Инозитол-трифосфат-3-киназа - Inositol-trisphosphate 3-kinase - Wikipedia
| Инозитолтрифосфат-3-киназа | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Инозитол-трифосфат-3-киназа А. Каталитическое ядро. 1TZD | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 2.7.1.127 | ||||||||
| Количество CAS | 106283-10-7 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
Инозитол (1,4,5) трифосфат-3-киназа (ЕС 2.7.1.127 ), сокращенно здесь ITP3K, представляет собой фермент, который способствует переносу фосфогруппы из аденозинтрифосфат к 1D-мио-инозитол 1,4,5-трифосфат. Этот фермент принадлежит к семейству трансферазы, особенно те, которые переносят фосфорсодержащие группы (фосфотрансферазы ) с спиртовой группой в качестве акцептора. В систематическое название этого класса ферментов является АТФ: 1D-мио-инозитол-1,4,5-трифосфат-3-фосфотрансфераза. ITP3K катализирует перенос гамма-фосфата из АТФ на 3 позицию инозитол 1,4,5-трифосфат с образованием инозитола 1,3,4,5-тетракисфосфата.[1] ITP3K очень специфичен для 1,4,5-изомер IP3, и это исключительно фосфорилаты положение 3-ОН, производящее Ins (1,3,4,5) P4, также известный как тетракисфосфат инозита или IP4.
В биологии фермент ITP3K обозначается различными сокращениями, в том числе 1D-мио-инозитол-трифосфат-3-киназа, ITP3K, ИТПК, IP3-киназа, IP3-3-киназа, Ins (1,4,5) P3 3-киназа. Кроме того, фермент можно назвать продуктом одного из 3 генов человека. ИТПКА, ИТПКБ, и ITPKC, или один из двух у плодовых мух, IP3K1 и IP3K2 - мутант, известный генетикам как волнистый.[2] В геноме нематод есть одна форма фермента, кодируемая Ген LFE-2. Ферменты ITP3K экспрессируются только в многоклеточные животные; они не экспрессируются в дрожжах или растениях.
Все ITP3K принадлежат к более крупному структурному семейству, инозитолполифосфаткиназы, или IPK. Однако обратите внимание, что геном человека также содержит ген другой киназы, известной как ИТПК1, что является инозитол 1, 3, 4-трифосфат-5/6-киназа и не является членом семейства IPK.
Семейство ферментов ITP3K иногда путают с другим семейством ферментов, имеющим похожее название, то есть фосфатидилинозитол 3-киназы или же фосфоинозитид-3-киназа (PI3-K), субстратами которого являются липиды инозита, а не растворимый вторичный мессенджер трифосфат инозита.
Открытие и характеристика
Научный интерес к инозитолфосфатам усилился после открытия 1983 г. трифосфат инозита был внутриклеточным мессенджером, который высвобождает кальций из внутриклеточных хранилищ в эндоплазматическом ретикулуме.[3] К концу десятилетия было обнаружено большое количество инозитолфосфаткиназ и фосфатаз, включая ITP3K в 1986 году.[4][5]Биохимические и молекулярные исследования в 1990-х годах привели к очистке фермента из мозга крысы и его молекулярному клонированию, и эти исследования выявили различные механизмы обратной связи, с помощью которых фермент регулируется кальцием и протеинкиназами.[6] В 1999 году ITP3K был идентифицирован как член большой семьи Инозитолполифосфаткиназы, которые имеют схожую структуру и каталитический механизм.[7][8] Ферменты ITP3K имеют общие структурные особенности, включая консервативное каталитическое ядро, которое связывает АТФ, расположенный рядом с С-концом, и различные регуляторные домены ближе к N-концу.[9]
Каталитический домен
Доказательства этой исключительной специфичности и каталитического механизма были обнаружены, когда апофермент, комплекс, связанный с субстратом, и комплекс, связанный с продуктом Рентгеновские кристаллические структуры из ИТПКА были определены.[10][11] На рисунке справа изображен каталитический механизм, при котором 3'OH IP3 атакует гамма-фосфат АТФ и аминокислотные остатки ITPK, важные для стабилизации субстратов и продуктов в активном центре.
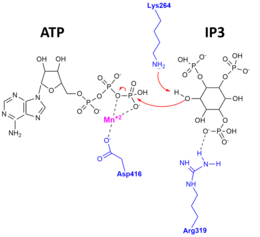
Было показано, что структура каталитического домена ITP3KA человека делится на три субдомена. Эти субдомены отображаются в виде N-доли, которая является N-концевым доменом, C-доли, которая является C-концевым поддоменом, и третьей альфа-субдоменом. Каталитический домен ITP3K несколько отличается от суперсемейства протеинкиназ и имеет новый четырехспиральный субстрат-связывающий домен. В этом киназа, два домена находятся в открытой конформации, что указывает на то, что оба домена доступны одновременно. Это говорит о том, что субстрат признание и катализ by ITP3K включает динамический конформационный цикл. Кроме того, этот уникальный спиральный домен ITPK блокирует доступ к активный сайт мембраносвязанными фосфоинозитидами, что объясняет структурную основу растворимый специфичность полифосфата инозита. Еще одна особенность каталитического ядра - сайт связывания АТФ. Здесь одна молекула ADP связан в щели основного домена, что указывает на активный сайт киназы.
Более подробно, большая область структура белка имеет структуру α / β-классов. Домен имеет N-концевую и C-концевую доли с щелью между ними, и каждая из этих долей построена вокруг антипараллельный β-лист. На N-конце лист состоит из трех нитей, тогда как на C-конце имеется пятицепочечный лист. Второй домен является α-спиральным и состоит из четырех α-спиралей, соединенных длинными петлями. Спирали слабо упакованы друг относительно друга, и весь домен очень подвижен по сравнению с большим доменом α / β. Спиральный домен прилегает к одному концу щели в большом домене.
Регулирование
ITP3K регулируется различными посттрансляционный механизмы. ITP3K стимулируются непосредственно кальций / кальмодулин (Ca2 + / CaM) привязка.[12] Как правило, ITP3K у млекопитающих в различной степени активируются кальцием и кальмодулином. Метод, в котором это работает, заключается в том, что кальмодулин распознает последовательности, которые содержат амфифильные альфа-спирали с кластерами положительно заряженных и гидрофобный аминокислоты.[13] Определенные последовательности требуются для связывания CaM и фермент активации, и этот уровень стимуляции, по-видимому, специфичен для клетки, ткани или изоформы. ITP3K от нематоды и Arabidopsis thaliana лишены CaM-связывающих сайтов и поэтому нечувствительны к кальцию и кальмодулину.[14] Еще одна важная посттрансляционная модификация, важная для регуляции ITP3K, - это фосфорилирование. Активность ITP3K косвенно стимулируется фосфорилированием кальций / кальмодулин-зависимая киназа II (CaMKII). Кроме того, есть доказательства того, что ITP3K могут активироваться при фосфорилировании посредством протеинкиназа C (PKC) и ингибируется при фосфорилировании протеинкиназа А (ПКА).
Изоформы
Есть три ITP3K, которые кодируются геномом человека: ИТПКА, ИТПКБ, и ITPKC. Все они имеют общий консервативный С-концевой каталитический домен, но различаются механизмами регуляции, а также тканевой экспрессией. ИТПКА преобладает в нейроны и в яички. Он локализован на дендритные шипы ассоциацией с нитчатым актин что согласуется с его вероятной ролью в функциях памяти. ИТПКБ экспрессируется более широко, но он часто обогащен иммунными тканями и имеет различные внутриклеточные локализации, которые зависят от ткани, взаимодействия с актиновыми филаментами и протеолиза в N-концевых областях. ITPKC также экспрессируется во многих различных тканях и в большей степени обогащен ядром по сравнению с другими изоформами.
Функции передачи сигналов кальция
ITP3K играет роль в регуляции или взаимодействии с внутриклеточными кальциевые сигналы которые происходят после освобождения трифосфат инозита. На этом пути либо Рецептор, связанный с G-белком (GPCR) или же рецепторная тирозинкиназа (RTK) активируется событием связывания внеклеточного лиганда. Инициирование пути приводит к активному Субъединица G-альфа из гетеротримерный G-белок (в случае GPCR-опосредованного преобразование сигнала ) или же аутофошорилирование цитоплазматических доменов RTK (в случае RTK-опосредованной передачи сигнала). Эти внутриклеточные события в конечном итоге приводят к активации фосфолипаза C (PLC), который расщепляет фосфолипид PIP2 в диацилглицерин (ДАГ) и инозитол 1,4,5-трифосфат (IP3). DAG остается связанным с плазматическая мембрана, а IP3 выпущен в цитоплазма. IP3 затем диффундирует через цитозоль и связывается с IP3 рецепторы на эндоплазматический ретикулум или же саркоплазматический ретикулум, что приводит к открытию мембранного канала и притоку кальций ионы в цитоплазму.[15] Кальций служит второй посланник для различных последующих событий сотовой связи, включая гликоген метаболизм, сокращение мышц, выпуск нейротрансмиттера, и транскрипционная регуляция.[15] Следовательно, гомеостаз кальция необходим для правильного функционирования клеток и ответа на внеклеточные сигналы.[16]
Чтобы подготовить клетку к будущему сигнальному событию, кальциевый путь должен быть строго отрегулирован. ITP3K, по-видимому, играет важную роль в прекращении сигнала. Как уже упоминалось, ITP3K катализирует фосфорилирование IP3 сделать IP4. В отличие от IP3, IP4 не вызывает открытия кальциевых каналов в эндоплазматическом или саркоплазматическом ретикулуме.[17] За счет снижения концентрации IP3 в цитоплазме ITP3K прекращает распространение кальциевого сигнального пути.[14]

Дополнительные роли
ITP3K - не единственный фермент, отвечающий за очистку IP3 из цитоплазмы. Второй фермент под названием инозитол-5-фосфатаза катализирует дефосфорилирование IP3 создать IP2.[18] Как правило, природа не одобряет развитие второго фермента, выполняющего уже существующую идентичную функцию.[19] Более пристальное рассмотрение эволюционной истории инозитол-5-фосфатазы и ITP3K дает начало нескольким интересным гипотезам о роли этих ферментов в клетке.
Инозитол-5-фосфатаза существовала до того, как ITP3K развился в клетке млекопитающих. Как и другие фосфатазы, инозитол-5-фосфатаза - это энергонезависимый фермент, который расщепляет фосфат группа вне субстрат.[20] Напротив, ITP3K (как и все киназы ) является энергетически зависимым, что означает, что для осуществления химии переноса фосфорила требуется молекула АТФ.[21] Если в природе уже был энергонезависимый механизм прекращения кальциевого сигнального пути, почему развитие ITP3K было выгодным? Это очевидное избыточность функции, или «растрата» энергии клеткой, предполагает, что ITP3K может иметь более важную функцию в клетке, чем простая очистка IP3 второй вестник из цитоплазмы.[20] Текущие гипотезы о дополнительных ролях для ИТПК объясняются в следующих двух подразделах.
Продукт ИТПК может быть вторым мессенджером
Как упоминалось ранее, ITP3K катализирует реакцию переноса фосфорила, которая превращает IP3 в IP4. IP4 не стимулирует приток кальция через ИП3 рецепторные каналы на эндоплазматическом или саркоплазматическом ретикулуме. Однако было показано, что IP4 стимулирует открытие кальциевых каналов на плазматической мембране. Таким образом, IP4 может фактически служить для продления кальциевого сигнала, активируя приток запасов кальция из внеклеточного пространства. Кроме того, есть свидетельства того, что IP4 связывает два Белки, активирующие ГТФазу, GAP1IP4BP и GAP1m.[18] GAP часто используются при преобразовании сигналов в качестве переключателей включения / выключения. IP4 Связывание с GAP предполагает, что ITPK может участвовать в параллельном пути передачи сигнала. Точная роль IP4 Однако привязка к этим GAP не определена, поэтому для более полного понимания потребуются дополнительные исследования в этой области.[22]
Роль в метаболизме инозитолфосфата
В дополнение к его потенциальной роли второй посланник, IP4 также может функционировать как важный предшественник для других более фосфорилированных фосфатов инозитола, таких как IP5, IP6, IP7, и IP8. Такое обслуживание необходимо для подготовки ячейки к будущему входящему сигналу.[22]
Актуальность для физиологии и болезней человека
ИТПКА Белок сильно обогащен дендритными шипами.[23] ITPKA участвует в процессах обучения и памяти в нейрональных клетках как за счет своей каталитической активности, так и за счет взаимодействия с нитчатым актином.
Несмотря на то что ИТПКА физиологически экспрессируется в нейронах и семенниках, ген становится экспрессируемым в ряде типов раковых клеток. В большинстве случаев экспрессия ITP3K делает рак более агрессивным.[24]
ИТПКБ участвует в физиологической иммунной функции.[25]
ITPKC был связан с болезнью Кавасаки, аутоиммунным заболеванием.[26][27]
Рекомендации
- ^ "UniProtKB - P23677 (IP3KA_HUMAN)". Получено 19 февраля 2015.
- ^ Dean DM, Maroja LS, Cottrill S, Bomkamp BE, Westervelt KA, Deitcher DL (ноябрь 2015 г.). «Волнистые карты мутаций гена инозитол-1,4,5-трифосфат-3-киназы 2 (IP3K2) дрозофилы и взаимодействуют с IP3R, влияя на развитие крыльев». G3. 6 (2): 299–310. Дои:10.1534 / g3.115.024307. ЧВК 4751550. PMID 26613949.
- ^ Streb H, Ирвин РФ, Берридж MJ, Шульц I (1983). «Высвобождение Ca2 + из немитохондриального внутриклеточного хранилища в ацинарных клетках поджелудочной железы с помощью инозитол-1,4,5-трифосфата». Природа. 306 (5938): 67–9. Bibcode:1983Натура 306 ... 67С. Дои:10.1038 / 306067a0. PMID 6605482. S2CID 4359904.
- ^ Ирвин РФ, Летчер AJ, Хеслоп JP, Берридж MJ (1986). «Инозитолтрис / тетракисфосфатный путь - демонстрация активности Ins (1,4,5) P3 3-киназы в тканях животных». Природа. 320 (6063): 631–4. Bibcode:1986Натура.320..631I. Дои:10.1038 / 320631a0. PMID 3010126. S2CID 4249596.
- ^ Hansen CA, Mah S., Williamson JR (июнь 1986 г.). «Образование и метаболизм инозитол 1,3,4,5-тетракисфосфата в печени». Журнал биологической химии. 261 (18): 8100–3. PMID 3487541.
- ^ Takazawa K, Vandekerckhove J, Dumont JE, Erneux C (ноябрь 1990 г.). «Клонирование и экспрессия в Escherichia coli кДНК головного мозга крысы, кодирующей Ca2 + / кальмодулин-чувствительную инозитол-1,4,5-трифосфат-3-киназу». Биохимический журнал. 272 (1): 107–12. Дои:10.1042 / bj2720107. ЧВК 1149663. PMID 2176078.
- ^ Saiardi A, Erdjument-Bromage H, Snowman AM, Tempst P, Snyder SH (ноябрь 1999 г.). «Синтез дифосфоинозитолпентакисфосфата недавно идентифицированным семейством высших полифосфаткиназ инозитола». Текущая биология. 9 (22): 1323–6. Дои:10.1016 / s0960-9822 (00) 80055-х. PMID 10574768. S2CID 15311443.
- ^ Odom AR, Stahlberg A, Wente SR, York JD (март 2000 г.). «Роль ядерной инозитол-1,4,5-трифосфаткиназы в контроле транскрипции». Наука. 287 (5460): 2026–9. Bibcode:2000Sci ... 287.2026O. Дои:10.1126 / science.287.5460.2026. PMID 10720331.
- ^ Шелл MJ (июнь 2010 г.). «Инозитолтрифосфат-3-киназы: фокус на иммунной и нейрональной передаче сигналов». Клеточные и молекулярные науки о жизни. 67 (11): 1755–78. Дои:10.1007 / s00018-009-0238-5. PMID 20066467. S2CID 25121695.
- ^ Гонсалес Б., Шелл М.Дж., Летчер А.Дж., Вепринцев Д.Б., Ирвин РФ, Уильямс Р.Л. (сентябрь 2004 г.). «Структура инозитол-1,4,5-трифосфат-3-киназы человека: связывание с субстратом показывает, почему это не фосфоинозитид-3-киназа». Молекулярная клетка. 15 (5): 689–701. Дои:10.1016 / j.molcel.2004.08.004. PMID 15350214.
- ^ Миллер Дж. Дж., Херли Дж. Х. (сентябрь 2004 г.). «Кристаллическая структура каталитического ядра инозитол-1,4,5-трифосфат-3-киназы». Молекулярная клетка. 15 (5): 703–11. Дои:10.1016 / j.molcel.2004.08.005. PMID 15350215.
- ^ Ллойд-Бертон С.М., Ю. Дж. К., Ирвин РФ, Шелл М. Дж. (Март 2007 г.). «Регулирование инозитол-1,4,5-трифосфат-3-киназы кальцием и локализация в клетках». Журнал биологической химии. 282 (13): 9526–35. Дои:10.1074 / jbc.M610253200. PMID 17284449.
- ^ Franco-Echevarría E, Baños-Sanz JI, Monterroso B, Round A, Sanz-Aparicio J, González B (ноябрь 2014 г.). «Новый кальмодулин-связывающий мотив для регуляции инозитол-1,4,5-трифосфат-3-киназы». Биохимический журнал. 463 (3): 319–28. Дои:10.1042 / BJ20140757. PMID 25101901.
- ^ а б Ся HJ, Ян Г (февраль 2005 г.). «Инозитол-1,4,5-трифосфат-3-киназы: функции и регуляция». Клеточные исследования. 15 (2): 83–91. Дои:10.1038 / sj.cr.7290270. PMID 15740635.
- ^ а б Берридж MJ (январь 1993 г.). «Инозитол трифосфат и передача сигналов кальция». Природа. 361 (6410): 315–25. Bibcode:1993Натура.361..315Б. Дои:10.1038 / 361315a0. PMID 8381210. S2CID 4362262.
- ^ Воет, Дональд Воет, Джудит Г. (2011). Биохимия (4-е изд.). Хобокен, Нью-Джерси: Джон Уайли и сыновья. ISBN 978-0-470-57095-1.
- ^ Havas N (август 2011 г.). «Снова в воду». Журнал паллиативной медицины. 14 (8): 971–2. Дои:10.1089 / jpm.2011.0043. PMID 21809925.
- ^ а б Паттни К., Бантинг Дж. (Июнь 2004 г.). «Метаболизм Ins (1,4,5) P3 и семейство киназ IP3-3». Сотовая связь. 16 (6): 643–54. Дои:10.1016 / j.cellsig.2003.10.009. PMID 15093605.
- ^ «Понимание эволюции». Получено 19 февраля 2015.
- ^ а б Ирвин РФ, Ллойд-Бертон С.М., Ю. Дж. К., Летчер А. Дж., Шелл М. Дж. (2006). «Регуляция и функция инозитол-1,4,5-трифосфат-3-киназы». Достижения в регуляции ферментов. 46 (1): 314–23. Дои:10.1016 / j.advenzreg.2006.01.009. ЧВК 1820747. PMID 16857241.
- ^ "WikiKinome". Kinase.com. Получено 19 февраля 2015.
- ^ а б Ирвин РФ, Шелл MJ (май 2001 г.). «Назад в воду: возвращение инозитолфосфатов». Обзоры природы. Молекулярная клеточная биология. 2 (5): 327–38. Дои:10.1038/35073015. PMID 11331907. S2CID 2259401.
- ^ Ямада М., Какита А., Мидзугути М., Ри С.Г., Ким Су, Икута Ф. (март 1993 г.). «Специфическая экспрессия инозитол-1,4,5-трифосфат-3-киназы в дендритных шипах». Исследование мозга. 606 (2): 335–40. Дои:10.1016 / 0006-8993 (93) 91004-c. PMID 8387863. S2CID 10790958.
- ^ Виндхорст С., Флигерт Р., Блехнер С., Мёлльманн К., Хоссейни З., Гюнтер Т., Эйбен М., Чанг Л., Лин Х.Й., Фаник В., Шумахер Ю., Брандт Б., Майр Г. В. (февраль 2010 г.). «Инозитол 1,4,5-трифосфат 3-киназа-A - это новый белок, способствующий подвижности клеток, который увеличивает метастатический потенциал опухолевых клеток за счет двух функциональных активностей».. Журнал биологической химии. 285 (8): 5541–54. Дои:10.1074 / jbc.M109.047050. ЧВК 2820782. PMID 20022963.
- ^ Миллер А.Т., Дальберг С., Сандберг М.Л., Вен Б.Г., Байснер Д.Р., Хортер Дж. А., Паркер А., Шмедт С., Стинсон М., Авис Дж., Сьенфуэгос С., Макпейт М., Трантер П., Гослинг М., Грут-Кормелинк П.Дж., Доусон Дж. Пан С., Тиан СС, Зайдель Х.М., Кук МП (2015). «Ингибирование инозитолкиназы Itpkb усиливает передачу сигналов кальция в лимфоцитах и раскрывает новую стратегию лечения аутоиммунного заболевания». PLOS ONE. 10 (6): e0131071. Bibcode:2015PLoSO..1031071M. Дои:10.1371 / journal.pone.0131071. ЧВК 4488288. PMID 26121493.
- ^ Onouchi Y, Gunji T, Burns JC, Shimizu C, Newburger JW, Yashiro M, Nakamura Y, Yanagawa H, Wakui K, Fukushima Y, Kishi F, Hamamoto K, Terai M, Sato Y, Ouchi K, Saji T, Nariai A , Kaburagi Y, Yoshikawa T., Suzuki K, Tanaka T, Nagai T, Cho H, Fujino A, Sekine A, Nakamichi R, Tsunoda T, Kawasaki T, Nakamura Y, Hata A (январь 2008 г.). «Функциональный полиморфизм ITPKC, связанный с восприимчивостью к болезни Кавасаки и образованием аневризм коронарных артерий». Природа Генетика. 40 (1): 35–42. Дои:10,1038 / нг.2007,59. ЧВК 2876982. PMID 18084290.
- ^ Альфонс М.П., Дуонг Т.Т., Шумицу К., Хоанг Т.Л., МакКриндл Б.В., Франко А., Шурманс С., Филпотт Д.Д., Хибберд М.Л., Бернс Дж., Куиджперс Т.В., Йунг Р.С. (ноябрь 2016 г.). «Инозитол-трифосфат-3-киназа С опосредует активацию инфламмасом и ответ на лечение при болезни Кавасаки». Журнал иммунологии. 197 (9): 3481–3489. Дои:10.4049 / jimmunol.1600388. PMID 27694492.