Регуляторные Т-клетки - Regulatory T cell
В регуляторные Т-клетки (Tregs /ˈтярɛɡ/ или Tрег ячейки), ранее известная как супрессорные Т-клетки, являются субпопуляцией Т-клетки которые модулируют иммунная система, поддерживать толерантность к аутоантигенам, и предотвратить аутоиммунное заболевание. Треги иммунодепрессивный и вообще подавить или подавлять индукция и распространение эффекторные Т-клетки.[1] Tregs экспрессируют биомаркеры CD4, FOXP3, и CD25 и считаются производными от того же родословная как наивно Клетки CD4.[2] Поскольку эффекторные Т-клетки также экспрессируют CD4 и CD25, Treg очень сложно эффективно отличить от эффекторных CD4 +, что затрудняет их изучение. Недавние исследования показали, что цитокин TGFβ необходим для дифференциации Treg от наивных CD4 + клеток и важен для поддержания Treg. гомеостаз.[3]
Мышь модели предположили, что модуляция Tregs может лечить аутоиммунные заболевания и рак и может облегчить трансплантация органов[4] и лечение раны.[5] Их последствия для рака сложны. Tregs, как правило, активируются у людей с онкологическими заболеваниями, и, по-видимому, они задействованы во многих местах. опухоли. Исследования как на людях, так и на животных моделях показали, что высокое количество Treg в микроокружении опухоли указывает на плохое прогноз, и считается, что Tregs подавляют иммунитет к опухоли, препятствуя врожденной способности организма контролировать рост раковых клеток.[6] Недавний иммунотерапия Исследования изучают, как регулирование Т-клеток может быть использовано в лечении рака.[7]
Населения
Т-регуляторные клетки являются компонентом иммунной системы, подавляющим иммунные ответы других клеток. Это важная «самопроверка», встроенная в иммунную систему для предотвращения чрезмерных реакций. Регуляторные Т-клетки бывают разных форм, из которых наиболее хорошо изучены те, которые экспрессируют CD4, CD25 и FOXP3 (CD4 + CD25 + регуляторные Т-клетки). Эти «трэги» отличаются от хелперные Т-клетки.[8] Другой подгруппой регуляторных Т-клеток является Клетки Treg17.[9] Регуляторные Т-клетки участвуют в отключении иммунных ответов после того, как они успешно уничтожили вторгшиеся организмы, а также в предотвращении аутоиммунитета.[10]
CD4 + Foxp3 + CD25 (высокий) регуляторные Т-клетки были названы "естественными" регуляторными Т-клетками.[11] чтобы отличить их от популяций «супрессоров» Т-клеток, которые генерируются in vitro. Дополнительные популяции регуляторных Т-клеток включают: Tr1, Th3, CD8 + CD28- и Qa-1 рестриктированные Т-клетки. Вклад этих популяций в самотолерантность и иммунный гомеостаз менее точно определен. Foxp3 можно использовать в качестве хорошего маркера CD4 + CD25 + Т-клеток мыши, хотя недавние исследования также показали доказательства экспрессии Foxp3 в CD4 + CD25-Т-клетках. У людей Foxp3 также экспрессируется недавно активированными обычными Т-клетками и, таким образом, не определяет специфически человеческие Treg.[12]
Разработка
Все Т-клетки происходят из клеток-предшественников в Костный мозг, которые становятся приверженцами своего происхождения в вилочковая железа. Все Т-клетки начинаются как CD4 -CD8 -TCR - клетки на стадии DN (дважды отрицательной), когда отдельная клетка перестраивает свои гены рецепторов Т-клеток, чтобы сформировать уникальную функциональную молекулу, которую они, в свою очередь, тестируют против клеток коры тимуса на минимальный уровень взаимодействия с само-MHC. Если они получают эти сигналы, они пролиферируют и экспрессируют как CD4, так и CD8, становясь дважды положительными клетками. Отбор Treg происходит на радиоустойчивых гематопоэтических клетках, экспрессирующих MHC класса II в мозговом веществе или Тельца Хассала в тимусе. На стадии DP (двойной положительный результат) они выбираются путем взаимодействия с клетками в тимусе, начинают транскрипцию Foxp3 и становятся Treg-клетками, хотя они могут не начать экспрессировать Foxp3 до стадии одиночного положительного результата, т.е. в каком смысле они являются функциональными Tregs. У Tregs нет ограниченного TCR экспрессия NKT или γδ Т-клеток; Tregs имеют большее разнообразие TCR, чем эффекторные Т-клетки, смещенные в сторону собственных пептидов.
Процесс отбора Treg определяется аффинностью взаимодействия с самопептидным комплексом MHC. Выбор стать Treg - это «Златовласка Процесс - т.е. не слишком высокий, не слишком низкий, но в самый раз;[13] Т-клетка, которая получает очень сильные сигналы, подвергнется апоптотической смерти; клетка, которая получает слабый сигнал, выживет и станет эффекторной клеткой. Если Т-клетка получает промежуточный сигнал, она становится регуляторной клеткой. Из-за стохастический природа процесса активации Т-лимфоцитов, все популяции Т-лимфоцитов с данным TCR будут иметь смесь Т-лимфоцитов.эфф и Treg - относительные пропорции, определяемые сродством Т-клетки к собственному пептиду-MHC. Даже в моделях мышей с TCR-трансгенными клетками, отобранными на специфически-секретирующей антиген строме, делеция или преобразование не завершаются.
Foxp3+ Генерация Treg в тимусе задерживается на несколько дней по сравнению с Tэфф клеток и не достигает взрослых уровней ни в тимусе, ни на периферии примерно до трех недель после родов. Клеткам Treg требуется CD28 совместная стимуляция и B7.2 экспрессия в основном ограничена мозговым веществом, развитие которого, по-видимому, идет параллельно с развитием Foxp3+ клетки. Было высказано предположение, что эти два процесса связаны, но окончательной связи между процессами пока не было. TGF-β не требуется для функционирования Treg в тимусе, поскольку Treg тимуса из TGF-β нечувствительные к TGFβRII-DN мыши функциональны.
Тимическая рециркуляция
Было замечено, что некоторые Foxp3 + Treg-клетки рециркулируют обратно в тимус, где они и развиваются. Эти Treg в основном присутствовали в медуле тимуса, который является основным местом дифференцировки Treg-клеток.[14]Присутствие этих клеток в тимусе или добавление в культуру ткани тимуса плода подавляет развитие новых Treg-клеток на 34-60%,[15]но клетки Tconv не затронуты. Это означает, что рециркуляция Treg в тимус подавляет только de novo Развитие Treg-клеток. Молекулярный механизм этого процесса работает благодаря способности Treg адсорбировать ИЛ-2 из микроокружения, таким образом, будучи способным вызывать апоптоз других Т-клеток, которым ИЛ-2 является основным фактором роста.[16]рециркулирующие T reg клетки в тимусе экспрессируют высокое количество α-цепи высокоаффинного рецептора IL-2 (CD25 ) закодировано Il2ra ген, который собирает ИЛ-2 из мозгового вещества тимуса и снижает его концентрацию. Новые генерируемые Foxp3 + Treg-клетки в тимусе имеют не столь высокое количество Il2ra выражение.[17] IL-2 - это цитокин, необходимый для развития Treg-клеток в тимусе. Он важен для пролиферации и выживания Т-клеток, но в случае его дефицита может быть заменен ИЛ-15. Однако развитие Treg-клеток зависит от IL-2.[18] У человека обнаружена популяция CD31 отрицательные Treg-клетки в тимусе.[19] CD31 может использоваться в качестве маркера новых генерируемых Treg-клеток, как и другие Т-лимфоциты. Зрелые и периферические Treg-клетки снизили свою экспрессию.[20]Таким образом, возможно, что этот регуляторный механизм развития Treg-клеток тимуса также работает у человека.
Вероятно, существует также положительная регуляция развития Treg-клеток тимуса, вызванная рециркуляцией Treg-клеток в тимус. Обнаружена популяция CD24 низкий Foxp3 + в тимусе с повышенной экспрессией ИЛ-1Р2 (Il1r2) по сравнению с периферическими Treg-клетками.[21][22] Высокая концентрация ИЛ-1β вызванное воспалением уменьшение de novo развитие Treg-клеток в тимусе.[22] Присутствие рециркулирующих клеток Treg в тимусе с высокой экспрессией IL1R2 во время воспалительных состояний помогает поглощать IL1β и снижать его концентрацию в микросреде мозгового вещества, таким образом, они способствуют развитию de novo Treg-клетки.[22] Высокая концентрация ИЛ-1β вызванное воспалением уменьшение de novo развитие Treg-клеток в тимусе.[22] Связывание IL-1β с IL1R2 на поверхности Treg-клеток не вызывает какой-либо передачи сигнала, поскольку отсутствует внутрицеллюлозный (МДП ) Toll-рецепторный домен интерлейкина-1, который обычно присутствует в клетках врожденного иммунитета.[23]
Функция
В иммунная система должен уметь различать «я» и «не-я». Когда само / несамо-различение не удается, иммунная система разрушает клетки и ткани тела и, как следствие, вызывает аутоиммунные заболевания. Регуляторные Т-клетки активно подавляют активацию иммунной системы и предотвращают патологическую самореактивность, то есть аутоиммунное заболевание. Критическая роль регуляторных Т-клеток в иммунной системе подтверждается тяжелым аутоиммунным синдромом, который возникает в результате генетической недостаточности регуляторных Т-клеток (Синдром IPEX - см. Также ниже).
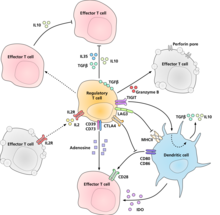
Молекулярный механизм, с помощью которого регуляторные Т-клетки проявляют свою супрессорную / регуляторную активность, окончательно не охарактеризован и является предметом интенсивных исследований. В пробирке эксперименты дали смешанные результаты относительно необходимости контакта клетки с клеткой, которая подавляется. Ниже представлены некоторые из предложенных механизмов подавления иммунитета:
- Регуляторные Т-клетки продуцируют ряд ингибирующих цитокинов. К ним относятся Преобразование фактора роста в бета,[24] Интерлейкин 35,[25] и Интерлейкин 10.[26] Также оказывается, что регуляторные Т-клетки могут побуждать другие типы клеток к экспрессии интерлейкина-10.[27]
- Регуляторные Т-клетки могут производить Гранзим B, что, в свою очередь, может вызвать апоптоз эффекторных клеток. Сообщается, что регуляторные Т-клетки от мышей с дефицитом гранзима В являются менее эффективными супрессорами активации эффекторных Т-клеток.[28]
- Обратная передача сигналов посредством прямого взаимодействия с дендритные клетки и индукция иммуносупрессивного индоламин-2,3-диоксигеназа.[29]
- Передача сигналов через эктоферменты CD39 и CD73 с выработкой иммуносупрессивных аденозин.[30][31]
- Через прямое взаимодействие с дендритными клетками LAG3 и по TIGIT.[32][33]
- Другой механизм управления - через контур обратной связи ИЛ-2. Активированные антигеном Т-клетки продуцируют ИЛ-2, который затем воздействует на рецепторы ИЛ-2 на регуляторные Т-клетки, предупреждая их о том, что в этом регионе наблюдается высокая активность Т-клеток, и они вызывают подавляющий ответ против них. Это цикл отрицательной обратной связи, позволяющий избежать чрезмерной реакции. Если присутствует реальная инфекция, другие воспалительные факторы снижают подавление. Нарушение петли приводит к гиперреактивности, регуляция может изменять силу иммунного ответа.[34] Связанное предложение относительно интерлейкин 2 заключается в том, что активированные регуляторные Т-клетки поглощают интерлейкин 2 настолько сильно, что лишают эффекторные Т-клетки достаточного количества, чтобы избежать апоптоза.[16]
- Основным механизмом подавления регуляторными Т-клетками является предотвращение совместная стимуляция через CD28 на эффекторные Т-клетки под действием молекулы CTLA-4.[35]
Индуцированные регуляторные Т-клетки
Индуцированные регуляторные T (iTreg) клетки (CD4+ CD25+ Foxp3+) являются супрессивными клетками, участвующими в толерантности. Было показано, что клетки iTreg подавляют пролиферацию Т-клеток и экспериментальные аутоиммунные заболевания. Эти ячейки включают Клетки Treg17. Клетки iTreg развиваются из зрелых CD4+ обычные Т-клетки вне тимуса: определяющее различие между естественными регуляторными Т-клетками (nTreg) и клетками iTreg. Хотя клетки iTreg и nTreg имеют схожую функцию, недавно было показано, что клетки iTreg являются «существенным неизбыточным регуляторным подмножеством, которое дополняет клетки nTreg, частично за счет увеличения разнообразия TCR в рамках регуляторных ответов».[36] Острое истощение пула клеток iTreg на моделях мышей привело к воспалению и потере веса. Вклад клеток nTreg по сравнению с клетками iTreg в поддержании толерантности неизвестен, но оба важны. Эпигенетический наблюдались различия между клетками nTreg и iTreg, причем первые имели более стабильную экспрессию Foxp3 и более широкую деметилирование.
