Система секреции третьего типа - Type three secretion system
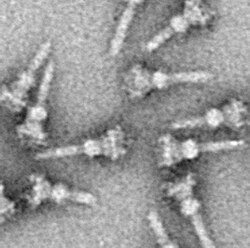
Система секреции третьего типа (часто пишется Система секреции типа III и сокращенно TTSS или же T3SS, также называемый Инъекционный) это белок придаток найден в нескольких Грамотрицательный бактерии.
У патогенных бактерий игольчатая структура используется в качестве сенсорного зонда для обнаружения присутствия эукариотический организмы и выделять белки, которые помогают бактериям заразить их. Секретный эффекторные белки секретируются непосредственно из бактериальных клетка в эукариотическую (хозяйскую) клетку, где они оказывают ряд эффектов, которые помогают патогену выжить и избежать иммунного ответа.
Обзор
Термин «система секреции типа III» был придуман в 1993 году.[1] Эта система секреции отличается как минимум от пяти других системы секреции содержится в грамотрицательных бактериях. Многие бактерии, ассоциированные с животными и растениями, обладают аналогичными T3SS. Эти T3SS схожи в результате дивергентной эволюции, и филогенетический анализ поддерживает модель, в которой грамотрицательные бактерии могут переносить T3SS. генная кассета по горизонтали к другим видам. Наиболее изученные T3SS относятся к видам Шигелла (причины бактериальная дизентерия ), Сальмонелла (брюшной тиф ), кишечная палочка (Кишечная флора, некоторые штаммы вызывают пищевое отравление ), Вибрион (гастроэнтерит и понос ), Burkholderia (сап ), Иерсиния (чума ), Хламидиоз (венерическая болезнь ), Псевдомонады (заражает люди, животные и растения ) и патогены растений Эрвиния, Ralstonia и Ксантомонады, и растительный симбионт Ризобий.
T3SS состоит примерно из 30 различных белков, что делает его одной из самых сложных систем секреции. Его структура имеет много общего с бактериальным жгутики (длинные жесткие внеклеточные структуры, используемые для подвижность ). Некоторые из белков, участвующих в T3SS, разделяют аминокислота гомология последовательностей жгутиковых белков. Некоторые из бактерий, обладающих T3SS, также имеют жгутики и подвижны (Сальмонелла, например), а некоторые нет (Шигелла, например). С технической точки зрения, секреция типа III используется как для секреции белков, связанных с инфекцией, так и для жгутиковых компонентов. Однако термин «секреция типа III» используется в основном в отношении инфекционного аппарата. Жгутик бактерий имеет общего предка с системой секреции типа III.[2][3]
T3SS необходимы для патогенности (способности инфицировать) многих патогенных бактерий. Дефекты T3SS могут сделать бактерию непатогенной. Было высказано предположение, что некоторые неинвазивные штаммы грамотрицательных бактерий потеряли T3SS, потому что энергетически дорогостоящая система больше не используется.[4] Хотя традиционный антибиотики были эффективны против этих бактерий в прошлом, устойчивый к антибиотикам постоянно возникают штаммы. Понимание того, как работает T3SS, и разработка лекарств, нацеленных на него, стали важной целью многих исследовательских групп по всему миру с конца 1990-х годов.
Структура
| Система секреции типа III | |
|---|---|
 Игольчатый комплекс T3SS | |
| Идентификаторы | |
| Символ | T3SS |
| TCDB | 1.B.22 |
| OPM суперсемейство | 348 |
| Белок OPM | 5ткк |
Отличительная черта T3SS - игла.[5][6] (в более общем смысле игольчатый комплекс (NC) или Аппарат T3SS (T3SA); также называемый инъекционный когда АТФаза исключен; Смотри ниже). Бактериальные белки, которые необходимо секретировать, проходят от бактериального цитоплазма через иглу прямо в цитоплазму хозяина. Три мембраны разделяют две цитоплазмы: двойную мембрану (внутреннюю и внешнюю) грамотрицательных бактерий и эукариотическую мембрану. Игла обеспечивает плавный проход через эти высокоселективные и почти непроницаемые мембраны. У одной бактерии может быть несколько сотен игольчатых комплексов, распределенных по ее мембране. Было высказано предположение, что комплекс иглы является универсальным признаком всех T3SS патогенных бактерий.[7]
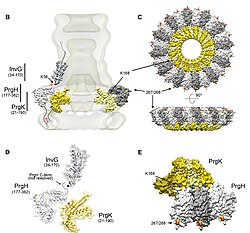
Комплекс иглы начинается в цитоплазме бактерии, пересекает две мембраны и выходит из клетки. Часть, закрепленная в мембране, является основание (или же базальное тело) из T3SS. Внеклеточная часть - игла. Так называемый внутренний стержень соединяет иглу с основанием. Сама игла, хотя и является самой большой и наиболее заметной частью T3SS, состоит из множества единиц одного белка. Таким образом, большинство различных белков T3SS - это те, которые образуют основу, и те, которые секретируются в хозяина. Как упоминалось выше, комплекс игл имеет сходство с бактериальными жгутиками. Более конкретно, основание комплекса игл структурно очень похоже на основание жгутика; сама игла аналогична крючку жгутика, структуре, соединяющей основание с нитью жгутика.[8][9]
Основание состоит из нескольких круглых колец и является первой структурой, построенной в новом комплексе игл. Когда основа готова, она служит машиной для секреции внешних белков (иглы). Когда весь комплекс завершен, система переключается на секретирование белков, которые предназначены для доставки в клетки-хозяева. Предполагается, что игла построена снизу вверх; единицы иглы мономер белковые груды друг на друга, так что единица на кончике иглы добавляется последней. Субъединица иглы - один из самых маленьких белков T3SS, его длина составляет около 9 тыс.Да. Каждая игла состоит из 100-150 субъединиц.
Игла T3SS измеряет около 60-80 нм по длине и 8 нм по внешней ширине. Он должен иметь минимальную длину, чтобы другие внеклеточные бактериальные структуры (адгезины и липополисахарид слой, например) не препятствуют секреции. Отверстие иглы имеет диаметр 3 нм. Большинство свернутых эффекторных белков слишком велики, чтобы пройти через отверстие иглы, поэтому большая часть секретируемых белков должна проходить через иглу. развернутый, задача, выполняемая АТФаза в основании конструкции.[10]
Белки T3SS

Белки T3SS можно разделить на три категории:
- Структурные белки: построить основу, внутренний стержень и иглу.
- Эффекторные белки: секретируются в клетку-хозяин и способствуют инфицированию / подавлению защиты клетки-хозяина.
- Шапероны: связывают эффекторы в цитоплазме бактерий, защищают их от агрегации и деградация и направить их к комплексу игл.
Большинство генов T3SS расположены в опероны. Эти опероны расположены на бактериальной хромосоме у некоторых видов и на выделенном участке. плазмида у других видов. Сальмонелла, например, имеет хромосомную область, в которой собрано большинство генов T3SS, так называемые Сальмонелла остров патогенности (SPI). Шигелла, с другой стороны, имеет большую плазмиду вирулентности, на которой расположены все гены T3SS. Важно отметить, что многие островки патогенности и плазмиды содержат элементы, которые позволяют частый горизонтальный перенос генов острова / плазмиды новому виду.
Эффекторные белки, которые должны секретироваться через иглу, должны распознаваться системой, поскольку они плавают в цитоплазме вместе с тысячами других белков. Признание осуществляется через сигнал секреции- короткая последовательность аминокислот, расположенная в начале ( N-конец ) белка (обычно в пределах первых 20 аминокислот), который комплекс иглы способен распознать. В отличие от других систем секреции, сигнал секреции белков T3SS никогда не отщепляется от белка.
Индукция секреции
Контакт иглы с клеткой-хозяином запускает секрецию T3SS;[11] Об этом спусковом механизме известно немного (см. ниже). Секреция также может быть вызвана снижением концентрации кальций ионы в среда роста (за Иерсиния и Псевдомонады; сделано путем добавления хелатор Такие как EDTA или же EGTA ) и добавив ароматный краситель Конго красный к питательной среде (для Шигелла), например. Эти и другие методы используются в лабораториях для искусственного стимулирования секреции III типа.
Также имеет место индукция секреции внешними сигналами, кроме контакта с клетками-хозяевами. in vivo в инфицированных организмах. Бактерии улавливают такие сигналы, как температура, pH, осмолярность и кислород уровни, и используйте их, чтобы «решить», активировать ли их T3SS. Например, Сальмонелла может лучше воспроизводиться и вторгаться в подвздошная кишка а не в слепая кишка животных кишечник. Бактерии могут знать, где они находятся, благодаря различным ионам, присутствующим в этих областях; подвздошная кишка содержит форматировать и ацетат, а слепая кишка - нет. Бактерии ощущают эти молекулы, определяют, что они находятся в подвздошной кишке, и активируют свой секреторный аппарат. Молекулы, присутствующие в слепой кишке, такие как пропионат и бутират, являются негативным сигналом для бактерий и подавляют секрецию. Холестерин, а липид обнаружен в мембранах большинства эукариотических клеток, способен вызывать секрецию Шигелла.
Перечисленные выше внешние сигналы регулируют секрецию либо напрямую, либо через генетический механизм. Несколько факторы транскрипции которые регулируют выражение генов T3SS известны. Некоторые шапероны, связывающие эффекторы T3SS, также действуют как факторы транскрипции. Был предложен механизм обратной связи: когда бактерия не секретирует, ее эффекторные белки связываются с шаперонами и плавают в цитоплазме. Когда начинается секреция, шапероны отделяются от эффекторов, и последние секретируются и покидают клетку. Затем одиночные шапероны действуют как факторы транскрипции, связываясь с генами, кодирующими их эффекторы, и индуцируют их транскрипцию и тем самым производство большего количества эффекторов.
Структуры, аналогичные инъекциомам Type3SS, были предложены для заклепки внешней и внутренней мембран грамотрицательных бактерий, чтобы помочь высвободить везикулы внешней мембраны, нацеленные на доставку бактериального секрета к эукариотическим клеткам-хозяевам или другим клеткам-мишеням in vivo.[12]
T3SS-опосредованная инфекция
Эффекторы T3SS входят в комплекс иглы в основании и продвигаются внутрь иглы к клетке-хозяину. Точный путь проникновения эффекторов в организм хозяина в основном неизвестен. Ранее предполагалось, что игла сама по себе способна пробить отверстие в мембране клетки-хозяина; эта теория была опровергнута. Теперь ясно, что некоторые эффекторы, названные вместе транслокаторы, секретируются первыми и образуют пору или канал ( Translocon) в мембране клетки-хозяина, через которую могут проникать другие эффекторы. Мутировавший бактерии, у которых отсутствуют транслокаторы, способны секретировать белки, но не могут доставлять их в клетки-хозяева. Обычно каждый T3SS включает три транслокатора. Некоторые транслокаторы выполняют двойную роль; после того, как они участвуют в порообразовании, они попадают в клетку и действуют как добросовестный эффекторы.
Эффекторы T3SS манипулируют клетками-хозяевами несколькими способами. Самый поразительный эффект - это стимулирование поглощения бактерии клеткой-хозяином. Многие бактерии, обладающие T3SS, должны проникать в клетки-хозяева, чтобы реплицировать и распространять инфекцию. Эффекторы, которые они вводят в клетку-хозяин, побуждают хозяина поглотить бактерию и практически «съесть» ее. Для этого бактериальные эффекторы воздействуют на актин полимеризация машины клетки-хозяина. Актин входит в состав цитоскелет он также участвует в подвижности и изменении формы клеток. Благодаря своим эффекторам T3SS бактерия может использовать собственный механизм клетки-хозяина для собственной выгоды. После того, как бактерия попала в клетку, она способна более легко секретировать другие эффекторы, может проникать в соседние клетки и быстро инфицировать всю клетку. ткань.
Также было показано, что эффекторы T3SS влияют на работу хозяина. клеточный цикл и некоторые из них способны вызвать апоптоз. Одним из наиболее исследованных эффекторов T3SS является IpaB из Шигелла флекснери. Он выполняет двойную роль как транслокатор, создавая поры в мембране клетки-хозяина, и как эффектор, оказывая множественные пагубные эффекты на клетку-хозяина. Было продемонстрировано, что IpaB вызывает апоптоз у макрофаги —Клетки животного иммунная система - после того, как их поглотили.[13] Позже было показано, что IpaB достигает этого, взаимодействуя с каспаза 1, главный регуляторный белок в эукариотических клетках.[14]
Другой хорошо охарактеризованный класс эффекторов T3SS - это эффекторы, подобные активаторам транскрипции (TAL эффекторы ) из Ксантомонады. При введении в растения эти белки могут проникать в ядро растительной клетки, связывать промоторные последовательности растений и активировать транскрипцию генов растений, которые способствуют бактериальной инфекции.[15] Недавно было продемонстрировано, что распознавание эффекторной ДНК TAL состоит из простого кода.[16][17] и это значительно улучшило понимание того, как эти белки могут изменять транскрипцию генов в клетках растения-хозяина.
Нерешенные вопросы
Сотни статей о T3SS были опубликованы с середины девяностых годов. Однако многие вопросы, касающиеся системы, остаются нерешенными:
- Белки T3SS. Из примерно 30 белков T3SS менее 10 в каждом организме были непосредственно обнаружены с помощью биохимический методы. Остальные, возможно, редкие, оказалось трудно обнаружить, и они остаются теоретическими (хотя генетические, а не биохимические исследования были выполнены на многих генах / белках T3SS). Локализация каждого белка также полностью не известна.
- Длина иглы. Неизвестно, как бактерия «узнает», когда новая игла достигла нужной длины. Существует несколько теорий, в том числе о существовании «протеина-линейки», который каким-то образом соединяет кончик и основание иглы. Добавление новых мономеров на кончик иглы должно растягивать белок линейки и тем самым сигнализировать длину иглы до основания.
- Энергетика. Сила, которая вызывает прохождение белков внутри иглы, полностью не известна. An АТФаза связан с основанием T3SS и участвует в направлении белков в иглу; но неясно, поставляет ли он энергию для транспорта.
- Сигнал секреции. Как упоминалось выше, существование сигнала секреции у эффекторных белков известно. Сигнал позволяет системе отличить T3SS-транспортируемые белки от любых других белков. Его природа, требования и механизм распознавания плохо изучены, но недавно были разработаны методы прогнозирования того, какие бактериальные белки могут транспортироваться системой секреции типа III.[19]
- Активация секреции. Бактерия должна знать, когда пора выделять эффекторы. Излишняя секреция, когда поблизости нет клетки-хозяина, расточительна для бактерии с точки зрения энергии и ресурсов. Бактерия каким-то образом способна распознавать контакт иглы с клеткой-хозяином. Как это делается, все еще исследуются, и этот метод вполне может зависеть от патогена. Некоторые теории постулируют тонкое конформационное изменение структуры иглы при контакте с клеткой-хозяином; это изменение, возможно, служит сигналом для начала секреции. Один метод распознавания был открыт в Сальмонелла, который основан на ощущении цитозольного pH через остров патогенности 2, кодируемый T3SS, чтобы включить секрецию эффекторов.[20]
- Привязка шаперонов. Неизвестно, когда шапероны связывают свои эффекторы (во время или после перевод ) и как они отделяются от своих эффекторов перед секрецией.
- Эффекторные механизмы. Хотя с начала 21 века многое было раскрыто о способах, которыми эффекторы T3SS манипулируют хозяином, большинство эффектов и путей остается неизвестным.
- Эволюция. Как уже упоминалось, T3SS тесно связан с жгутиком бактерий.[21] Есть три конкурирующие гипотезы:[22] во-первых, что жгутик развился первым и T3SS произошел от этой структуры, во-вторых, что T3SS развился первым и жгутик произошел от него, и в-третьих, что две структуры произошли от общего предка. Были некоторые разногласия по поводу различных сценариев,[2][22] поскольку все они объясняют гомологию белков между двумя структурами, а также их функциональное разнообразие.[23] Тем не менее, недавние филогеномные данные подтверждают гипотезу, что T3SS происходит из жгутика в результате процесса, включающего первоначальную потерю гена, а затем приобретение гена.[24] Ключевым этапом последнего процесса было рекрутирование секретинов в T3SS, событие, которое происходило по крайней мере три раза из других мембраносвязанных систем.
Номенклатура белков T3SS

С начала 1990-х годов новые белки T3SS стабильно обнаруживаются у разных видов бактерий. Аббревиатуры были даны независимо для каждой серии белков в каждом организме, и названия обычно мало что говорят о функции белка. Некоторые белки, независимо открытые в разных бактериях, позже показали, что они гомологичный; исторические названия, однако, в основном были сохранены, что может вызвать путаницу. Например, белки SicA, IpgC и SycD являются гомологами из Сальмонелла, Шигелла и Иерсиниясоответственно, но последняя буква («порядковый номер») в их имени этого не показывает.
Ниже приводится сводка наиболее распространенных названий серий белков у нескольких видов, содержащих T3SS. Обратите внимание, что эти названия включают белки, которые формируют механизм T3SS, а также секретируемые эффекторные белки:
- Иерсиния
- Йоп: Иерсиния внешний белок
- Ysc: Иерсиния секреция (компонент)
- Ypk: Иерсиния протеинкиназа
- Сальмонелла
- спа: Поверхностное представление антигена
- Sic: Сальмонелла компаньон вторжения
- Глоток: Сальмонелла белок инвазии
- Prg: PhoP-репрессированный ген
- Инв.: Вторжение
- Org: Регулируемый кислородом ген
- Ssp: Сальмонелла-секретированный белок
- Iag: Ген, связанный с инвазией
- Шигелла
- IPG: Ген плазмиды вторжения
- Ипа: Антиген плазмиды инвазии
- Mxi: Мембранная экспрессия Ipa
- спа: Поверхностное представление антигена
- Osp: Внешний Шигелла белок
- Эшерихия
- Тир: Транслоцированный рецептор интимина
- Сен: Секрет Кишечная палочка белки
- Esc: Эшерихия секреция (компонент)
- Esp: Эшерихия белок секрета
- Ces: Шаперона Кишечная палочка секреция
- Псевдомонады
- Hrp: Гиперчувствительность и патогенность.
- Hrc: Гиперчувствительный ответ сохранен (или Hrp сохранен)
- Ризобий
- Нет: Нодулирующий белок
- Rhc: Ризобий консервированный
- У нескольких видов:
- Вир: Вирулентность
- «Протохламидия амебофила»
- "Sodalis glossinidius"[25]
После этих сокращений следует буква или число. Буквы обычно обозначают «порядковый номер», либо хронологический порядок открытия, либо физический порядок появления гена в оперон. Цифры, реже, обозначают молекулярную массу белка в кДа. Примеры: IpaA, IpaB, IpaC; MxiH, MxiG, MxiM; Спа9, Спа47.
Во всех T3SS присутствуют несколько ключевых элементов: мономер иглы, внутренний стержень иглы, кольцевые белки, два транслокатора, белок кончика иглы, белок-линейка (который, как считается, определяет длину иглы; см. Выше) и в АТФаза, который поставляет энергию для секреции. В следующей таблице показаны некоторые из этих ключевых белков у четырех бактерий, содержащих T3SS:
| ↓ Функция / Род → | Шигелла | Сальмонелла | Иерсиния | Эшерихия |
|---|---|---|---|---|
| Мономер иглы | MxiH | PrgI | YscF | EscF |
| Внутренний стержень | MxiI | PrgJ | YscI | EscI |
| Белок кончика иглы | IpaD | SipD | LcrV | EspA |
| Транслокатор | IpaB | SipB | YopB | EspD |
| Транслокатор | IpaC | SipC | YopD | EspB |
| Шаперон для двух транслокаторов | IpgC | SicA | SycD | CesD |
| АТФаза | Спа47 | InvC | YscN | SepB (EscN) |
| Линейка белка | Спа32 | InvJ | YscP | Orf16 |
| Выключатель | Спа40 | SpaS | YscU | EscU |
| Привратник | MxiC | InvE | YopN (TyeA) | SepL |
Методы, использованные в исследовании T3SS
Выделение игольчатых комплексов T3SS
Изоляция больших, хрупких, гидрофобный мембранные структуры из клеток были проблемой в течение многих лет. Однако к концу 1990-х годов было разработано несколько подходов к изоляции T3SS NC. В 1998 г. были выделены первые НК из Сальмонелла тифимуриум.[26]
Для изоляции бактерии выращивают в большом объеме жидкости. среда роста пока они не достигнут этап журнала. Они тогда центрифугированный; то супернатант (среда) отбрасывается, а гранула (бактерии) ресуспендируют в буфер для лизиса обычно содержащий лизоцим а иногда моющее средство Такие как LDAO или же Тритон Х-100. Этот буфер разрушает клеточная стенка. После нескольких циклов лизиса и отмывки открытые бактерии подвергаются серии ультрацентрифугирование. Эта обработка обогащает крупные макромолекулярные структуры и удаляет более мелкие клеточные компоненты. Необязательно, конечный лизат подвергают дальнейшей очистке путем CsCl градиент плотности.
Дополнительный подход для использования в дальнейшей очистке аффинная хроматография. Рекомбинантный Белки T3SS, несущие белковая метка (а гистидиновый тег, например) производятся молекулярное клонирование а затем представил (преобразованный ) в исследуемые бактерии. После первоначального выделения NC, как описано выше, лизат пропускают через колонку, покрытую частицами с высоким сродством к метке (в случае гистидиновых меток: никель ионы ). Меченый белок остается в колонке, а вместе с ним и весь комплекс иглы. С помощью таких методов можно достичь высокой степени чистоты. Эта чистота важна для многих деликатных анализов, которые использовались для характеристики NC.
Эффекторы типа III были известны с начала 1990-х годов, но то, как они доставляются в клетки-хозяева, оставалось полной загадкой. Гомология между многими жгутиковый и белки T3SS заставили исследователей подозревать существование внешней структуры T3SS, подобной жгутикам. Идентификация и последующее выделение структуры иглы позволило исследователям:
- подробно охарактеризовать трехмерную структуру NC и на основании этого сделать выводы относительно механизма секреции (например, что узкая ширина иглы требует развертывания эффекторов перед секрецией),
- анализировать белковые компоненты NC, подвергая изолированные иглы протеомному анализу (см. ниже),
- назначать роли различным компонентам ЧПУ, выбивание Гены T3SS, выделение NC из мутировавших бактерий и изучение изменений, вызванных мутациями.
Микроскопия, кристаллография и твердотельный ЯМР
Как и почти все белки, визуализация NC T3SS возможна только с электронная микроскопия. Первые изображения NC (1998) показали игольчатые структуры, выступающие из клеточной стенки живых бактерий и плоские, двумерные изолированные NC.[26] В 2001 г. изображения СК из Шигелла флекснери были подвергнуты цифровому анализу и усреднены для получения первой полу-3D-структуры NC.[5] Винтовая структура НК из Шигелла флекснери был разрешен с разрешением 16 Å с помощью рентгеновский снимок дифракция волокна в 2003 г.,[27] а через год 17-Å 3D-структура ЧПУ из Сальмонелла тифимуриум был опубликован.[28] Последние достижения и подходы позволили получить трехмерные изображения СК с высоким разрешением,[29][30] дальнейшее уточнение сложной структуры СК.
Многочисленные белки T3SS были кристаллизованы на протяжении многих лет. К ним относятся структурные белки NC, эффекторы и шапероны. Первой структурой игольчатого сложного мономера была структура ЯМР BsaL из "Burkholderia pseudomallei", а затем кристаллическая структура MixH из Шигелла флекснери, которые были разрешены в 2006 году.[31][32]
В 2012 году комбинация производства рекомбинантных игл дикого типа, твердотельный ЯМР, электронная микроскопия[33] и моделирование Розетты выявило супрамолекулярные границы раздела и, в конечном итоге, полную атомную структуру Сальмонелла тифимуриум Игла T3SS.[34] Было показано, что субъединицы PrgI с 80 остатками образуют правую спиральную сборку с примерно 11 субъединицами на два витка, аналогично таковой у субъединиц PrgI. жгутик из Сальмонелла тифимуриум. Модель также выявила протяженный аминоконцевой домен, который расположен на поверхности иглы, в то время как высококонсервативный карбокси-конец указывает на просвет.[34]
Протеомика
Несколько методов были использованы для идентификации массива белков, составляющих T3SS. Изолированные игольчатые комплексы можно разделить с помощью SDS-СТРАНИЦА. Полосы, которые появляются после окрашивания, можно индивидуально вырезать из геля и проанализировать с помощью секвенирование белков и масс-спектрометрии. Структурные компоненты NC могут быть отделены друг от друга (например, игольчатая часть от основной части), и путем анализа этих фракций можно определить белки, участвующие в каждой из них. В качестве альтернативы изолированные НК можно непосредственно анализировать масс-спектрометрией без предварительного электрофорез, чтобы получить полное представление о НК протеом.
Генетические и функциональные исследования
Исследователи манипулировали T3SS у многих бактерий. Наблюдение за влиянием отдельных манипуляций может быть использовано для понимания роли каждого компонента системы. Примеры манипуляций:
- Удаление одного или нескольких генов T3SS (нокаут гена ).
- Сверхэкспрессия одного или нескольких генов T3SS (другими словами: производство in vivo белка T3SS в количествах больше, чем обычно).
- Точечные или региональные изменения в генах или белках T3SS. Это делается для того, чтобы определить функцию определенных аминокислот или участков в белке.
- Введение гена или белка из одного вида бактерий в другой (анализ перекрестной комплементации). Это делается для того, чтобы проверить различия и сходства между двумя T3SS.
Манипуляции с компонентами T3SS могут влиять на несколько аспектов бактериальной функции и патогенности. Примеры возможных влияний:
- Способность бактерий проникать в клетки-хозяева в случае внутриклеточных патогенов. Это можно измерить с помощью анализ вторжения (анализ защиты от гентамицина ).
- Способность внутриклеточных бактерий мигрировать между клетками-хозяевами.
- Способность бактерий убивать клетки-хозяева. Это можно измерить несколькими методами, например LDH -выпускной анализ, в котором фермент ЛДГ, который вытекает из мертвых клеток, определяется путем измерения его ферментативной активности.
- Способность T3SS секретировать определенный белок или вообще секретировать. Чтобы проверить это, у бактерий, растущих в жидкой среде, индуцируется секреция. Затем бактерии и среда разделяются центрифугированием, и фракция среды (супернатант) затем анализируется на наличие секретируемых белков. Чтобы предотвратить секрецию обычно секретируемого белка, к нему можно искусственно присоединить большую молекулу. Если тогда не секретируемый белок остается «застрявшим» на дне комплекса иглы, секреция эффективно блокируется.
- Способность бактерий собирать неповрежденный игольчатый комплекс. NC могут быть выделены из обработанных бактерий и исследованы под микроскопом. Однако незначительные изменения не всегда можно обнаружить с помощью микроскопии.
- Способность бактерий заражать живых животных или растений. Даже если показаны управляемые бактерии in vitro Чтобы иметь возможность инфицировать клетки-хозяева, их способность поддерживать инфекцию в живом организме не может считаться само собой разумеющимся.
- Уровни экспрессии других генов. Это можно оценить несколькими способами, в частности северное пятно и ОТ-ПЦР. Уровни экспрессии всего геном может быть проанализирован микрочип. Многие типа III факторы транскрипции и регуляторные сети были обнаружены с использованием этих методов.
- Рост и приспособленность бактерий.
Ингибиторы T3SS
Было обнаружено несколько соединений, которые ингибируют T3SS в грамотрицательные бактерии, в том числе гуадиномины которые естественным образом производятся Streptomyces разновидность.[35] Моноклональные антитела были разработаны, которые также подавляют T3SS.[36]
Инструменты для прогнозирования сигнального пептида типа III
Рекомендации
- ^ Салмонд Г.П., Ривз П.Дж. (1993). «Управляющие мембранным движением и секреция белка у грамотрицательных бактерий». Тенденции в биохимических науках. 18 (1): 7–12. Дои:10.1016/0968-0004(93)90080-7. PMID 8438237.
- ^ а б Гофна Ю., Рон Э.З., Граур Д. (июль 2003 г.). «Системы секреции бактерий типа III являются древними и развивались в результате множественных событий горизонтального переноса». Ген. 312: 151–63. Дои:10.1016 / S0378-1119 (03) 00612-7. PMID 12909351.
- ^ Нгуен Л., Полсен И.Т., Чьеу Дж., Хьюк С.Дж., Сайер М.Х. (апрель 2000 г.). «Филогенетический анализ составляющих систем секреции белка III типа». Журнал молекулярной микробиологии и биотехнологии. 2 (2): 125–44. PMID 10939240.
- ^ Gong H, Vu GP, Bai Y, Yang E, Liu F, Lu S. (январь 2010 г.). «Дифференциальная экспрессия факторов системы секреции сальмонеллы типа III InvJ, PrgJ, SipC, SipD, SopA и SopB в культурах и у мышей». Микробиология. 156 (Pt 1): 116–27. Дои:10.1099 / микрофон.0.032318-0. ЧВК 2889428. PMID 19762438.
- ^ а б Blocker A, Jouihri N, Larquet E, Gounon P, Ebel F, Parsot C, Sansonetti P, Allaoui A (2001). «Структура и состав« игольчатого комплекса »Shigella flexneri, часть его секретона III типа». Мол Микробиол. 39 (3): 652–663. Дои:10.1046 / j.1365-2958.2001.02200.x. PMID 11169106.
- ^ Галан Дж. Э., Вольф-Ватц Х (2006). «Доставка белка в эукариотические клетки с помощью машин секреции типа III». Природа. 444 (7119): 567–573. Bibcode:2006Натура 444..567Г. Дои:10.1038 / природа05272. PMID 17136086.
- ^ Паллен М. Дж .; Bailey C.M .; Битсон С. А. (2006). «Эволюционные связи между Flih / Yscl-подобными белками из бактериальных систем секреции III типа и компонентами второго стебля FoF1 и вакуолярных АТФаз». Белковая наука. 15 (4): 935–940. Дои:10.1110 / пс. 051958806. ЧВК 2242474. PMID 16522800.
- ^ Айзава С (2001). «Бактериальные жгутики и системы секреции III типа». Письма о микробиологии FEMS. 202 (2): 157–164. Дои:10.1111 / j.1574-6968.2001.tb10797.x. PMID 11520608.
- ^ Дулиттл В. Ф .; Жакыбаева, Ольга (2007). «Эволюция: сводимая сложность - случай бактериальных жгутиков». Текущая биология. 17 (13): R510–512. Дои:10.1016 / j.cub.2007.05.003. PMID 17610831.
- ^ Акеда Й., Галан Дж. Э. (октябрь 2005 г.). «Высвобождение шаперона и развертывание субстратов в секреции III типа». Природа. 437 (7060): 911–5. Bibcode:2005Натура.437..911A. Дои:10.1038 / природа03992. PMID 16208377.
- ^ Kimbrough T. G .; Миллер С. И. (2000). «Вклад компонентов секреции Salmonella typhimurium type iii в формирование комплекса игл». Труды Национальной академии наук Соединенных Штатов Америки. 97 (20): 11008–11013. Bibcode:2000PNAS ... 9711008K. Дои:10.1073 / pnas.200209497. ЧВК 27139. PMID 10984518.
- ^ Яшрой Р.С. (2003). «Интоксикация эукариотических клеток грамотрицательными патогенами: новая модель бактериального нановезикулярного экзоцитоза, связанного с внешней мембраной, для системы секреции типа III». Международная токсикология. 10 (1): 1–9.
- ^ Зихлинский А., Кенни Б., Менард Р., Прево М.С., Голландия И.Б., Сансонетти П.Дж. (1994). «IpaB опосредует апоптоз макрофагов, индуцированный Shigella flexneri». Мол Микробиол. 11 (4): 619–627. Дои:10.1111 / j.1365-2958.1994.tb00341.x. PMID 8196540.
- ^ Хилби Х., Мосс Дж. Э., Херш Д., Чен Й., Арондел Дж., Банерджи С., Флавелл Р. А., Юань Дж., Сансонетти П. Дж., Зихлински А. (1998). «Апоптоз, индуцированный шигеллами, зависит от каспазы-1, которая связывается с IpaB». J Biol Chem. 273 (49): 32895–32900. Дои:10.1074 / jbc.273.49.32895. PMID 9830039.
- ^ Boch, J .; Бонас, У. (2010). «Эффекторы семейства XanthomonasAvrBs3 типа III: открытие и функция». Ежегодный обзор фитопатологии. 48: 419–436. Дои:10.1146 / annurev-phyto-080508-081936. PMID 19400638.
- ^ Moscou, M.J .; Богданове, А. Дж. (2009). «Простой шифр управляет распознаванием ДНК с помощью TAL Effectors». Наука. 326 (5959): 1501. Bibcode:2009Научный ... 326.1501M. Дои:10.1126 / science.1178817. PMID 19933106.
- ^ Бох Дж., Шольце Х., Шорнак С. и др. (Декабрь 2009 г.). «Нарушение кода специфичности связывания ДНК эффекторов TAL-типа III». Наука. 326 (5959): 1509–12. Bibcode:2009Sci ... 326.1509B. Дои:10.1126 / science.1178811. PMID 19933107.
- ^ Schraidt, O .; Lefebre, M.D .; Brunner, M. J .; Schmied, W. H .; Schmidt, A .; Radics, J .; Mechtler, K .; Galán, J. E .; Марловиц, Т. К. (2010). Стеббинс, К. Эрек (ред.). «Топология и организация компонентов комплекса игл для секреции Salmonella typhimurium III типа». Патогены PLOS. 6 (4): e1000824. Дои:10.1371 / journal.ppat.1000824. ЧВК 2848554. PMID 20368966.
- ^ Гринберг М., Годзик А. (апрель 2009 г.). Стеббинс, К. Эрек (ред.). «Сигнал для сигнализации, обнаружен». PLOS Pathog. 5 (4): e1000398. Дои:10.1371 / journal.ppat.1000398. ЧВК 2668190. PMID 19390616.
- ^ Ю. XJ и др. (Май 2010 г.). «Определение pH внутриклеточной сальмонеллой вызывает эффекторную транслокацию». Наука. 328 (5981): 1040–3. Bibcode:2010Sci ... 328.1040Y. Дои:10.1126 / science.1189000. HDL:10044/1/19679. ЧВК 6485629. PMID 20395475.
- ^ Медини Д., Коваччи А., Донати С. (декабрь 2006 г.). «Семейства сетей гомологии белков демонстрируют постепенную диверсификацию систем секреции типа III и типа IV». PLOS Comput. Биол. 2 (12): e173. Bibcode:2006PLSCB ... 2..173M. Дои:10.1371 / journal.pcbi.0020173. ЧВК 1676029. PMID 17140285.
- ^ а б Сайер, М. (2004). «Эволюция бактериальных систем секреции белка III типа». Тенденции в микробиологии. 12 (3): 113–115. Дои:10.1016 / j.tim.2004.01.003. PMID 15001186.
- ^ Макканн ХК, Гутман Д.С. (2008). «Эволюция системы секреции типа III и ее эффекторов в растительно-микробных взаимодействиях». Новый Фитол. 177 (1): 33–47. Дои:10.1111 / j.1469-8137.2007.02293.x. PMID 18078471.
- ^ Эбби, Софи С .; Роча, Эдуардо П. К. (01.09.2012). «Не жгутиковая система секреции типа III произошла от бактериального жгутика и превратилась в адаптированные системы клеток-хозяев». PLOS Genetics. 8 (9): e1002983. Дои:10.1371 / journal.pgen.1002983. ISSN 1553-7404. ЧВК 3459982. PMID 23028376.
- ^ Моран, Н. А. (13 февраля 2001 г.). «Бактериальные зверинцы внутри насекомых». Труды Национальной академии наук. 98 (4): 1338–1340. Дои:10.1073 / пнас.98.4.1338.
- ^ а б Кубори Т., Мацусима Ю., Накамура Д., Уралил Дж., Лара-Теджеро М., Сухан А., Галан Дж. Э., Айзава С. И. (апрель 1998 г.). «Супрамолекулярная структура системы секреции белка Salmonella typhimurium III типа». Наука. 280 (5363): 602–5. Bibcode:1998Научный ... 280..602K. Дои:10.1126 / science.280.5363.602. PMID 9554854.
- ^ Кордес Ф.С., Комория К., Ларке Э., Ян С., Эгельман Э. Х., Блокатор А, Леа С. М. (2003). «Спиральное строение иглы системы секреции III типа Shigella flexneri». J Biol Chem. 278 (19): 17103–17107. Дои:10.1074 / jbc.M300091200. PMID 12571230.
- ^ Марловиц Т.С., Кубори Т., Сухан А., Томас Д.Р., Галан Дж.Э., Унгер В.М. (2004). «Структурные сведения о сборке комплекса игл для секрета III типа». Наука. 306 (5698): 1040–1042. Bibcode:2004Наука ... 306.1040M. Дои:10.1126 / science.1102610. ЧВК 1459965. PMID 15528446.
- ^ Сани М., Аллауи А., Фусетти Ф, Oostergetel GT, Кегстра В., Боекема Э. Дж. (2007). «Структурная организация игольчатого комплекса секреционного аппарата типа III Shigella flexneri» (PDF). Микрон. 38 (3): 291–301. Дои:10.1016 / j.micron.2006.04.007. PMID 16920362.
- ^ Ходжкинсон Дж. Л., Хорсли А., Стабат Д., Саймон М., Джонсон С., да Фонсека ПК, Моррис Е. П., Уолл Дж. С., Леа С. М., Блокировщик А. Дж. (2009). «Трехмерная реконструкция трансмембранных областей Shigella T3SS показывает 12-кратную симметрию и новые особенности повсюду». Нат Структ Мол Биол. 16 (5): 477–485. Дои:10.1038 / nsmb.1599. ЧВК 2681179. PMID 19396171.
- ^ Чжан, Л; Ван, Y; Комплектация, WL; Комплектация, WD; Де Гусман, Р. Н. (2 июня 2006 г.). "Структура раствора мономерного BsaL, белка иглы секреции типа III Burkholderia pseudomallei". Журнал молекулярной биологии. 359 (2): 322–30. Дои:10.1016 / j.jmb.2006.03.028. PMID 16631790.
- ^ Дин Дж. Э., Роверси П., Кордес Ф. С., Джонсон С., Кенджал Р., Даниэлл С., Буй Ф., Сбор У. Д., Сбор У. Л., Блокировщик А. Дж., Ли С. М. (2006). «Молекулярная модель иглы системы секреции типа III: значение для восприятия клеток-хозяев». Proc Natl Acad Sci USA. 103 (33): 12529–12533. Bibcode:2006PNAS..10312529D. Дои:10.1073 / pnas.0602689103. ЧВК 1567912. PMID 16888041.
- ^ Галкин В.Е .; Schmied WH; Шрейдт О; Марловиц Т.С. и Эгельман (2010). «Структура иглы системы секреции Salmonella typhimurium типа III показывает отклонение от жгутиковой системы». Дж Мол Биол. 396 (5): 1392–1397. Дои:10.1016 / j.jmb.2010.01.001. ЧВК 2823972. PMID 20060835.
- ^ а б Локет А., Сгуракис Н.Г., Гупта Р., Гиллер К., Ридель Д., Гусманн К., Гризинджер С., Кольбе М., Бейкер Д., Беккер С., Ланге А. (2012). «Атомная модель иглы системы секреции III типа». Природа. 486 (7402): 276–279. Bibcode:2012Натура 486..276л. Дои:10.1038 / природа11079. ЧВК 3598588. PMID 22699623.
- ^ Holmes, T. C .; May, A.E .; Залета-Ривера, К .; Ruby, J. G .; Skewes-Cox, P .; Fischbach, M. A .; Derisi, J. L .; Ивацуки, М .; Mura, S .; Хосла, К. (2012). «Молекулярные исследования биосинтеза гвадиномина: ингибитор системы секреции типа III». Журнал Американского химического общества. 134 (42): 17797–17806. Дои:10.1021 / ja308622d. ЧВК 3483642. PMID 23030602.
- ^ Теуретцбахер У., Пиддок Л.Дж. (июль 2019 г.). «Нетрадиционные антибактериальные терапевтические возможности и проблемы». Клеточный хозяин и микроб. 26 (1): 61–72. Дои:10.1016 / j.chom.2019.06.004. PMID 31295426.
дальнейшее чтение
- Мгновенное понимание описание химического состава инъекционных сомов Королевское химическое общество
- Взаимодействие хозяин-патоген в Pseudomonas syringae pv. помидор и томат, ведущий к бактериальной пятнистой болезни.