Вирус герпеса, связанный с саркомой Капоза - Kaposis sarcoma-associated herpesvirus - Wikipedia
| Человеческий гаммагерпесвирус 8 | |
|---|---|
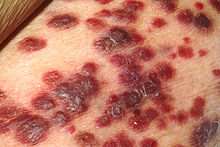 | |
| Саркома Капоши | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область: | Дуплоднавирия |
| Королевство: | Heunggongvirae |
| Тип: | Пепловирикота |
| Учебный класс: | Herviviricetes |
| Заказ: | Herpesvirales |
| Семья: | Herpesviridae |
| Род: | Радиновирус |
| Разновидность: | Человеческий гаммагерпесвирус 8 |
Вирус герпеса, связанный с саркомой Капоши (КШВ) является девятым известным человеком герпесвирус; его официальное название согласно Международный комитет по таксономии вирусов (ICTV) это Человеческий гаммагерпесвирус 8, или же HHV-8 короче.[1] Как и другие герпесвирусы, его неофициальные названия взаимозаменяемы с официальным названием ICTV. Этот вирус вызывает Саркома Капоши, рак, обычно встречающийся в СПИД пациенты,[2] а также первичная выпотная лимфома,[3] Многоцентровая болезнь Кастлемана, связанная с HHV-8 и Синдром воспалительных цитокинов KSHV.[4] Это один из семи известных в настоящее время вирусов рака человека, или онковирусы.[2] Даже после стольких лет открытия KSHV / HHV8 не существует известного лекарства от связанного с KSHV туморогенеза.
История
В 1872 г. Мориц Капоши описал опухоль кровеносного сосуда[5] (первоначально называвшаяся «идиопатическая множественная пигментная саркома кожи»), которая с тех пор носит одноименное название Саркома Капоши (KS). Сначала считалось, что СК является необычной опухолью среди еврейского и средиземноморского населения, но позже было установлено, что он чрезвычайно распространен среди африканского населения к югу от Сахары. Это привело к появлению первых предположений в 1950-х годах о том, что эта опухоль может быть вызвана вирусом. С наступлением СПИД эпидемии в начале 1980-х годов произошло внезапное возрождение СК, затронувшее в первую очередь гомосексуальных и бисексуальных пациентов со СПИДом, при этом до 50% зарегистрированных пациентов со СПИДом имели эту опухоль - чрезвычайно высокий уровень предрасположенности к раку.
Тщательный анализ эпидемиологических данных Валери Берал, Томасом Петерманом и Гарольдом Джаффе,[6] побудили этих исследователей предположить, что СК вызывается неизвестным вирусом, передающимся половым путем, который редко вызывает опухоли, если только хозяин не становится иммуносупрессивный, как в СПИДе.[нужна цитата ]

Еще в 1984 году ученые сообщили о том, что видели структуры, похожие на герпесвирус, в опухолях саркоидоза, исследованных под электронная микроскопия. Ученые занимались поиском агента, вызывающего KS, и в качестве возможной причины были предложены более 20 агентов, в том числе цитомегаловирус и ВИЧ сам. В конечном итоге возбудитель был идентифицирован в 1994 г. Юань Чанг и Патрик С. Мур, команда жены и мужа в Колумбийский университет, путем выделения фрагментов ДНК из герпесвирус обнаружен в опухоли СК у больного СПИДом.[7][8][9] Чанг и Мур использовали анализ репрезентативных различий, или RDA, чтобы найти KSHV, сравнивая опухолевую ткань KS пациента со СПИДом с его собственной непораженной тканью. Идея этого эксперимента заключалась в том, что если вирус вызывает СК, геномная ДНК в двух образцах должна быть абсолютно идентична, за исключением ДНК, принадлежащей вирусу. В своем первоначальном эксперименте RDA они выделили два небольших фрагмента ДНК, которые составляли менее 1% фактического вирусного генома. Эти фрагменты были похожи (но все еще отличны) от известных последовательностей герпевируса, что указывало на присутствие нового вируса. Исходя из этих фрагментов, менее чем через два года исследовательская группа смогла секвенировать весь геном вируса.[нужна цитата ]
Открытие этого герпесвирус вызвали серьезные споры и научные споры, пока не было собрано достаточно данных, чтобы показать, что действительно KSHV был возбудителем саркомы Капоши.[10] В настоящее время известно, что этот вирус является широко распространенной инфекцией людей, живущих в Африке к югу от Сахары; промежуточные уровни инфекции наблюдаются в средиземноморских популяциях (включая Ливан, Саудовскую Аравию, Италию и Грецию), а низкие уровни инфекции наблюдаются в большинстве групп населения Северной Европы и Северной Америки. Мужчины-геи и бисексуалы более восприимчивы к инфекции (до сих пор неизвестными путями передачи половым путем), тогда как в развивающихся странах вирус передается несексуальным путем.[нужна цитата ]
Вирусология
КШВ - это герпесвирус, и представляет собой большой двухцепочечный ДНК-вирус с белковым покрытием, которое упаковывает его нуклеиновые кислоты, называемый капсид, который затем окружен слоем аморфного белка, называемым оболочка и, наконец, заключен в липидную оболочку, частично происходящую из клеточной мембраны. КШВ имеет геном что составляет приблизительно 165 000 оснований нуклеиновой кислоты в длину. КШВ - это радиновирус, род герпеса, замечательный, поскольку он украл множество генов из клеток-хозяев, в том числе в случае генов KSHV, которые кодируют дополнять -связывающий белок, Ил-6, BCL-2, циклин -D, а Рецептор, связанный с G-белком, регуляторный фактор интерферона и белок, ингибирующий Flice (КУВЫРОК ), а также белки синтеза ДНК, в том числе дигидрофолатредуктаза, тимидинкиназа, тимидилатсинтетаза, ДНК полимераза и много других. Хотя ни один другой опухолевый вирус человека не обладает такими же генами, другие опухолевые вирусы нацелены на одни и те же клеточные пути, демонстрируя, что на базовом уровне все опухолевые вирусы, по-видимому, атакуют одни и те же пути клеточного контроля, так называемые пути подавления опухоли.[нужна цитата ]
После заражения вирус попадает в лимфоциты через макропиносомы где он остается в скрытом («тихом») состоянии. Только подмножество генов, которые кодируются в латентно-ассоциированной области (KLAR) KSHV, экспрессируются в латентный период, включая ассоциированный с латентностью ядерный антиген (LANA), vFLIP, vCyclin и 12 микроРНК. Латентный период является отличительной чертой всех известных на сегодняшний день этиологий, связанных с KSHV, включая весь онкогенез, связанный с KSHV. Было показано, что как гены, кодирующие белок, так и некодирующий ген (микроРНК), кодируемый в области KLAR, важны для связанного с KSHV туморогенеза. Для изучения функций микроРНК ведущими исследователями в этой области были созданы подробный протокол бактериального мутагенеза и полный набор клеточных линий, несущих мутанты с делецией микроРНК, которые доступны в качестве ресурса для любого международного исследователя, работающего в области вирусов. -ассоциированный рак.[11] Кроме того, в другом исследовании было показано, что vFLIP и vCyclin косвенно вмешиваются в сигнальный путь TGF-β, индуцируя кластер mir17-92 онкогенного хозяина.[12] Эти наблюдения представляют собой новый механизм, который может быть важным для онкогенеза и ангиогенеза KSHV, отличительного признака KS. Разработка важных инструментов, таких как полный набор из 12 мутантов с делецией микроРНК, является важным шагом в изучении функций гена KLAR в контексте канцерогенеза, связанного с KSHV. [13] Решающий для входа в КШВ [14] это EPH рецептор A2,[15] Часы,[16] TSG101[17] и несколько Интегрины, личность которого еще не подтверждена.[18] Вирус существует в виде кольцевого фрагмента ДНК, называемого эписома и использует аппарат клеточной репликации для самовоспроизведения. LANA - единственный вирусный белок, который необходим для репликации вирусной ДНК в латентный период. При попадании в ядро клетки-хозяина вирусная эписома хроматинируется.[19] LANA связывает вирусную ДНК с клеточными хромосомами, подавляет p53 и белок ретинобластомы и подавляет вирусные гены, необходимые для полной продукции и сборки вируса («литическая репликация»). Почему только часть вирусных генов экспрессируется в латентный период, не совсем понятно. Но было показано, что связанная с латентным периодом экспрессия генов может быть частично объяснена характерным эпигенетическим состоянием, которое эписома KSHV приобретает во время латентного периода. LANA играет важную роль во время задержки. Показано, что он регулирует транскрипты как хозяина, так и вируса и связывается с множеством активных промоторов, включая промоторы хозяина, в течение латентного периода.[20] В том же исследовании также было показано, что LANA ассоциирует с хост-машиной hSET1, которая создает метки H3K4me3.[20] Однако точный механизм, почему область KLAR остается активной во время задержки, не известен. Также показано, что различные сигналы, такие как воспаление, могут спровоцировать начало литической репликации вируса. Когда происходит литическая репликация, вирусная эписома начинает реплицироваться в форме линейных молекул ДНК, которые упаковываются в вирусные частицы, которые изгоняются из клетки. , чтобы заразить новые клетки или передать новому хозяину. Когда вирус начинает литическую репликацию, тысячи вирусных частиц могут быть получены из одной клетки, что обычно приводит к гибели инфицированной клетки.[нужна цитата ]
Вирусный геном состоит из уникальной области длиной ~ 145 т.п.н., кодирующей все экспрессируемые вирусные гены, которая фланкирована ~ 20-30 т.п.н. концевых повторяющихся последовательностей.[21] Каждая конечная повторяющаяся единица имеет длину 801 п.н., содержит 85% G + C и ориентирована повторяющимся образом от головы к хвосту. Во время латентного периода геном вируса зависит от аппарата репликации хозяина и реплицируется в виде замкнутой кольцевой эписомы («плазмиды») с использованием последовательностей в концевых повторах в качестве точки начала репликации. Когда вирус реактивируется в литическую репликацию, считается, что вирусный геном реплицируется как непрерывная линейная молекула, выходящая из эписомы (так называемая модель катящегося круга). По мере репликации каждой единицы генома он разрезается в области концевого повтора, а затем упаковывается в вирусную частицу (вирион). Затем вирус покрывается липидной мембраной, когда он проходит через ядро и цитоплазму, чтобы покинуть клетку. Таким образом, в то время как геном KSHV является кольцевым в ядре латентно инфицированных клеток, он упакован в инфекционные вирусы в виде линейной молекулы. Как только вирус вновь заражает клетку, липидная мембрана сбрасывается, и вирион перемещается к ядру. Вирусный геном высвобождается там, где он рециркуляризируется посредством плохо изученного процесса, который, как представляется, включает гомологичная рекомбинация.[нужна цитата ]
Первичный вирусный белок, ответственный за переключение между латентной и литической репликацией, известен как активатор трансактивации репликации ORF50 (RTA). Когда условия клеточной передачи сигналов активируют генерацию RTA, это, в свою очередь, активирует синтез стереотипного каскада вторичных и третичных вирусных белков, которые в конечном итоге составляют компоненты вирусного капсида, а также ферменты синтеза ДНК, необходимые для репликации вирусного генома.[22]
Патофизиология
Механизмы заражения вирусом изучены недостаточно. Здоровые люди могут быть инфицированы вирусом и не проявлять никаких признаков или симптомов из-за способности иммунной системы сдерживать инфекцию. Инфекция вызывает особую озабоченность с ослабленным иммунитетом. Больные раком, получающие химиотерапию, больные СПИДом и пациенты с трансплантацией органов, все имеют высокий риск появления признаков инфекции.[нужна цитата ].
Последние достижения в области технологий секвенирования показали, что вирус хроматинируется во время латентного периода. Также было показано, что кодируемая вирусом микроРНК манипулирует и взаимодействует не только с мРНК хозяина, но также нарушает регуляцию длинной некодирующей РНК хозяина (днРНК).[23] Совсем недавно кольцевые РНК (circRNA) были обнаружены как у EBV, так и у KSHV. [24][25][26]
Считается, что заражение этим вирусом продолжается всю жизнь, но здоровая иммунная система будет держать вирус под контролем. Многие люди, инфицированные KSHV, никогда не проявляют никаких симптомов. Саркома Капоши возникает, когда человек, инфицированный KSHV, становится ослабленным из-за СПИДа, лечения или очень редко старения.
КШВ - известный возбудитель четырех заболеваний:[4][27]
- Саркома Капоши - ангиопролиферативная опухоль, которая может поражать кожу (чаще всего), лимфатические узлы или внутренние органы,
- Мультицентрическая болезнь Кастлемана, связанная с HHV-8 - а лимфопролиферативное заболевание,
- Лимфома с первичным выпотом - KSHV-ассоциированная агрессивная лимфома с вовлечением незрелых В-клеток плазматические клетки называемые плазмобластами и рассматриваемые как один тип лимфоидные новообразования с плазмобластной дифференцировкой,
- Синдром воспалительных цитокинов KSHV - синдром с симптомами, подобными МКД, но без сопутствующей патологии.
Эпидемиология
В 1970-е годы глобальный уровень распространенности HHV-8 составлял от 2 до 10%.[28] Распространенность HHV-8 значительно различается географически, а уровень инфицирования в странах Северной Европы, Юго-Восточной Азии и Карибского бассейна составляет 2-4%.[29] в странах Средиземноморья - примерно 10%, а в странах Африки к югу от Сахары - примерно 40%.[30] В Южной Америке уровень инфицирования в целом низкий, но высокий среди Индейцы.[31] Даже в пределах отдельных стран можно наблюдать значительные различия в разных регионах, при этом уровень инфицирования составляет около 19,2% в Синьцзян по сравнению с примерно 9,5% в Хубэй, Китай.[32] Хотя было показано, что распространенность серотипа линейно увеличивается с возрастом,[32][33][34][35] в странах с высоким уровнем инфицирования может наблюдаться более высокая распространенность серотипа в более молодых возрастных группах.[36] Уровень образования показал обратную корреляцию с уровнем инфицирования.[33][34] Люди, инфицированные ВИЧ-1 или остроконечными кондиломами, как правило, более подвержены коинфекции HHV-8.[30][32][37]
В странах с низкой распространенностью ВИЧ-инфекции HHV-8 в основном ограничивается пациентами со СПИДом и СК.[38] В странах с высокой серологической распространенностью инфекция часто встречается в детстве,[39] указывая на вероятный передача от матери ребенку по слюне.[40][41] В Замбийский По данным исследования, у всех детей с СК были матери, положительные на HHV-8, тогда как не все дети, матери которых были с СК, были положительными на HHV-8.[42] По данным другого исследования в Замбии, 13,8% детей были серопозитивными по HHV-8 к 4 годам.[43] Не было показано, что распространенность серопротеина существенно зависит от пола или семейного положения.[35]
Эволюция
В самый последний общий предок этого вируса в Средиземном море, Иране и Синьцзяне, Китай, по оценкам, эволюционировал 29 872 года (95% наивысшая плотность вероятности 26 851-32 760 лет) назад.[44] самый недавний общий предок вирусов, выделенных в Синьцзяне, был зарегистрирован 2037 лет назад (95% самая высокая плотность вероятности 1843–2229 лет) назад. Учитывая исторические связи между Средиземным морем и Синьцзяном во время Римский период кажется вероятным, что этот вирус был завезен в Синьцзян во время Шелковый путь. Частота мутаций составила 3,44 × 10−6 замен на сайт в год (95% наивысшая плотность вероятности от 2,26 × -6 до 4,71 × 10−6). Однако глобальное распространение различных генотипов KSHV и потенциальный путь передачи требуют дальнейших исследований.[нужна цитата ]
Типирование изолятов основано на вариабельном мембранном белке K1. Различают шесть типов (A-F).[45]
Профилактика
Поскольку люди, инфицированные KSHV, передают вирус бессимптомно, следует соблюдать осторожность. половые партнеры в наличии незащищенный секс и действия, при которых слюна может выделяться во время сексуальной активности. Разумный совет - использовать презервативы при необходимости и избегать глубокие поцелуи с партнерами с KSHV и ВИЧ-инфекцией или чей статус неизвестен. Есть анализы крови на антитела к вирусу.[нужна цитата ]
Уход
Саркома Капоши обычно представляет собой локализованную опухоль, которую можно лечить хирургическим путем или с помощью местного облучения. Химиотерапия препаратами, например липосомальными антрациклины или же паклитаксел может использоваться, особенно при инвазивном заболевании. Противовирусные препараты, такие как ганцикловир, которые нацелены на репликацию герпесвирусов, таких как KSHV, были использованы для успешного предотвращения развития саркомы Капоши,[46] хотя, как только опухоль разовьется, от этих препаратов мало или вообще нет пользы. Для больных СПИД-СК наиболее эффективной терапией является высокоактивная антиретровирусная терапия для снижения ВИЧ-инфекции.[47] Пациенты со СПИДом, получающие адекватное лечение против ВИЧ, могут иметь до 90% снижение частоты возникновения саркомы Капоши.
Хотя KSHV влияет на иммунную систему хозяина, клиническое вмешательство может исправить это изменение. Одной из проблем является ингибирование сверхэкспрессии иммунного подавления клеток-мишеней. При длительной воспалительной стимуляции клетка-мишень не может ответить, что приводит к истощенному фенотипу. Активационная иммунотерапия может оживить и улучшить функцию иммунных клеток. По сравнению с другими видами иммунотерапии, терапии, нацеленные на белок 1, направленный против запрограммированной гибели клеток (ПД-1 ) / запрограммированная смерть-лиганд 1 (PD-L1 ) имел большой успех. Из-за инфекции KSHV моноциты увеличивают экспрессию PD-1, который является ингибирующей молекулой, и вызывают ускользание иммунитета во многих типах опухолей. В NK-клетках пациентов с KS-HIV наблюдается высокая экспрессия PD-1, что вызывает истощенный фенотип. Антитело против PD-1, (ниволумаб или же пембролизумаб ), продемонстрировал значительный противоопухолевый эффект. Ниволумаб в настоящее время находится на стадии I клинических испытаний, а пембролизумаб продемонстрировал свою функцию в лечении пациентов с ВИЧ и СК в фазе I и проходит II фазу испытаний для лечения. Лекарство-аналог талидомида - Помалидомид также был предоставлен FDA в 2011 году. Было показано, что помалидомид восстанавливает экспрессию MHC-1, которые помогают клетке отображать внутриклеточные белки цитотоксическим Т-клеткам, а также могут подавлять экспрессию PD-L1 и увеличивать гибель CD8 + Т-клеток.[48]
Гены KSHV
KSHV кодирует ~ 90 генов и множество некодирующих РНК, таких как микроРНК.[49] Гены ORF названы на основе положения в геноме гомологичных генов первого описанного радиновируса, вируса герпеса saimiri. Гены «K» уникальны для KSHV. Некоторые гены KSHV обладают хорошо охарактеризованными функциями, в то время как другие остаются не охарактеризованными.
ORF2 - дигидрофолатредуктаза
ORF8 - gB - конверт гликопротеин участвует в проникновении вируса
ORF9 - Pol8 - ДНК-полимераза требуется для репликации вирусной ДНК
ORF10 - регулирует экспорт РНК и ответы на IFN типа I
ORF16 - vBcl2
ORF18, ORF24, ORF30, ORF31, ORF34, ORF66 - факторы вирусной транскрипции, необходимые для экспрессии поздних генов
ORF21 - vTK - тимидинкиназа
ORF22 - gH - конверт гликопротеин участвует в проникновении вируса
ORF23 - без характеристики
ORF25, ORF26 и ORF65 - капсид белки
ORF33 - участвует в образовании вирусных частиц
ORF34 - неясная функция
ORF35 - функция неясна, мутант не экспрессирует ранние вирусные гены
ORF36 - vPK - вирусная протеинкиназа с множеством ролей в цикле репликации
ORF37 - SOX - белок двойной функции - ДНКаза активность, необходимая для упаковки генома и РНКаза активность регулирует экспрессию гена хозяина
ORF38 - участвует в образовании вирусных частиц
ORF39 - gM - конверт гликопротеин
ORF40 и ORF41 - геликаза и прима - Репликация ДНК
ORF42 - без характеристики
ORF45 - оболочка белок, связывает и предотвращает дефосфорилирование p90 рибосомальных киназ S6 (RSK) и ERK для модуляции ERK / RSK MAPK сигнальный путь
ORF47 - gL - гликопротеин оболочки, участвующий в проникновении вируса
ORF49 - может потребоваться для экспрессии вирусного гена
ORF50 - RTA, активатор репликации и транскрипции - основной фактор транскрипции, приводящий к литической реактивации KSHV
ORF52 - KicGAS - оболочка белок, необходимый для образования вирионов и ингибирования cGAS Зондирование ДНК
ORF53 - gN - гликопротеин оболочки
ORF55 - без характеристики
ORF57 - MTA - регулирует стабильность РНК, экспорт и трансляцию вирусных генов
ORF59 - PF – 8 - фактор процессивности полимеразы, дополнительная субъединица вирусной ДНК-полимеразы
ORF67 и ORF69 - ядерный выход
ORF70 - тимидилатсинтаза
ORF72 - vЦиклин
ORF73 - LANA, связанный с латентностью ядерный антиген - связывает геном с хромосомой во время латентного периода, а также регулирует экспрессию гена хозяина. Цитоплазматическая форма LANA может подавлять активацию иммунных ответов.
ORF74 - vGPCR
ORF75 - ФГАРАТ
СКОВОРОДА, полиаденилированный ядерный РНК - некодирующие линейные и кольцевые РНК
миРНК (mirKs) - вирусные микроРНК, экспрессируемые во время латентного периода для регулирования пролиферации и гибели клеток
K1 - участвует в онкогенезе
K2 - Интерлейкин 6 гомолог Q2HRC7
К3 и К5 - убиквитин Лигазы Е3 - регулировать презентацию антигена
K4 - vCCL2 - хемокин
K4.1 - vCCL3 - хемокин
K8 - репрессор транскрипции - модулирует хроматин
К8.1 - конверт гликопротеин
К9 - vIRF 1, вирусный регуляторный фактор интерферона 1
K10 - vIRF 4. Кольцевая РНК (circRNA ) также генерируется из этого локус.
K10.5 - vIRF3 / LANA2 - выражается во время задержки в В-клетки
К11 - vIRF 2
К12 - капосин
К13 - vКУВЫРОК
Смотрите также
- Онковирус (вирус рака)
- 22-й международный семинар по герпесвирусу саркомы Карпози (KSHV) и родственным агентам, 30 июня - 3 июля 2019 г., Нью-Йорк, США
Рекомендации
- ^ «Список основных видов ICTV 2018b.v2». Международный комитет по таксономии вирусов. Получено 13 июн 2019.
- ^ а б Boshoff, C .; Вайс, Р. (2002). «Злокачественные новообразования, связанные со СПИДом». Обзоры природы Рак. 2 (5): 373–382. Дои:10.1038 / nrc797. PMID 12044013. S2CID 13513517.
- ^ Cesarman, E .; Chang, Y .; Мур, П. С .; Said, J. W .; Ноулз, Д. М. (1995). «Связанные с саркомой Капоши последовательности ДНК, подобные герпесвирусу, в лимфомах, связанных со СПИДом». Медицинский журнал Новой Англии. 332 (18): 1186–1191. Дои:10.1056 / NEJM199505043321802. PMID 7700311.
- ^ а б Goncalves, Priscila H .; Зигельбауэр, Джозеф; Улдрик, Томас С .; Ярчоан, Роберт (2017). «Раковые заболевания, связанные с герпесвирусом саркомы Капоши, и связанные с ними заболевания». Curr Opin ВИЧ СПИД. 12 (1): 47–56. Дои:10.1097 / COH.0000000000000330. ЧВК 6311702. PMID 27662501.
- ^ Капоши, М. (1872). «Идиопатический множественный Пигментарком дер Га». Архив дерматологии и сифилиса. 4 (2): 265–273. Дои:10.1007 / BF01830024. S2CID 31438763.
Переведено на Капоши, М. (2008). «Идиопатическая множественная пигментная саркома кожи». КА: Онкологический журнал для клиницистов. 32 (6): 342–347. Дои:10.3322 / canjclin.32.6.342. S2CID 72037330. - ^ Берал В., Петерман Т.А., Беркельман Р.Л., Яффе Х.В. (1990). «Саркома Капоши у больных СПИДом: инфекция, передающаяся половым путем?». Ланцет (Представлена рукопись). 335 (8682): 123–8. Дои:10.1016 / 0140-6736 (90) 90001-Л. PMID 1967430. S2CID 35639169.
- ^ Chang, Y .; Cesarman, E .; Пессин, М. С .; Ли, Ф .; Culpepper, J .; Ноулз, Д. М .; Мур, П. С. (1994). «Идентификация последовательностей ДНК, подобных герпесвирусу, в СПИД-ассоциированной саркоме Капоши». Наука. 266 (5192): 1865–1869. Bibcode:1994Наука ... 266.1865C. Дои:10.1126 / science.7997879. PMID 7997879.
- ^ Мур PS, Чанг И (май 1995 г.). «Обнаружение последовательностей ДНК, подобных герпесвирусу, в саркоме Капоши у пациентов с ВИЧ-инфекцией и без нее». Медицинский журнал Новой Англии. 332 (18): 1181–5. Дои:10.1056 / NEJM199505043321801. PMID 7700310. S2CID 13465834.
- ^ Antman, K .; Чанг, Ю. (2000). «Саркома Капоши». Медицинский журнал Новой Англии. 342 (14): 1027–1038. Дои:10.1056 / NEJM200004063421407. PMID 10749966.
- ^ Boshoff C, Weiss RA (май 2000 г.). «Разрешение споров по поводу саркомы Капоши». Журнал Национального института рака. 92 (9): 677–679. Дои:10.1093 / jnci / 92.9.677. PMID 10793096.
- ^ Джайн, вайбхав; Плезанс-Бонстафф, Карли; Сангани, Раджникумар; Ланье, Кертис; Дольче, Александр; Ху, Цзяньхун; Брулуа, Кевин; Хеккер, Ирина; Тернер, Питер; Ренне, Рольф; Крюгер, Брайан (19 февраля 2016 г.). «Набор инструментов для исследования миРНК герпесвируса: создание полного набора мутантов для деления миРНК KSHV». Вирусы. 8 (2): 54. Дои:10.3390 / v8020054. ISSN 1999-4915. ЧВК 4776209. PMID 26907327.
- ^ Чой, HS; Джайн, V; Крюгер, В; Маршалл, В .; Kim, CH; Шислер, JL; Whitby, D; Ренне, Р. (2015). «Связанный с саркомой герпесвирус Капоши (KSHV) индуцирует онкогенный кластер miR-17-92 и подавляет передачу сигналов TGF-β». Патогены PLOS. 11 (11): e1005255. Дои:10.1371 / journal.ppat.1005255. ЧВК 4636184. PMID 26545119.
- ^ Джайн, V; Plaisance-Bonstaff, K; Sangani, R; Lanier, C; Dolce, A; Ху, Дж; Brulois, K; Хеккер, я; Тернер, П; Renne, R; Крюгер, Б. (19 февраля 2016 г.). «Набор инструментов для исследования миРНК герпесвируса: создание полного набора мутантов для деления миРНК KSHV». Вирусы. 8 (2): 54. Дои:10.3390 / v8020054. ЧВК 4776209. PMID 26907327.
- ^ Кумар, Бинод; Чандран, Бала (14 ноября 2016 г.). «Вхождение и торговля KSHV в клетках-мишенях - захват клеточных сигнальных путей, актина и мембранной динамики». Вирусы. 8 (11): 305. Дои:10.3390 / v8110305. ISSN 1999-4915. ЧВК 5127019. PMID 27854239.
- ^ Neipel F, Fleckenstein B, Hahn A и др. (Июнь 2012 г.). «Тирозинкиназа А2 рецептора эфрина является клеточным рецептором вируса герпеса, связанного с саркомой Капоши». Природа Медицина. 18 (6): 961–968. Дои:10,1038 / нм.2805. ЧВК 3645317. PMID 22635007.
- ^ Веттил, Моханан Валия; Кумар, Бинод; Ансари, Майрадж Ахмед; Дутта, Дипанджан; Икбал, Джавед; Гжыши, Олси; Боттеро, Вирджиния; Чандран, Бала (апрель 2016 г.). «Компонент Hrs ESCRT-0 способствует макропиноцитозу герпесвируса, ассоциированного с саркомой Капоши, в эндотелиальных клетках кожных микрососудов человека». Журнал вирусологии. 90 (8): 3860–3872. Дои:10.1128 / JVI.02704-15. ISSN 1098-5514. ЧВК 4810545. PMID 26819309.
- ^ Кумар, Бинод; Дутта, Дипанджан; Икбал, Джавед; Ансари, Майрадж Ахмед; Рой, Арунава; Чикоти, Лила; Пизано, Джина; Веттил, Моханан Валия; Чандран, Бала (октябрь 2016 г.). «Белок ESCRT-I Tsg101 играет роль в постмакропиноцитарном трафике и инфицировании эндотелиальных клеток герпесвирусом Капоши, ассоциированным с саркомой». Патогены PLOS. 12 (10): e1005960. Дои:10.1371 / journal.ppat.1005960. ISSN 1553-7374. ЧВК 5072609. PMID 27764233.
- ^ Кампаделли-Фьюме Дж., Коллинз-МакМиллен Д. и др. (Июль 2016 г.). «Интегрины как рецепторы герперсвирусов и медиаторы сигнаносомы хозяина». Ежегодный обзор вирусологии. 3 (1): 215–236. Дои:10.1146 / annurev-virology-110615-035618. PMID 27501260.
- ^ Пурушотаман и др. (Февраль 2016). «Репликация и поддержание генома KSHV». Границы микробиологии. 7: 54. Дои:10.3389 / fmicb.2016.00054. ЧВК 4740845. PMID 26870016.
- ^ Руссо Дж. Дж., Богенцкий Р. А., Чиен М. С. и др. (Декабрь 1996 г.). «Нуклеотидная последовательность вируса герпеса, связанного с саркомой Капоши (HHV8)». Труды Национальной академии наук Соединенных Штатов Америки. 93 (25): 14862–7. Bibcode:1996PNAS ... 9314862R. Дои:10.1073 / пнас.93.25.14862. ЧВК 26227. PMID 8962146.
- ^ Бу В., Палмери Д., Кришнан Р. и др. (Ноябрь 2008 г.). «Идентификация прямых транскрипционных мишеней белка литического переключателя Rta, ассоциированного с саркомой Капоши, путем условной ядерной локализации». Журнал вирусологии. 82 (21): 10709–23. Дои:10.1128 / JVI.01012-08. ЧВК 2573185. PMID 18715905.
- ^ Сетураман, S; Гей, Лос-Анджелес; Джайн, V; Хеккер, я; Ренне, Р. (июль 2017 г.). «Зависимая от микроРНК и независимая дерегуляция длинных некодирующих РНК онкогенным вирусом герпеса». Патогены PLOS. 13 (7): e1006508. Дои:10.1371 / journal.ppat.1006508. ЧВК 5531683. PMID 28715488.
- ^ Топтан, Т; Абере, Б; Налесник, Массачусетс; Swerdlow, SH; Ранганатан, S; Ли, Н.; Шаир, К.Х .; Мур, ПС; Чанг, И (11 сентября 2018 г.). «Циркулярные ДНК опухолевых вирусов образуют кольцевые РНК». Труды Национальной академии наук Соединенных Штатов Америки. 115 (37): E8737 – E8745. Дои:10.1073 / pnas.1811728115. ЧВК 6140489. PMID 30150410.
- ^ Тагава, Т; Gao, S; Копарде, ВН; Gonzalez, M; Spouge, JL; Serquiña, AP; Лурайн, К; Рамасвами, Р. Uldrick, TS; Ярчоан, Р. Зигельбауэр, JM (11 декабря 2018 г.). «Открытие кольцевых РНК, кодируемых герпесвирусом саркомы Капоши, и противовирусной кольцевой РНК человека». Труды Национальной академии наук Соединенных Штатов Америки. 115 (50): 12805–12810. Дои:10.1073 / pnas.1816183115. ЧВК 6294913. PMID 30455306.
- ^ Унгерлейдер, штат Северная Каролина; Джайн, V; Ван, Y; Манесс, штат Нью-Джерси; Blair, RV; Альварес, X; Midkiff, C; Колсон, Д; Bai, S; Робертс, К; Moss, WN; Ван, Х; Serfecz, J; Седдон, М; Lehman, T; Ма, Т; Донг, Й; Renne, R; Tibbetts, SA; Флемингтон, EK (15 марта 2019 г.). «Сравнительный анализ репертуаров кольцевой РНК гаммагерпесвируса: консервативные и уникальные вирусные кольцевые РНК». Журнал вирусологии. 93 (6). Дои:10.1128 / JVI.01952-18. ЧВК 6401440. PMID 30567979.
- ^ Чен Би Джей, Чуанг СС (март 2020 г.). «Лимфоидные новообразования с плазмобластной дифференцировкой: всесторонний обзор и диагностические подходы». Достижения в анатомической патологии. 27 (2): 61–74. Дои:10.1097 / PAP.0000000000000253. PMID 31725418.
- ^ Галло, Роберт К. (1998-12-04). "Загадки саркомы Капоши". Наука. 282 (5395): 1837–1839. Дои:10.1126 / science.282.5395.1837. ISSN 0036-8075. PMID 9874635.
- ^ Чжан, Тиецзюнь; Ван, Линдинг (2016). «Эпидемиология герпесвируса, связанного с саркомой Капоши в Азии: проблемы и возможности». Журнал медицинской вирусологии. 89 (4): 563–570. Дои:10.1002 / jmv.24662. ISSN 0146-6615. PMID 27531516.
- ^ а б Chatlynne, L.G .; Аблаши, Д. В. (1999). "Сероэпидемиология герпесвируса, связанного с саркомой Капоши (KSHV)". Семинары по биологии рака. 9 (3): 175–85. Дои:10.1006 / scbi.1998.0089. PMID 10343069.
- ^ Моханна, S; Maco, V; Браво, F; Готуццо, Э (2005). «Эпидемиология и клинические характеристики классической саркомы Капоши, серологическая распространенность и варианты вируса герпеса человека 8 в Южной Америке: критический обзор старой болезни». Международный журнал инфекционных болезней. 9 (5): 239–50. Дои:10.1016 / j.ijid.2005.02.004. PMID 16095940.
- ^ а б c Fu, B; Вс, F; Ли, Б; Ян, Л; Zeng, Y; Солнце, X; Сюй, Ф; Rayner, S; Guadalupe, M; Gao, S.J .; Ван, Л. (2009). "Распространенность вируса герпеса, связанного с саркомой Капоши, и факторы риска в Синьцзяне, Китай". Журнал медицинской вирусологии. 81 (8): 1422–31. Дои:10.1002 / jmv.21550. ЧВК 2755560. PMID 19551832.
- ^ а б Пелсер, К; Vitale, F; Whitby, D; Graubard, B.I .; Мессина, А; Gafà, L; Brown, E. E .; Андерсон, Л. А .; Романо, N; Лаурия, К; Гёдерт, Дж. Дж. (2009). «Социально-экономические и другие корреляты серологической распространенности герпесвируса, связанного с саркомой Капоши, среди пожилых людей на Сицилии». Журнал медицинской вирусологии. 81 (11): 1938–44. Дои:10.1002 / jmv.21589. ЧВК 2784645. PMID 19777527.
- ^ а б Wang, H; Лю, Дж; Дилимулати; Ли, Л; Рен, Z; Вен, H; Ван, X (2011). «Распространенность серотипа и факторы риска герпесвирусной инфекции, связанной с саркомой Капоши, среди уйгурского населения в целом из южного и северного региона Синьцзяна, Китай». Журнал вирусологии. 8: 539. Дои:10.1186 / 1743-422X-8-539. ЧВК 3266657. PMID 22168313.
- ^ а б Olsen, S.J .; Чанг, Й; Мур, П. С .; Biggar, R.J .; Мельбай, М. (1998). «Повышение серологической распространенности герпесвируса, связанного с саркомой Капоши, с возрастом в регионе с высокой степенью эндемичности саркомы Капоши, Замбия, в 1985 году». СПИД. 12 (14): 1921–5. Дои:10.1097/00002030-199814000-00024. PMID 9792393. S2CID 1734745.
- ^ Wawer, M. J .; Eng, S. M .; Serwadda, D; Sewankambo, N.K .; Kiwanuka, N; Ли, С; Грей, Р. Х. (2001). «Распространенность герпесвируса, связанного с саркомой Капоши, по сравнению с отдельными заболеваниями, передаваемыми половым путем, у подростков и молодых людей в сельском районе Ракаи, Уганда». Заболевания, передающиеся половым путем. 28 (2): 77–81. Дои:10.1097/00007435-200102000-00003. PMID 11234789. S2CID 7592869.
- ^ Класкала, З; Brayfield, B.P .; Канкаса, C; Бхат, G; West, J. T .; Mitchell, C.D .; Вуд, С. (2005). «Эпидемиологические характеристики инфекции вируса герпеса-8 человека в большой популяции женщин в дородовой период в Замбии». Журнал медицинской вирусологии. 75 (1): 93–100. Дои:10.1002 / jmv.20242. PMID 15543582.
- ^ Кури, V; Eng, S. M .; Родригес, М. Э .; Resik, S; Оррака, О; Мур, П. С .; Чанг, Y (2004). «Распространенность вируса герпеса, связанного с саркомой Капоши, в различных группах населения Кубы». Revista Panamericana de Salud Pública. 15 (5): 320–5. Дои:10.1590 / с1020-49892004000500006. PMID 15231079.
- ^ Шульц, Т. Ф. (2000). «Вирус герпеса, связанный с саркомой Капоши (вирус герпеса человека 8): эпидемиология и патогенез». Журнал антимикробной химиотерапии. 45 Дополнение T3 (90004): 15–27. Дои:10.1093 / jac / 45.suppl_4.15. PMID 10855768.
- ^ Месри, Энрике А .; Сезарман, Этель; Бошофф, Крис (октябрь 2010 г.). «Вирус герпеса саркомы Капоши / вирус герпеса человека-8 (KSHV / HHV8) и онкогенез саркомы Капоши». Обзоры природы. Рак. 10 (10): 707. Дои:10.1038 / nrc2888. PMID 20865011.
- ^ Brayfield, B.P .; Фири, S; Канкаса, C; Muyanga, J; Mantina, H; Kwenda, G; West, J. T .; Бхат, G; Маркс, Д. Б .; Класкала, З; Mitchell, C.D .; Вуд, С. (2003). «Послеродовая инфекция вируса герпеса человека 8 и вируса иммунодефицита человека типа 1 у матерей и младенцев из Замбии». Журнал инфекционных болезней. 187 (4): 559–68. Дои:10.1086/367985. PMID 12599072.
- ^ Он, Дж; Бхат, G; Канкаса, C; Чинту, C; Митчелл, К; Дуань, Вт; Вуд, С. (1998). «Распространенность вируса герпеса человека 8 среди замбийских женщин детородного возраста без саркомы Капоши (СК) и пар мать-ребенок с СК». Журнал инфекционных болезней. 178 (6): 1787–90. Дои:10.1086/314512. PMID 9815235.
- ^ Минхас, В; Crabtree, K. L .; Чао, А; m'Soka, T. J .; Канкаса, C; Bulterys, M; Mitchell, C.D .; Вуд, С. (2008). «Инфекция в раннем детстве вирусом герпеса человека 8 в Замбии и роль коинфекции вируса иммунодефицита человека типа 1 в высокоэндемичной зоне». Американский журнал эпидемиологии. 168 (3): 311–20. Дои:10.1093 / aje / kwn125. ЧВК 2727264. PMID 18515794.
- ^ Liu Z, Fang Q, Zuo J, Minhas V, Wood C, He N, Zhang T (2017) Был ли вирус герпеса, связанный с саркомой Капоши, занесен в Китай через древний Шелковый путь? Эволюционная перспектива. Arch Virol
- ^ Джэри А., Ледук В., Дезире Н, Пети Х, Палич Р., Джоли В., Канестри А., Готланд А, Ламберт-Никло С., Сургерс Л., Амиэль С., Декамп D, Спано Дж. П., Катлама С., Кальвез В., Марселин АГ ( 2020) Новый вариант герпесвируса Капоши, связанный с саркомой, у мужчин, имеющих половые контакты с мужчинами, связанные с тяжелыми патологиями. J Infect Dis
- ^ Мартин Д.Ф., Купперманн Б.Д., Волиц Р.А., Палестина АГ, Ли Х., Робинсон Калифорния (апрель 1999 г.). «Ганцикловир для перорального применения для пациентов с цитомегаловирусным ретинитом, получавших имплантат ганцикловира. Группа исследования ганцикловира Roche». Медицинский журнал Новой Англии. 340 (14): 1063–70. Дои:10.1056 / NEJM199904083401402. PMID 10194235.
- ^ Yarchoan, R .; Tosato, G .; Литтл, Р. Ф. (2005). «Взгляд на терапию: злокачественные новообразования, связанные со СПИДом - влияние противовирусной терапии на патогенез и лечение». Природа Клиническая Практика Онкологии. 2 (8): 406–415, викторина 415. Дои:10.1038 / ncponc0253. PMID 16130937. S2CID 23476060.
- ^ Бруссард, Грант; Дамания, Цветение (07.02.2020). «КШВ: Иммуномодуляция и иммунотерапия». Границы иммунологии. 10. Дои:10.3389 / fimmu.2019.03084. ISSN 1664-3224. ЧВК 7025529. PMID 32117196.
- ^ Самолс М.А., Ху Дж., Скальский Р.Л., Ренне Р. (июль 2005 г.). «Клонирование и идентификация кластера микроРНК в ассоциированной с латентностью области герпесвируса Капоши, ассоциированного с саркомой». Журнал вирусологии. 79 (14): 9301–5. Дои:10.1128 / JVI.79.14.9301-9305.2005. ЧВК 1168752. PMID 15994824.
дальнейшее чтение
- Эдельман, округ Колумбия (2005). «Вирус герпеса человека 8 - новый патоген для человека». Журнал вирусологии. 2: 78. Дои:10.1186 / 1743-422X-2-78. ЧВК 1243244. PMID 16138925.
внешняя ссылка
- Human Herpesvirus-8: Связанные ресурсы HIV InSite
- Ежегодный семинар КШВ 22-й Международный семинар по герпесвирусу карпози-саркомы (KSHV) и родственным агентам, 30 июня - 3 июля 2019 г., Нью-Йорк, США