Окружающая среда тонкого кишечника богата витамином А и является местом производства ретиноевой кислоты.[37] Ретиноевая кислота и TGF-бета, продуцируемые дендритными клетками в этой области, сигнализируют о продукции регуляторных Т-клеток.[37] Витамин А и TGF-бета способствуют дифференцировке Т-клеток в регуляторные Т-клетки, в отличие от Клетки Th17, даже при наличии Ил-6.[38][39] Окружающая среда кишечника может привести к индуцированию регуляторных Т-клеток (iTregs) с TGF-бета и ретиноевой кислотой,[40] некоторые из них экспрессируют лектин-подобный рецептор CD161 и специализируются на поддержании целостности барьера за счет ускорения заживления ран.[41] Treg в кишечнике дифференцируются от наивных Т-клеток после введения антигена.[42]
Болезнь
Важным вопросом в области иммунологии является то, как модулируется иммуносупрессивная активность регуляторных Т-клеток в ходе продолжающегося иммунного ответа. Хотя иммуносупрессивная функция регуляторных Т-клеток предотвращает развитие аутоиммунного заболевания, это нежелательно во время иммунных ответов на инфекционные микроорганизмы. Современные гипотезы предполагают, что при столкновении с инфекционными микроорганизмами активность регуляторных Т-клеток может прямо или косвенно подавляться другими клетками, чтобы облегчить устранение инфекции. Экспериментальные данные, полученные на мышах, предполагают, что некоторые патогены, возможно, эволюционировали, чтобы манипулировать регуляторными Т-клетками, подавляя иммунитет хозяина и тем самым усиливая свое собственное выживание. Например, сообщалось о повышении активности регуляторных Т-клеток в некоторых инфекционных контекстах, таких как ретровирусный инфекции (наиболее известными из которых являются ВИЧ ), микобактериальные инфекции (например, туберкулез ), и различные паразитарные инфекции, включая Лейшмания и малярия.
Треги играют главные роли во время ВИЧ инфекционное заболевание. Они подавляют иммунную систему, таким образом ограничивая клетки-мишени и уменьшая воспаление, но это одновременно нарушает клиренс вируса клеточно-опосредованным иммунным ответом и увеличивает резервуар, выталкивая CD4.+ Т-клетки в состоянии покоя, включая инфицированные клетки. Кроме того, Treg могут быть инфицированы ВИЧ, что напрямую увеличивает размер резервуара ВИЧ. Таким образом, Treg исследуются как цели для исследований по лечению ВИЧ.[43] Некоторые стратегии истощения Treg были протестированы в SIV зараженный нечеловеческие приматы, и показано, что он вызывает реактивацию вируса и увеличивает специфический для SIV CD8+ Ответы Т-клеток.[44]
Регуляторные Т-клетки играют большую роль в патологии висцеральный лейшманиоз и в предотвращении чрезмерного воспаления у пациентов, излеченных от висцерального лейшманиоза.
CD4+ регуляторные Т-клетки часто связаны с солидными опухолями как у людей, так и у мышей. Повышенное количество регуляторных Т-клеток при раке молочной железы, колоректального рака и яичников связано с более плохим прогнозом.[45]
CD70 + В-клетки неходжкинской лимфомы индуцируют экспрессию Foxp3 и регулирующую функцию внутриопухолевого CD4+CD25- Т-клетки.[46]
Недавнее исследование показывает, что церебральная ишемия может увеличивать CD4 (+) CD25 (+) Foxp3 (+) регуляторные Т-клетки костного мозга через сигналы симпатической нервной системы.[47]
Есть некоторые свидетельства того, что Tregs могут быть дисфункциональными и вызывать нейровоспаление в боковой амиотрофический склероз из-за более низкой экспрессии Foxp3.[48] Ex vivo Расширение Treg для последующей аутотрансплантации в настоящее время изучается после получения многообещающих результатов в ходе фазы I клинических испытаний.[49]
Кроме того, хотя было показано, что регуляторные Т-клетки увеличиваются посредством поликлональной экспансии как системно, так и локально во время здоровой беременности, чтобы защитить плод от материнского иммунного ответа (процесс, называемый материнской иммунной толерантностью), есть доказательства того, что это поликлональное расширение нарушается при преэклампсии матери и их потомство.[50] Исследования показывают, что снижение выработки и развития регуляторных Т-клеток во время преэклампсии может ухудшить материнскую иммунную толерантность, что приведет к гиперактивному иммунному ответу, характерному для преэклампсии.[51]
Рак

Большинство опухолей вызывают у хозяина иммунный ответ, опосредованный опухолевыми антигенами, что позволяет отличить опухоль от других незлокачественных клеток. Это вызывает большое количество лимфоциты, инфильтрирующие опухоль (TIL), которые можно найти в микросреде опухоли.[52] Хотя это не совсем понятно, считается, что эти лимфоциты нацелены на раковые клетки и, следовательно, замедляют или прекращают развитие опухоли. Однако этот процесс сложен, потому что кажется, что Treg предпочтительно доставляются в микросреду опухоли. Хотя Treg обычно составляют только около 4% CD4.+ Т-клетки, они могут составлять до 20–30% от общего количества CD4.+ популяция вокруг микросреды опухоли.[53]
Хотя изначально считалось, что высокие уровни TIL важны для определения иммунного ответа против рака, в настоящее время широко признано, что соотношение Treg к Teffectors в микроокружении опухоли является определяющим фактором успеха иммунного ответа против рака. Высокие уровни Treg в микросреде опухоли связаны с плохим прогнозом многих видов рака,[54] такие как рак яичников, груди, почек и поджелудочной железы.[53] Это указывает на то, что Treg подавляют клетки Teffector и препятствуют иммунному ответу организма против рака. Однако при некоторых типах рака верно обратное, и высокие уровни Treg связаны с положительным прогнозом. Эта тенденция наблюдается при раке, таком как колоректальная карцинома и фолликулярная лимфома. Это может быть связано со способностью Treg подавлять общее воспаление, которое, как известно, вызывает пролиферацию клеток и метастазирование.[53] Эти противоположные эффекты указывают на то, что роль Treg в развитии рака сильно зависит как от типа, так и от местоположения опухоли.
Хотя до сих пор не совсем понятно, каким образом Treg преимущественно доставляются в микросреду опухоли, хемотаксис, вероятно, обусловлен производством хемокинов опухолью. Инфильтрация Treg в микроокружение опухоли облегчается связыванием хемокинового рецептора CCR4, который экспрессируется на Treg, с его лигандом CCL22, который секретируется многими типами опухолевых клеток.[55] Экспансия Treg в месте опухоли также может объяснить повышенные уровни Treg. Известно, что цитокин TGF-β, который обычно продуцируется опухолевыми клетками, вызывает дифференцировку и экспансию Treg.[55]
В общем, иммуносупрессия микросреды опухоли в значительной степени способствовала неудачным результатам многих видов лечения иммунотерапией рака. Истощение Treg в моделях на животных показало повышенную эффективность иммунотерапевтического лечения, и поэтому многие виды иммунотерапевтического лечения теперь включают истощение Treg.[2]
Молекулярная характеристика
Подобно другим Т-клеткам, регуляторные Т-клетки развиваются в вилочковая железа. Последние исследования показывают, что регуляторные Т-клетки определяются экспрессией семейства вилкоедов. фактор транскрипции Foxp3 (коробка с вилкой p3). Выражение Foxp3 необходим для развития регуляторных Т-клеток и, по-видимому, контролирует генетическую программу, определяющую судьбу этой клетки.[56] Подавляющее большинство регуляторных Т-клеток, экспрессирующих Foxp3, находится в главный комплекс гистосовместимости (MHC) класс II ограничен CD 4-экспрессирующий (CD4+) населения и выражают высокие уровни интерлейкин-2 рецепторная альфа-цепь (CD25). Помимо Foxp3-экспрессирующего CD4+ CD25+, также, по-видимому, существует небольшая популяция CD8, ограниченная MHC класса I+ Foxp3-экспрессирующие регуляторные Т-клетки. Эти Foxp3-экспрессирующие CD8+ Т-клетки, по-видимому, не функционируют у здоровых людей, но индуцируются при аутоиммунных болезненных состояниях за счет стимуляции рецептора Т-клеток для подавления опосредованных IL-17 иммунных ответов.[57] В отличие от обычных Т-клеток, регуляторные Т-клетки не продуцируют ИЛ-2 и поэтому изначально анергичны.
В исследованиях используется ряд различных методов для идентификации и мониторинга Treg-клеток. Первоначально использовалась высокая экспрессия поверхностных маркеров CD25 и CD4 (CD4+CD25+ ячеек). Это проблематично, поскольку CD25 также экспрессируется на нерегулирующих Т-клетках в условиях активации иммунной системы, например, во время иммунного ответа на патоген. Согласно определению экспрессии CD4 и CD25, регуляторные Т-клетки составляют около 5-10% зрелых CD4.+ Субпопуляция Т-клеток у мышей и людей, в то время как в цельной крови можно измерить около 1-2% Treg. Дополнительное измерение клеточной экспрессии белка Foxp3 позволило провести более специфический анализ клеток Treg (CD4+CD25+Foxp3+ ячеек). Однако Foxp3 также временно экспрессируется в активированных эффекторных Т-клетках человека, что усложняет правильный анализ Treg с использованием CD4, CD25 и Foxp3 в качестве маркеров у людей. Поэтому некоторые исследовательские группы используют другой маркер отсутствия или низкого уровня экспрессии поверхностного белка CD127 в сочетании с наличием CD4 и CD25. Было описано несколько дополнительных маркеров, например, высокие уровни CTLA-4 (молекулы-4, ассоциированные с цитотоксическими Т-лимфоцитами) и GITR (глюкокортикоид-индуцированный рецептор TNF) также экспрессируются на регуляторных Т-клетках, однако функциональное значение этой экспрессии еще предстоит определить. Существует большой интерес к идентификации маркеров клеточной поверхности, которые уникально и специфически экспрессируются на всех экспрессирующих Foxp3 регуляторных Т-клетках. Однако на сегодняшний день такая молекула не идентифицирована.
Помимо поиска новых белковых маркеров, в литературе был описан другой метод более точного анализа и мониторинга Treg-клеток. Этот метод основан на Метилирование ДНК анализ. Только в Treg-клетках, но не в любом другом типе клеток, включая активированные эффекторные Т-клетки, определенная область в пределах гена Foxp3 (TSDR, Treg-специфически-деметилированная область) обнаруживается деметилированной, что позволяет контролировать Treg-клетки с помощью реакции ПЦР. или другие методы анализа на основе ДНК.[58]Взаимодействие между клетками Th17 и регуляторными Т-клетками важно при многих заболеваниях, таких как респираторные заболевания.[59]
Последние данные свидетельствуют о том, что тучные клетки могут быть важными медиаторами Treg-зависимой периферической толерантности.[60]
Эпитопы
Регуляторные Т-клеточные эпитопы («Трегитопы») были открыты в 2008 году и состоят из линейных последовательностей аминокислот, содержащихся в моноклональных антителах и иммуноглобулинах G (IgG). С момента их открытия данные показали, что трегитопы могут иметь решающее значение для активации естественных регуляторных Т-клеток.[61][62][63]
Были выдвинуты гипотезы о возможных применениях регуляторных Т-клеточных эпитопов: толеризация трансплантатов, белковые препараты, методы лечения трансплантации крови и диабет I типа а также сокращение иммунная реакция для лечения аллергия.[64][65][66][67][68][69][63]
Генетическая недостаточность
Генетические мутации в гене, кодирующем Foxp3, были идентифицированы как у людей, так и у мышей на основании наследственного заболевания, вызванного этими мутациями. Это заболевание является наиболее ярким свидетельством того, что регуляторные Т-клетки играют решающую роль в поддержании нормальной функции иммунной системы. Люди с мутациями в Foxp3 страдают тяжелым и быстро фатальным аутоиммунным заболеванием, известным как янарушение регуляции mmune, полендокринопатия, Eэнтеропатия Икс-связанный (IPEX) синдром.[70][71]
В IPEX Синдром характеризуется развитием подавляющего системного аутоиммунитета в течение первого года жизни, что приводит к обычно наблюдаемой триаде водянистой диареи, экзематозного дерматита и эндокринопатии, которая чаще всего рассматривается как инсулинозависимая сахарный диабет. У большинства людей есть другие аутоиммунные явления, включая гемолитическую анемию, положительную по Кумбсу, аутоиммунную тромбоцитопению, аутоиммунную нейтропению и тубулярную нефропатию. Большинство заболевших мужчин умирают в течение первого года жизни либо от метаболических нарушений, либо от сепсиса. Аналогичное заболевание также наблюдается у спонтанных мутантных по Foxp3 мышей, известных как «scurfy».
Рекомендации
- ^ Bettelli E, Carrier Y, Gao W., Korn T., Strom TB, Oukka M, Weiner HL, Kuchroo VK (май 2006 г.). «Взаимные пути развития для генерации патогенных эффекторных TH17 и регуляторных Т-клеток». Природа. 441 (7090): 235–8. Bibcode:2006Натура.441..235Б. Дои:10.1038 / природа04753. PMID 16648838. S2CID 4391497.
- ^ а б Куриэль Т.Дж. (май 2007 г.). «Tregs и переосмысление иммунотерапии рака». Журнал клинических исследований. 117 (5): 1167–74. Дои:10.1172 / JCI31202. ЧВК 1857250. PMID 17476346.
- ^ Чен В. (август 2011 г.). «Tregs в иммунотерапии: возможности и проблемы». Иммунотерапия. 3 (8): 911–4. Дои:10.2217 / imt.11.79. PMID 21843075.
- ^ Мияра М., Горохов Г., Эренштейн М., Мюссет Л., Сакагучи С., Amoura Z (октябрь 2011 г.). «Человеческие FoxP3 + регуляторные Т-клетки при системных аутоиммунных заболеваниях». Отзывы об аутоиммунности. 10 (12): 744–55. Дои:10.1016 / j.autrev.2011.05.004. PMID 21621000.
- ^ Носбаум А., Превел Н., Чыонг Х.А., Мехта П., Эттингер М., Шаршмидт Т.С., Али Н.Х., Паули М.Л., Аббас А.К., Розенблюм М.Д. (март 2016 г.). «Передний край: регуляторные Т-клетки способствуют заживлению кожных ран». J Immunol. 196 (5): 2010–4. Дои:10.4049 / jimmunol.1502139. ЧВК 4761457. PMID 26826250.
- ^ Адигбе Д.О., Нисикава Х. (2013). «Природные и индуцированные Т-регуляторные клетки при раке». Границы иммунологии. 4: 190. Дои:10.3389 / fimmu.2013.00190. ЧВК 3708155. PMID 23874336.
- ^ Куриэль Т.Дж. (апрель 2008 г.). «Регуляторные Т-клетки и лечение рака». Текущее мнение в иммунологии. 20 (2): 241–6. Дои:10.1016 / j.coi.2008.04.008. ЧВК 3319305. PMID 18508251.
- ^ Хори С., Номура Т., Сакагути С. (февраль 2003 г.). «Контроль развития регуляторных Т-клеток с помощью фактора транскрипции Foxp3». Наука. 299 (5609): 1057–61. Bibcode:2003Наука ... 299.1057H. Дои:10.1126 / science.1079490. PMID 12522256. S2CID 9697928.
- ^ Сингх Б., Шварц Дж. А., Сандрок С., Беллемор С. М., Никоопур Е. (ноябрь 2013 г.). «Модуляция аутоиммунных заболеваний с помощью продуцирующих интерлейкин (IL) -17 регуляторных Т-хелперных (Th17) клеток». Индийский журнал медицинских исследований. 138 (5): 591–4. ЧВК 3928692. PMID 24434314.
- ^ Шевач Э.М. (2000). «Регуляторные Т-клетки при аутоиммунитете *». Ежегодный обзор иммунологии. 18: 423–49. Дои:10.1146 / annurev.immunol.18.1.423. PMID 10837065. S2CID 15160752.
- ^ Schmetterer, Klaus G .; Нойнкирхнер, Алина; Пикл, Винфрид Ф. (июнь 2012 г.). «Природные регуляторные Т-клетки: маркеры, механизмы и манипуляции». Журнал FASEB. 26 (6): 2253–2276. Дои:10.1096 / fj.11-193672. PMID 22362896. S2CID 36277557.
- ^ Сакагути С. (2004). «Естественно возникающие регуляторные Т-клетки CD4 + для иммунологической толерантности и отрицательного контроля иммунных ответов». Ежегодный обзор иммунологии. 22: 531–62. Дои:10.1146 / annurev.immunol.21.120601.141122. PMID 15032588.
- ^ Ли М. (август 2016 г.). «Передача сигналов рецептора Т-клеток в контроле дифференцировки и функции регуляторных Т-клеток». Nature Reviews Иммунология. 16 (4): 220–233. Дои:10.1038 / нет.2016.26. ЧВК 4968889. PMID 27026074.
- ^ Тио, Николя; Дарригес, Джули; Адуэ, Вероник; Gros, Marine; Бине, Бенедикт; Пералс, Корин; Леобон, Бертран; Фазилло, Николя; Жоффр, Оливье П.; Роби, Эллен А.; van Meerwijk, Joost P.M; Романьоли, Паола (4 мая 2015 г.).«Периферические регуляторные Т-лимфоциты, рециркулирующие в тимус, подавляют развитие своих предшественников». Иммунология природы. 16 (6): 628–634. Дои:10.1038 / ni.3150. PMID 25939024. S2CID 7670443.
- ^ Тио, Николя; Дарригес, Джули; Адуэ, Вероник; Gros, Marine; Бине, Бенедикт; Пералс, Корин; Леобон, Бертран; Фазилло, Николя; Жоффр, Оливье П.; Роби, Эллен А.; van Meerwijk, Joost P.M; Романьоли, Паола (4 мая 2015 г.). «Периферические регуляторные Т-лимфоциты, рециркулирующие в тимус, подавляют развитие своих предшественников». Иммунология природы. 16 (6): 628–634. Дои:10.1038 / ni.3150. PMID 25939024. S2CID 7670443.
- ^ а б Пандиян, Пушпа; Чжэн, Лисинь; Исихара, Сатору; Рид, Дженнифер; Ленардо, Майкл Дж (4 ноября 2007 г.). «CD4 + CD25 + Foxp3 + регуляторные Т-клетки вызывают апоптоз эффекторных CD4 + Т-клеток, опосредованный депривацией цитокинов». Иммунология природы. 8 (12): 1353–1362. Дои:10.1038 / ni1536. PMID 17982458. S2CID 8925488.
- ^ Тио, Николя; Дарригес, Джули; Адуэ, Вероник; Gros, Marine; Бине, Бенедикт; Пералс, Корин; Леобон, Бертран; Фазилло, Николя; Жоффр, Оливье П.; Роби, Эллен А.; van Meerwijk, Joost P.M; Романьоли, Паола (4 мая 2015 г.). «Периферические регуляторные Т-лимфоциты, рециркулирующие в тимус, подавляют развитие своих предшественников». Иммунология природы. 16 (6): 628–634. Дои:10.1038 / ni.3150. PMID 25939024. S2CID 7670443.
- ^ Чэн, Гоянь; Ю, Айсинь; Малек, Томас Р. (май 2011 г.). «Т-клеточная толерантность и многофункциональная роль передачи сигналов IL-2R в Т-регуляторных клетках». Иммунологические обзоры. 241 (1): 63–76. Дои:10.1111 / j.1600-065X.2011.01004.x. ЧВК 3101713. PMID 21488890.
- ^ Тио, Николя; Дарригес, Джули; Адуэ, Вероник; Gros, Marine; Бине, Бенедикт; Пералс, Корин; Леобон, Бертран; Фазилло, Николя; Жоффр, Оливье П.; Роби, Эллен А.; van Meerwijk, Joost P.M; Романьоли, Паола (4 мая 2015 г.). «Периферические регуляторные Т-лимфоциты, рециркулирующие в тимус, подавляют развитие своих предшественников». Иммунология природы. 16 (6): 628–634. Дои:10.1038 / ni.3150. PMID 25939024. S2CID 7670443.
- ^ Киммиг, Соня; Przybylski, Grzegorz K .; Schmidt, Christian A .; Лауриш, Катя; Möwes, Беате; Радбрух, Андреас; Тиль, Андреас (18 марта 2002 г.). «Два подмножества наивных Т-хелперов с различным содержанием круга вырезания Т-клеточного рецептора в периферической крови взрослого человека». Журнал экспериментальной медицины. 195 (6): 789–794. Дои:10.1084 / jem.20011756. ЧВК 2193736. PMID 11901204.
- ^ Токер, Арас; Энгельберт, Дирк; Гарг, Гарима; Поланский, Юлия К .; Флоесс, Стефан; Мияо, Такахиса; Барон, Удо; Дюбер, Сандра; Гефферс, Роберт; Гир, Паскаль; Шалленберг, Соня; Кречмер, Карстен; Олек, Свен; Вальтер, Йорн; Вайс, Зигфрид; Хори, Шохей; Хаманн, Альф; Хуэн, Йохен (1 апреля 2013 г.). «Активное деметилирование Foxp3 Локус ведет к образованию стабильных регуляторных Т-клеток в тимусе ». Журнал иммунологии. 190 (7): 3180–3188. Дои:10.4049 / jimmunol.1203473. PMID 23420886.
- ^ а б c d Николули, Эйрини; Эльфаки, Ясин; Херппич, Сюзанна; Шельбауэр, Карстен; Делакер, Майкл; Фальк, Кристина; Муфазалов, Ильгиз А .; Вайсман, Ари; Feuerer, Маркус; Хуэн, Йохен (27 января 2020 г.). «Рециркулирующие IL-1R2 + Treg точно регулируют развитие интратимических Treg при воспалительных условиях». Клеточная и молекулярная иммунология. Дои:10.1038 / s41423-019-0352-8. HDL:10033/622148. PMID 31988493. S2CID 210913733.
- ^ Питерс, Ванесса А .; Джостинг, Дженнифер Дж .; Фройнд, Грегори Г. (2013). «Рецептор 2 IL-1 (IL-1R2) и его роль в иммунной регуляции». Мозг, поведение и иммунитет. 32: 1–8. Дои:10.1016 / j.bbi.2012.11.006. ЧВК 3610842. PMID 23195532.
- ^ Прочтите S, Malmström V, Powrie F (июль 2000 г.). «Цитотоксический Т-лимфоцит-ассоциированный антиген 4 играет важную роль в функции CD25 (+) CD4 (+) регуляторных клеток, которые контролируют воспаление кишечника». Журнал экспериментальной медицины. 192 (2): 295–302. Дои:10.1084 / jem.192.2.295. ЧВК 2193261. PMID 10899916.
- ^ Коллисон Л.В., Уоркман С.Дж., Куо Т.Т., Бойд К., Ван Й., Виньяли К.М., Кросс Р., Сехи Д., Блумберг Р.С., Виньяли Д.А. (ноябрь 2007 г.). «Ингибирующий цитокин IL-35 способствует регуляторной функции Т-клеток». Природа. 450 (7169): 566–9. Bibcode:2007Натура.450..566С. Дои:10.1038 / природа06306. PMID 18033300. S2CID 4425281.
- ^ Аннакер О., Ассеман С., Рид С., Паури Ф. (июнь 2003 г.). «Интерлейкин-10 в регуляции Т-клеточного колита». Журнал аутоиммунитета. 20 (4): 277–9. Дои:10.1016 / s0896-8411 (03) 00045-3. PMID 12791312.
- ^ Кирли Дж., Баркер Дж. Э., Робинсон Д.С., Ллойд К.М. (декабрь 2005 г.). «Разрешение воспаления дыхательных путей и гиперреактивности после переноса in vivo CD4 + CD25 + регуляторных Т-клеток зависит от интерлейкина 10». Журнал экспериментальной медицины. 202 (11): 1539–47. Дои:10.1084 / jem.20051166. ЧВК 1350743. PMID 16314435.
- ^ Гондек, округ Колумбия, Лу Л.Ф., Кесада С.А., Сакагути С., Ноэль Р.Дж. (февраль 2005 г.). «Передний край: контактно-опосредованное подавление регуляторными клетками CD4 + CD25 + включает гранзим B-зависимый, перфорин-независимый механизм». Журнал иммунологии. 174 (4): 1783–6. Дои:10.4049 / jimmunol.174.4.1783. PMID 15699103.
- ^ Пуччетти П., Громанн Ю. (октябрь 2007 г.). «IDO и регуляторные Т-клетки: роль в обратной передаче сигналов и неканонической активации NF-kappaB». Обзоры природы. Иммунология. 7 (10): 817–23. Дои:10.1038 / nri2163. PMID 17767193. S2CID 5544429.
- ^ Borsellino G, Kleinewietfeld M, Di Mitri D, Sternjak A, Diamantini A, Giometto R, Höpner S, Centonze D, Bernardi G, Dell'Acqua ML, Rossini PM, Battistini L, Rötzschke O, Falk K (август 2007). «Экспрессия эктонуклеотидазы CD39 клетками Foxp3 + Treg: гидролиз внеклеточного АТФ и подавление иммунитета». Кровь. 110 (4): 1225–32. Дои:10.1182 / кровь-2006-12-064527. PMID 17449799.
- ^ Коби Дж. Дж., Шах PR, Ян Л., Ребхан Дж. А., Фауэлл Д. Д., Мосманн Т. Р. (ноябрь 2006 г.). «Т-регуляторные и примированные незафиксированные CD4-Т-клетки экспрессируют CD73, который подавляет эффекторные Т-клетки CD4 путем преобразования 5'-аденозинмонофосфата в аденозин». Журнал иммунологии. 177 (10): 6780–6. Дои:10.4049 / jimmunol.177.10.6780. PMID 17082591.
- ^ Хуанг CT, Уоркман CJ, Flies D, Pan X, Marson AL, Zhou G, Hipkiss EL, Ravi S, Kowalski J, Levitsky HI, Powell JD, Pardoll DM, Drake CG, Vignali DA (октябрь 2004 г.). «Роль LAG-3 в регуляторных Т-клетках». Иммунитет. 21 (4): 503–13. Дои:10.1016 / j.immuni.2004.08.010. PMID 15485628.
- ^ Ю Икс, Харден К., Гонсалес Л.С., Франческо М., Чанг Э., Ирвинг Б., Том I, Ивелья С., Рефино С.Дж., Кларк Х., Итон Д., Гроган Д.Л. (январь 2009 г.). «Поверхностный белок TIGIT подавляет активацию Т-клеток, способствуя образованию зрелых иммунорегуляторных дендритных клеток». Иммунология природы. 10 (1): 48–57. Дои:10.1038 / ni.1674. PMID 19011627. S2CID 205361984.
- ^ Сакагути С., Ямагути Т., Номура Т., Оно М. (май 2008 г.). «Регуляторные Т-клетки и иммунная толерантность». Клетка. 133 (5): 775–87. Дои:10.1016 / j.cell.2008.05.009. PMID 18510923.
- ^ Уокер Л.С., Сансом Д.М. (ноябрь 2011 г.). «Возникающая роль CTLA4 как внешнего регулятора Т-клеточных ответов». Обзоры природы. Иммунология. 11 (12): 852–63. Дои:10.1038 / nri3108. PMID 22116087. S2CID 9617595.
- ^ Харибхай Д., Уильямс Дж. Б., Цзя С., Никерсон Д., Шмитт Э. Г., Эдвардс Б., Зигельбауэр Дж., Яссай М., Ли Ш., Релланд Л. М., Мудрый П. М., Чен А., Чжэн И. К., Симпсон П. М., Горски Дж., Зальцман Н.Х., Хесснер М.Дж. , Chatila TA, Williams CB (июль 2011 г.). «Необходимая роль индуцированных регуляторных Т-клеток в толерантности, основанная на расширении разнообразия рецепторов антигена». Иммунитет. 35 (1): 109–22. Дои:10.1016 / j.immuni.2011.03.029. ЧВК 3295638. PMID 21723159.
- ^ а б Sun CM, Hall JA, Blank RB, Bouladoux N, Oukka M, Mora JR, Belkaid Y (август 2007 г.). «Дендритные клетки собственной пластинки тонкой кишки способствуют de novo генерации клеток Foxp3 T reg с помощью ретиноевой кислоты». Журнал экспериментальной медицины. 204 (8): 1775–85. Дои:10.1084 / jem.20070602. ЧВК 2118682. PMID 17620362.
- ^ Муцида Д., Пак Й., Ким Дж., Туровская О., Скотт И., Кроненберг М., Черутре Х (июль 2007 г.). «Реципрокная дифференцировка TH17 и регуляторных Т-клеток, опосредованная ретиноевой кислотой». Наука. 317 (5835): 256–60. Bibcode:2007Наука ... 317..256М. Дои:10.1126 / science.1145697. PMID 17569825. S2CID 24736012.
- ^ Эркеленс М.Н., Мебиус Р.Э. (март 2017 г.). «Ретиноевая кислота и иммунный гомеостаз: баланс». Тенденции в иммунологии. 38 (3): 168–180. Дои:10.1016 / j.it.2016.12.006. PMID 28094101.
- ^ Циглер С.Ф., Бакнер Дж. Х. (апрель 2009 г.). «FOXP3 и регуляция дифференцировки Treg / Th17». Микробы и инфекции. 11 (5): 594–8. Дои:10.1016 / j.micinf.2009.04.002. ЧВК 2728495. PMID 19371792.
- ^ Поволери, Джованни А. М .; Нова-Ламперти, Эстефания; Скотта, Криштиану; Fanelli, Giorgia; Чен, Юнь-Цзин; Беккер, Пабло Д.; Бордман, Доминик; Костантини, Бенедетта; Романо, Марко; Павлидис, Полихронис; МакГрегор, Рубен; Пантази, Эйрини; Чаусс, Даниэль; Сунь, Хун-Вэй; Ши, Хан-Ю; Казинс, Дэвид Дж .; Купер, Николай; Пауэлл, Ник; Кемпер, Клаудиа; Пирожня, Мехди; Лоуренс, Ариан; Кордасти, Шахрам; Каземян, Маджид; Ломбарди, Джованна; Афзали, Бехдад (5 ноября 2018 г.). «Человеческие регулируемые ретиноевой кислотой CD161 + регуляторные Т-клетки поддерживают заживление ран в слизистой оболочке кишечника». Иммунология природы. 19 (12): 1403–1414. Дои:10.1038 / с41590-018-0230-з. ЧВК 6474659. PMID 30397350.
- ^ Coombes JL, Siddiqui KR, Arancibia-Cárcamo CV, Hall J, Sun CM, Belkaid Y, Powrie F (август 2007 г.). «Функционально специализированная популяция CD103 + DC слизистой оболочки индуцирует Foxp3 + регуляторные Т-клетки через TGF-бета и механизм, зависимый от ретиноевой кислоты». Журнал экспериментальной медицины. 204 (8): 1757–64. Дои:10.1084 / jem.20070590. ЧВК 2118683. PMID 17620361.
- ^ Клейнман А.Дж., Шиванандхам Р., Пандреа I, Шугнет, Калифорния, Апетрей С. (2018). «Регуляторные Т-клетки как потенциальные мишени для исследований по лечению ВИЧ». Границы иммунологии. 9: 734. Дои:10.3389 / fimmu.2018.00734. ЧВК 5908895. PMID 29706961.
- ^ Шиванандхам, Ранджит; Клейнман, Адам Дж .; Сетте, Паола; Брокка-Кофано, Эджидио; Килапандал Венкатраман, Синдхуджа Мурали; Policicchio, Benjamin B .; Он, Тяньюй; Сюй, Куйлинг; Swarthout, Джулия; Ван, Чжируй; Пандреа, Ивона (15.07.2020). «Тестирование нечеловеческих приматов воздействия различных стратегий истощения Treg на реактивацию и устранение скрытого вируса иммунодефицита обезьян». Журнал вирусологии. 94 (19): JVI.00533–20, jvi, JVI.00533–20v1. Дои:10.1128 / JVI.00533-20. ISSN 0022-538X. PMID 32669326. S2CID 220579402.
- ^ Дранофф G (декабрь 2005 г.). «Терапевтическое значение внутриопухолевых регуляторных Т-клеток». Клинические исследования рака. 11 (23): 8226–9. Дои:10.1158 / 1078-0432.CCR-05-2035. PMID 16322278.
- ^ Ян З.З., Новак А.Дж., Цисмер С.К., Витциг Т.Э., Анселл С.М. (октябрь 2007 г.). «CD70 + В-клетки неходжкинской лимфомы индуцируют экспрессию Foxp3 и регулирующую функцию во внутриопухолевых CD4 + CD25 Т-клетках». Кровь. 110 (7): 2537–44. Дои:10.1182 / кровь-2007-03-082578. ЧВК 1988926. PMID 17615291.
- ^ Ван Дж, Ю Л, Цзян Ц., Фу Х, Лю Х, Ван М., Оу Ц, Цуй Х, Чжоу Ц., Ван Дж. (Январь 2015 г.). «Церебральная ишемия увеличивает CD4 + CD25 + FoxP3 + регуляторные Т-клетки костного мозга у мышей посредством сигналов симпатической нервной системы». Мозг, поведение и иммунитет. 43: 172–83. Дои:10.1016 / j.bbi.2014.07.022. ЧВК 4258426. PMID 25110149.
- ^ Beers, David R .; Чжао, Вэйхуа; Ван, Цзинхун; Чжан, Сюцзюнь; Вэнь, Шисян; Нил, