональд переехали Стиракозавр оватус к собственному роду, Рубеозавр, найдя его родственным видом Эйниозавр, пока Стиракозавр альбертенз снова был расположен на Центрозавр ответвляться. Они также отнесли образец MOR 492, основание «Таксона А», к Рубеозавр.[8] В 2011 году последующее исследование Эндрю Т. Макдональда в этом отношении повторило результаты его предыдущего исследования:[61] как и публикация Андре Фарке и др.[62] В 2017 году Дж. П. Уилсон и Райан еще больше усложнили проблему, заключив, что MOR 492 («Таксон A») не подлежит отнесению к Рубеозавр и объявив, что в его честь будет назван еще один род.[9] Уилсон и его коллеги перевели MOR 492 в новый род Стеллазавр в 2020 году, что, следовательно, соответствует «Таксону А». Их исследование показало Рубеозавр оватус быть родственным видом Стиракозавр альбертенз, и заключил Рубеозавр быть синонимом Стиракозавр.[50]
Перед Ахелозавр был описан, Пахиринозавр канадский считался одиноким отклоняющийся от нормы образуют среди центрозавринов, выделяются своими необычными боссами. Ахелозавр дали эволюционный контекст канадским видам, одновременно расширив временной и географический диапазон того, что стало считаться «пахиринозаврами».[60] Во всех анализах Ахелозавр и Пахиринозавр мы сестринские группы. В 2008 году был назван еще один близкородственный вид: Пахиринозавр lakustai. В этом исследовании термин «пахиринозавры» использовался для обозначения клады, состоящей из Ахелозавр и Пахиринозавр.[68] Когда Пахиринозавр perotorum был описан в 2012 г., было придумано название клады Pachyrostra, объединяющее два рода; Ахелозавр самый базальный пахиростран. Общие производные черты (или синапоморфии ) группы - это увеличенный носовой орнамент и превращение носовых и надбровных рогов в выступы.[60] В конце кампана, по-видимому, наблюдалась тенденция замены пахиространов другими центрозавринами.[33] Также в 2012 году была названа клада Pachyrhinosaurini, состоящая из видов, более близких к Пахиринозавр или же Ахелозавр чем Центрозавр. Помимо Эйниозавр и Рубеозавр, это включало Синоцератопс и Ксеноцератопс, согласно исследованию 2013 года.[69]
Кладистический анализ развивается постепенно, отражая новые открытия и идеи. Их результаты можно показать в кладограмма, с найденными отношениями, упорядоченными в эволюционном дереве. На приведенной ниже кладограмме показано филогенетическое положение Ахелозавр на кладограмме Уилсона и его коллег, 2020.[50]

| Centrosaurinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
Функция орнамента черепа

В 1995 году Сэмпсон отметил, что более ранние исследования показали, что рога и оборки цератопсов, скорее всего, выполняли функцию внутривидовой дисплей и бой, и что эти особенности, следовательно, были результатом полового отбора для успешного спаривания.[22] Точно так же в 1997 году Хорнер пришел к выводу, что такое украшение использовалось самцами для установления доминирования, и что самки предпочли бы хорошо экипированных самцов, поскольку их потомство унаследовало бы эти черты, что дало возможность воспроизводить потомство.[57] Додсон считал, что у Centrosaurinae в целом визуальная ценность оборки была уменьшена по сравнению с носовой и надглазничной орнаментами.[70] Сэмпсон в 1995 году отверг возможность того, что различие в орнаменте черепа между Эйниозавр и Ахелозавр представляет половой диморфизм по трем причинам. Во-первых, обширная Эйниозавр костные пласты не содержали особей с выступами, как можно было бы ожидать, если бы они были у одного из полов. Во-вторых, Эйниозавр и Ахелозавр встречаются в пластах разного возраста. В-третьих, в ситуации полового диморфизма обычно только один из полов показывает преувеличенно второстепенные сексуальные персонажи. Эйниозавр и Ахелозавр однако у каждого из них развился отдельный набор таких черт.[22]
Иероним в 2009 году пришел к выводу, что носовые и надглазничные бобышки использовались для ударов или таранов в голову или бок соперника. Строение костей указывает на то, что боссы были покрыты ороговевшими подушечками, как у современных овцебыков, что предполагает борьбу за доминирование, аналогичную битвам представителей отряда. Caprinae подсемейство. В последней группе можно наблюдать эволюционный переход, когда изначально прямые рога становятся более прочными, мягкими и все более изогнутыми вниз. Эволюция роговых коров в боссов у Centrosaurinae также отразила бы изменение техники боя - от столкновения до высокоэнергетического удара головой. Удар головой был бы дорогим и рискованным поведением. Противники вступили бы в бой только после визуальной оценки сильных сторон друг друга. По этой причине Иероним считал маловероятным, что боссы служили для признание видов поскольку это уже было гарантировано врожденными видоспецифичными ритуалами демонстрации, предшествующими настоящей - а не ритуальной - битве. Боссы эволюционировали для реальных боевых действий, как часть социальный отбор в котором люди конкурировали за ограниченные ресурсы, такие как партнеров, еду и нерестилища.[53]

Ранее предполагалось, что слияние первых трех шейных позвонков, такое как у зрелого экземпляра MOR 571, могло быть палеопатология, случай болезни спондилоартропатия, но в 1997 году был сделан вывод, что это, скорее, нормальный онтогенетический признак, позвонки срослись, образуя так называемый «синервикальный» позвонок, поддерживающий тяжелую голову.[71] Все три основных известных экземпляра имеют синцервикалы, состоящие из трех сросшихся шейных позвонков;[72][73] эта черта могла быть унаследована от более мелкого предка, который использовал более жесткую шею для рытья нор или поиска пищи.[74]
Социальное поведение
Было заявлено, что динозавры цератопсов пасли животных из-за большого количества известных костных пластов, содержащих несколько представителей одного и того же вида цератопсов. В 2010 году Хант и Фарке указали, что это в основном верно для центрозавриновых цератопсов.[29] Хорнер предположил, что рогатые динозавры в Лэндслайд-Бьютте жили стадами, погибшими от засухи или болезней.[75] Додсон пришел к выводу, что тот факт, что Ахелозавр костные ложа были моноспецифичными (содержали только один вид), что подтверждало существование стад.[70]
Метаболизм
Уже давно ведутся споры о терморегуляция динозавров, сосредоточенных вокруг того, были ли они эктотермия («хладнокровный») или эндотермы («теплокровный»). Млекопитающие и птицы теплый эндотермы, которые генерируют тепло собственного тела и имеют высокий метаболизм, а рептилии гетеротермический эктотермы, которые получают большую часть тепла своего тела из окружающей среды. Исследование 1996 года изучило изотопы кислорода из кости фосфаты животных из формации Two Medicine, включая молодь Ахелозавр образец MOR 591. δ18О значения фосфата в костях позвоночных зависят от δ18Значения O в воде их тела и температура, когда кости откладывались, что позволяет измерять колебания температуры для каждой кости человека, когда они откладывались. В исследовании анализировались сезонные колебания температуры тела и различия в температуре между областями скелета, чтобы определить, поддерживают ли динозавры свою температуру сезонно. А варанид Ископаемые останки ящериц, отобранные для исследования, показали изотопные вариации, соответствующие гетеротермическому эктотерму. Вариация динозавров, в том числе Ахелозавр, согласуется с тем, что они являются гомеотермическими эндотермами. Скорость обмена веществ у этих динозавров, вероятно, была не такой высокой, как у современных млекопитающих и птиц, и, возможно, они были промежуточными эндотермами.[76]
Палеосреда

Ахелозавр известен из формации Ту-Медицина, в которой сохранились прибрежные отложения, датируемые кампанским этапом позднего периода. Меловой период, между 83 и 74 миллионами лет назад. Ахелозавр образцы обнаружены на самых высоких уровнях формации, вероятно, ближе к концу этого периода времени, 74 миллиона лет назад.[33]
Формация Two Medicine отличается теплой полузасушливый климат. Его слои располагались на восточном побережье островного материка Ларамидия (который состоял из западной части Северной Америки). Высота Кордильеры на западе, в сочетании с преимущественно западными ветрами, вызвали бы тень дождя, ограничение годового количества осадков. Дождь в основном шел летом, когда конвекционные бури затопили пейзаж. Таким образом, климат также был бы очень сезонным с длинным засушливым сезоном и коротким влажным сезоном. Растительность была бы редкой и мало разнообразной. В таких условиях рогатые динозавры зависели бы от старицы для непрерывного снабжения водой и пищей - основные речные русла имеют тенденцию пересыхать раньше - и погибали в них во время сильной засухи, когда животные концентрировались вокруг последних водопоев, вызывая костные ложа формировать.[77] Коричневый палеопочва в котором были найдены рогатые динозавры - смесь глины и углекислый обломки дерева - напоминают современные сезонно сухие болота. Окружающая растительность могла состоять из хвойных деревьев высотой около 25 м (80 футов).[78] Ахелозавр Однако ели растения гораздо меньшего размера: исследование 2013 года показало, что цератопсидные травоядные на Laramidia ограничены питанием растительностью высотой 1 м (3,5 фута) или ниже.[79]
Включены более или менее современные роды динозавров этого района. Прозавролоф, Сколозавр, Гипакрозавр, Эйниозавр и тираннозавриды неопределенной классификации. Как доказывают следы зубов, окаменелости рогатых динозавров в районе оползня-Бьютт-Филд были обнаружены крупными теропод хищник, который, по мнению Роджерса, был Альбертозавр.[80]

Точный состав фауны Ахелозавр входил в состав неизвестно, так как его окаменелости не были обнаружены в прямой связи с другими таксонами. Его промежуточное анагенетическое положение предполагает, что Ахелозавр разделял его среду обитания с формами, находящимися примерно в середине или в конце временного диапазона своего формирования. Как и в случае с рогатыми динозаврами, Хорнер предположил, что он обнаружил переходные таксоны в других группах динозавров формации Два Медицины. Один из них был формой между Ламбеозавр и Гипакрозавр;[23] в 1994 году он назвал бы это Гипакрозавр stebingeri.[81] Сегодня, Гипакрозавр stebingeri больше не рассматривается как эволюционировавший в результате анагенеза, потому что были идентифицированы аутапоморфии видов.[82] Хорнер видел некоторые пахицефалозавр черепа как показатель таксона между Стегоцеры и Пахицефалозавр;[23] их не всегда относили к новому роду. Наконец, Хорнер подумал, что существует таксон, который является переходным между Дасплетозавр и Тиранозавр.[23] В 2017 году останки тираннозавридов из формации Два лекарства были названы новым видом Дасплетозавр: Дасплетозавр горнери.[83] Исследование 2017 г. посчитало вероятным, что D. horneri был прямым потомком D. torosus в процессе анагенеза, но отвергает возможность того, что D. horneri был предком Тиранозавр.[84]
Другие цератопсы из формации двух лекарств включают Эйниозавр и Стеллазавр. Кроме того, остатки других неопределенных и сомнительных центрозавринов, в том числе Брахицератопс, известны по формированию, и хотя они могут представлять более молодые стадии трех допустимых родов, это невозможно продемонстрировать.[43][50] Тогда как Хорнер предположил, что Эйниозавр и Ахелозавр были разделены во времени, в 2010 году Дональд М. Хендерсон считал возможным, что по крайней мере их потомки или предки пересекались или сочувствующий и, таким образом, соревновались бы за источники пищи, если бы не было перегородка ниши. Череп Ахелозавр был более чем в два раза сильнее, чем у Эйниозавр в его прочности на изгиб и сопротивление кручению. Это могло указывать на разницу в диете, чтобы избежать конкуренции. Сила укуса Ахелозавризмеряется как предел прочности на растяжение, было 30,5ньютоны на квадратный миллиметр (Н / мм²) в ряду зубов верхней челюсти и 18 Н / мм² в клюве.[85] Уилсон и его коллеги обнаружили, что, поскольку центрозаврины Two Medicine были разделены стратиграфически, они, возможно, не были одновременными.[50]
Неопределенный экземпляр TMP 2002.76.1 происходит из формации парка динозавров и, если он принадлежит Ахелозавр, род будет самым старым стратиграфически известным таксоном пахиринозавров.[33] Ахелозавр тогда также будет единственным кампанским цератопсидом, известным из более чем одной формации. Оба животных встречаются прямо под морской сланцы формации Bearpaw, но из-за продольных различий, TMP 2002.76.1 примерно на 500000 лет старше, чем Ахелозавр окаменелости из формации Two Medicine.[35]
Смотрите также
Рекомендации
- ^ Крейслер, Б. (1999). "Бейпиаозавр и Каудиптерикс произношение". Список рассылки динозавров (Список рассылки). Получено 26 сентября, 2017.
Это должно произноситься как ак-э-ЛО-э-э-САВР-нас, а НЕ a-KEE-lo-SAWR-us, насколько я помню. Название происходит от греческого Akheloos (два o произносятся отдельно), который по латинским правилам превращается в Achelous (разделенный на a-che-lo-us, короткое e, долгое o и краткое u), снова произносимый в четырех слогах с ударением. на предпоследнем, это тот, у которого есть длинное o. Все источники, которые я проверил, указывают, что ak-e-LOH-us - это общепринятое английское произношение латинского имени. Поскольку научное название Achelousaurus было образовано путем произвольного сочетания Achelous и Saurus вместо использования формы основы Achelo-, букву «u» необходимо произносить.
- ^ Хорнер и Добб, 1997 г. С. 53–75.
- ^ Хорнер, Дж. Р .; Горман, Дж. (1988). Копание динозавров: поиски, которые раскрыли тайну маленьких динозавров. Нью-Йорк: Уоркман Паблишинг Ко. Стр.210. ISBN 978-0-06-097314-8.
- ^ Хорнер и Добб 1997 С. 60–61.
- ^ Гилмор, C.W. (1930). «О динозаврах-рептилиях из двух лекарств Монтаны». Труды Национального музея США. 16. 77 (2839): 1–39. Дои:10.5479 / si.00963801.77-2839.1.
- ^ Хорнер и Добб, 1997 г., п. 64.
- ^ Хорнер и Добб, 1997 г. С. 65–66.
- ^ а б c d е ж грамм Макдональд и Хорнер 2010 С. 156–168.
- ^ а б c Wilson, J.P .; Ryan, M.P .; Эванс, округ Колумбия (23–26 августа 2017 г.). «Новый центрозаврин цератопсид из двух медицинских формаций верхнего мела Монтаны и эволюция динозавров« Стиракозавр »» (PDF). In Farke, A .; MacKenzie, A .; Миллер-Кэмп, Дж. (Ред.). Тезисы докладов. Общество палеонтологии позвоночных: 77-е ежегодное собрание. Калгари, AB, Канада. п. 214.
- ^ Хорнер и Добб 1997, п. 82.
- ^ Хорнер и Добб 1997, п. 84.
- ^ Хорнер и Добб, 1997 г., п. 96.
- ^ Хорнер и Добб 1997, п. 103.
- ^ Хорнер и Добб 1997, п. 104.
- ^ а б c d е ж грамм Додсон, П. (1996). Рогатые динозавры: естественная история. Принстон, Нью-Джерси: Издательство Принстонского университета. стр.193–197. ISBN 978-0-691-62895-0.
- ^ Роджерс 1989, п. 49.
- ^ Додсон, Форстер и Сэмпсон 2004, п. 496.
- ^ Хорнер и Добб, 1997 г. С. 80–81.
- ^ Райан, Холмс и Рассел 2007, п. 944.
- ^ Czerkas, S.J .; Черкас, С.А. (1990). Динозавры: взгляд со всего мира. Лимпсфилд: мир драконов. п.208. ISBN 978-0-7924-5606-3.
- ^ Dodson, P .; Карри, П.Дж. (1990). «Неоцератопсия». In Weishampel, D.B .; Dodson, P .; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Калифорнийский университет Press. С. 593–618. ISBN 978-0-520-25408-4.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление Сэмпсон, С. (1995). «Два новых рогатых динозавра из формации Две Медицины верхнего мела Монтаны; с филогенетическим анализом Centrosaurinae (Ornithischia: Ceratopsidae)». Журнал палеонтологии позвоночных. 15 (4): 743–760. Дои:10.1080/02724634.1995.10011259.
- ^ а б c d е ж грамм час я j Хорнер, J.R .; Varricchio, D.J .; Гудвин, М. (1992). «Морские нарушения и эволюция меловых динозавров». Природа. 358 (6381): 59–61. Bibcode:1992Натура 358 ... 59H. Дои:10.1038 / 358059a0. S2CID 4283438.
- ^ Сэмпсон, С. Д. (1994). «Два новых рогатых динозавра (Ornithischia: Ceratopsidae) из формации Two Medicine в верхнемеловом периоде, Монтана, США». Журнал палеонтологии позвоночных. 14 (Приложение 3): 44A. Дои:10.1080/02724634.1994.10011592.
- ^ Найш, Д. (2009). Великие открытия динозавров. Лондон: A&C Black Publishers LTD. С. 144–145. ISBN 978-1-4081-1906-8.
- ^ Хорнер, Дж. Р. (2001). Динозавры под большим небом. Издательская компания Mountain Press. п. 109. ISBN 978-0-87842-445-0.
- ^ а б c Форд 2010, п. 53.
- ^ Майорино 2015, п. 210.
- ^ а б Хант и Фарке 2010 С. 447–455.
- ^ Майорино 2015, п. 334.
- ^ а б Wilson, J.P .; Сканнелла, Дж. Б. (2016). "Сравнительная черепная остеология недозревших динозавров центрозавринов из формации Два Медицины, Монтана". Журнал палеонтологии позвоночных, программа SVP и сборник тезисов, 2016: 252.
- ^ Майорино 2015, п. 365.
- ^ а б c d е ж грамм час я Райан и др. 2010 г. С. 141–155.
- ^ Макдональдс и Фарке 2011, п. 1.
- ^ а б Сэмпсон и Лёвен 2010 С. 405–427.
- ^ Райан, М.Дж. (2006). «Статус проблемного таксона Моноклониус (Ornithischia: Ceratopsidae) и признание таксонов динозавров взрослого размера ». Рефераты Геологического общества Америки с программами. 38 (4): 62.
- ^ Макдональдс и Фарке 2011, п. 11.
- ^ а б c Пол, Г.С. (2010). Принстонский полевой путеводитель по динозаврам. Издательство Принстонского университета. стр.249, 257, 263. ISBN 978-0-691-13720-9.
- ^ Карри, Лэнгстон и Танке, 2008 г., п. 17.
- ^ Карри, Лэнгстон и Танке, 2008 г., п. 25.
- ^ Майорино 2015, п. 139.
- ^ Sampson, S.D .; Lund, E.K .; Loewen, M.A .; Farke, A.A .; Клейтон, К. (2013). «Замечательный короткорогий динозавр из позднего мела (поздний кампан) на юге Ларамидии». Труды Королевского общества B: биологические науки. 280 (1766): 20131186. Дои:10.1098 / rspb.2013.1186. ЧВК 3730592. PMID 23864598.
- ^ а б Макдональдс и Фарке 2011, п. 3.
- ^ Карри, Лэнгстон и Танке, 2008 г., п. 29.
- ^ Карри, Лэнгстон и Танке, 2008 г., п. 36.
- ^ Карри, Лэнгстон и Танке, 2008 г., п. 41.
- ^ Карри, Лэнгстон и Танке, 2008 г., п. 50.
- ^ Sampson, S.D .; Райан, MJ; Танке, Д. Х. (1997). «Черепно-лицевой онтогенез у центрозавриновых динозавров (Ornithischia: Ceratopsidae): таксономические и поведенческие последствия». Зоологический журнал Линнеевского общества. 121 (3): 293–337. Дои:10.1111 / j.1096-3642.1997.tb00340.x.
- ^ а б Райан, Холмс и Рассел 2007, п. 951.
- ^ а б c d е ж грамм час Уилсон, Джон П .; Райан, Майкл Дж .; Эванс, Дэвид С. (2020). "Новый переходный центрозавриновый цератопсид из двухмедицинской формации верхнего мела штата Монтана и эволюция 'Стиракозавр-строка «динозавры». Королевское общество открытой науки. 7 (4): 200284. Bibcode:2020RSOS .... 700284W. Дои:10.1098 / rsos.200284. ЧВК 7211873. PMID 32431910.
- ^ Vickaryous, M.K .; Райан, М.Дж. (1997). «Орнамент». В Currie, P.J .; Падиан, К. (ред.). Энциклопедия динозавров. Сан-Диего: Academic Press. стр.488 –493. ISBN 978-0-12-226810-6.
- ^ Макдональдс и Фарке 2011, п. 6.
- ^ а б Иероним, Т.Л .; Witmer, L.M .; Tanke, D.H .; Карри, П.Дж. (2009). «Лицевой покров центрозавриновых цератопсидов: морфологические и гистологические корреляты новых структур кожи». Анатомические рекорды: достижения интегративной анатомии и эволюционной биологии. 292 (9): 1370–1396. Дои:10.1002 / ar.20985. PMID 19711467.
- ^ Хорнер и Добб, 1997 г., п. 195.
- ^ Хорнер, Дж. Р. (1984). «Три экологически различных сообщества фауны позвоночных из формации Ту-Медицина в штате Монтана, с обсуждением эволюционного давления, вызванного колебаниями внутренних морских путей». Руководство по полевой конференции и симпозиуму Геологического общества Монтаны 1984 г.: 299–303.
- ^ Хорнер и Добб, 1997 г., п. 193.
- ^ а б Хорнер и Добб, 1997 г., п. 198.
- ^ Хорнер и Добб, 1997 г., п. 196.
- ^ Хорнер и Добб, 1997 г. С. 196–197.
- ^ а б c d Fiorillo, A.R .; Тыкоски, Р. (2012). "Новый маастрихтский вид центрозаврина цератопсид. Пахиринозавр с северного склона Аляски ». Acta Palaeontologica Polonica. 57 (3): 561–573. Дои:10.4202 / app.2011.0033.
- ^ а б Макдональдс и Фарке 2011, п. 9.
- ^ а б Farke, A.A .; Райан, MJ; Barrett, P.M .; Tanke, D.H .; Braman, D.R .; Loewen, M.A .; Грэм, М.Р. (2011). «Новый центрозаврин из позднего мела в провинции Альберта, Канада, и эволюция теменного орнамента у рогатых динозавров». Acta Palaeontologica Polonica. 56 (4): 691–702. Дои:10.4202 / app.2010.0121.
- ^ Райан, MJ; Holmes, R .; Mallon, J .; Loewen, M .; Эванс, округ Колумбия (2017). «Базальный цератопсид (Centrosaurinae: Nasutoceratopsini) из формации Олдман (кампан) в Альберте, Канада». Канадский журнал наук о Земле. 54 (1): 1–14. Bibcode:2017CaJES..54 .... 1R. Дои:10.1139 / cjes-2016-0110.
- ^ Райан, Дж .; Рассел, А.П. (2005). «Новый центрозаврин цератопсид из формации Олдман в Альберте и его значение для таксономии и систематики центрозаврина» (PDF). Канадский журнал наук о Земле. 42 (7): 1369–1387. Bibcode:2005CaJES..42.1369R. Дои:10.1139 / e05-029. HDL:1880/47001.
- ^ Райан, М.Дж. (2007). «Новый базальный центрозаврин цератопсид из формации Олдман, юго-восточная Альберта». Журнал палеонтологии. 81 (2): 376–396. Дои:10.1666 / 0022-3360 (2007) 81 [376: ANBCCF] 2.0.CO; 2. JSTOR 4133785.
- ^ Лонгрич, Н. (2016). "Мохоцератопс перифания, новый цератопсид хазмозаврина из позднего кампана Западной Канады ". Журнал палеонтологии. 84 (4): 681–694. Дои:10.1017 / S002233600005839X.
- ^ Сюй, X .; Wang, K .; Чжао, X .; Ли, Д. (2010). «Первый динозавр цератопсид из Китая и его биогеографические последствия». Китайский научный бюллетень. 55 (16): 1631–1635. Bibcode:2010ЧСБУ..55.1631Х. Дои:10.1007 / s11434-009-3614-5. S2CID 128972108.
- ^ Карри, Лэнгстон и Танке, 2008 г., п. 16.
- ^ Sampson, S.D .; Lund, E.K .; Loewen, M.A .; Farke, A.A .; Клейтон, К. (2013). «Замечательный короткорогий динозавр из позднего мела (поздний кампан) на юге Ларамидии». Труды Королевского общества B: биологические науки. 280 (1766): 20131186. Дои:10.1098 / rspb.2013.1186. ISSN 0962-8452. ЧВК 3730592. PMID 23864598.
- ^ а б Додсон, П. (1997). «Неоцератопия». В Currie, P. J .; Падиан, К. (ред.). Энциклопедия динозавров. Сан-Диего: Academic Press. стр.473 –478. ISBN 978-0-12-226810-6.
- ^ Ротшильд, Б. (1997). «Палеопатология динозавров». In Farlow, J.O .; Бретт-Сурман, М. (ред.). Полный динозавр. Блумингтон и Индианаполис: Издательство Индианского университета. стр.441–442. ISBN 978-0-253-35701-4.
- ^ VanBuren 2013, п. 53.
- ^ VanBuren 2013, п. 100.
- ^ VanBuren 2013, п. 80–84.
- ^ Хорнер и Добб, 1997 г., п. 66.
- ^ Barrick, R.E .; Душевые кабины, W.J .; Фишер, А.Г. (1996). «Сравнение терморегуляции четырех орнитисхийских динозавров и варанидной ящерицы из формации« Две медицины мелового периода »: данные по изотопам кислорода». ПАЛАИ. 11 (4): 295–305. Bibcode:1996Палай..11..295Б. Дои:10.2307/3515240. JSTOR 3515240.
- ^ Роджерс 1989, п. 68–71.
- ^ Retallack, G.J. (1997). Вольберг, Д. (ред.). «Динозавры и грязь». Динофест Интернэшнл: Материалы симпозиума, спонсируемого Государственным университетом Аризоны, Академия естественных наук: 345–359.
- ^ Mallon, J.C .; Evans, D.C .; Райан, MJ; Андерсон, Дж. (2013). «Стратификация по высоте кормления среди травоядных динозавров из формации динозавров (верхний кампан) в Альберте, Канада». BMC Ecology. 13 (1): 14. Дои:10.1186/1472-6785-13-14. ЧВК 3637170. PMID 23557203.
- ^ Роджерс, Р.Р. (1990). «Тафономия трех слоев костей динозавров в верхнемеловой формации Two Medicine на северо-западе Монтаны: данные о смертности, связанной с засухой». ПАЛАИ. 5 (5): 394–413. Bibcode:1990 Палай ... 5..394R. Дои:10.2307/3514834. JSTOR 3514834.
- ^ Хорнер, J.R .; Карри, П.Дж. (1994). «Эмбриональная и неонатальная морфология и онтогенез нового вида Гипакрозавр (Ornithischia, Lambeosauridae) из Монтаны и Альберты ". In Carpenter, K .; Hirsch, K.F .; Horner, J.R. (eds.). Яйца динозавров и младенцы. Кембридж: Издательство Кембриджского университета. С. 312–336. ISBN 978-0-521-56723-7.
- ^ Brink et al. 2015 г. С. 245–267.
- ^ Carr et al. 2017 г., п. 3.
- ^ Carr et al. 2017 г., п. 7.
- ^ Хендерсон 2010 С. 293–307.
Библиография
- Бринк, К. С .; Зеленицкий, Д.К .; Evans, D.C .; Хорнер, Дж. Р .; Терриен, Ф. (2015), "Морфология черепа и вариации в Гипакрозавр stebingeri (Ornithischia: Hadrosauridae) ", в Eberth, D.A .; Evans, D.C. (ред.), Гадрозавры, Издательство Индианского университета, ISBN 978-0-253-01385-9
- Карр, Томас Д .; Варриккио, Дэвид Дж .; Sedlmayr, Jayc C .; Робертс, Эрик М .; Мур, Джейсон Р. (2017), «Новый тираннозавр с доказательствами анагенеза и крокодилоподобной лицевой сенсорной системой», Научные отчеты, 7: 44942, Bibcode:2017НатСР ... 744942C, Дои:10.1038 / srep44942, ЧВК 5372470, PMID 28358353
- Карри, П. Дж .; Langston, W. Jr; Танке, Д. Х. (2008), Новый рогатый динозавр из костного пласта верхнего мела в Альберте, Оттава, Онтарио: NRC Research Press, стр.1 –108, Дои:10.1139/9780660198194, ISBN 978-0-660-19819-4
- Dodson, P .; Forster, C.A .; Sampson, S. D. (2004), «Ceratopsidae», в Weishampel, D. B .; Dodson, P .; Осмольска, Х. (ред.), Динозаврия (2-е изд.), Беркли: Калифорнийский университет Press, стр.494 –513, ISBN 978-0-520-25408-4
- Хорнер, Дж. Р .; Добб, Э. (1997), Жизни динозавров: история эволюционной саги, Сан-Диего, Нью-Йорк, Лондон: Hartcourt Brace & Company
- Майорино, Л. (2015), Макроэволюционный паттерн у цератопсов динозавров (Dinosauria: Ornithischia) и биомеханика: комплексный подход с помощью геометрической морфометрии и анализа методом конечных элементов (Докторская диссертация), Università degli studi Roma Tre
- McDonald, A. T .; Фарке, А. А. (2011), "Незрелый образец Рубеозавр оватус (Dinosauria: Ceratopsidae), с наблюдениями за другими цератопсидами из формации Two Medicine », PLOS ONE, 6 (8): e22710, Bibcode:2011PLoSO ... 622710M, Дои:10.1371 / journal.pone.0022710, ЧВК 3154267, PMID 21853043
- Роджерс, Р. Р. (1989), Тафономия трех моноспецифических пластов костей динозавров в позднемеловой формации Two Medicine, северо-западная Монтана: данные о массовой смертности динозавров, связанной с эпизодической засухой (Магистерская диссертация), Университет Монтаны
- Райан, М. Дж .; Holmes, R .; Рассел, А. П. (2007), "Ревизия позднего кампанского рода центрозавринов ceratopsid" Стиракозавр из западных внутренних районов Северной Америки », Журнал палеонтологии позвоночных, 27 (4): 944–962, Дои:10.1671 / 0272-4634 (2007) 27 [944: AROTLC] 2.0.CO; 2
- Райан, М. Дж .; Chinnery-Allgeier, B.J .; Эберт, Д. А., ред. (2010), Новые взгляды на рогатых динозавров: Цератопсовский симпозиум Королевского музея Тиррелла, Издательство Индианского университета, ISBN 978-0-253-35358-0
- Форд, Т. Л., "Цератопсовский компендиум", в Райан, Чиннери-Аллгейер и Эберт (2010), Дополнительный CD-ROM
- Хендерсон, Д. М., «Формы черепа как индикаторы разделения ниши симпатрическими хазмозавринами и центрозавринами», в Райан, Чиннери-Аллгейер и Эберт (2010)
- Hunt, R .; Фарке, А., "Поведенческие интерпретации от цератопсидных костных пластов", в Райан, Чиннери-Аллгейер и Эберт (2010)
- McDonald, A. T .; Хорнер, Дж. Р., "Новый материал" Styracosaurus "ovatus из формации Two Medicine в Монтане", in Райан, Чиннери-Аллгейер и Эберт (2010)
- Райан, М. Дж .; Eberth, D. A .; Бринкман, Д. Б .; Карри, П. Дж .; Танке Д. Х. "Новое Пахиринозавр-Как цератопсид из формации Верхнего парка динозавров (поздний кампан) в Южной Альберте, Канада ", в Райан, Чиннери-Аллгейер и Эберт (2010)
- Sampson, S.D .; Лёвен, М. А., "Распутывая радиацию: обзор разнообразия, стратиграфического распределения, биогеографии и эволюции рогатых динозавров (Ornithischia: Ceratopsidae)", в Райан, Чиннери-Аллгейер и Эберт (2010)
- ВанБурен, С. С. (2013), Функция и эволюция синервикального канала у цератопсов динозавров с обзором слияния шейки матки у четвероногих (Магистерская диссертация), Университет Торонто
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||





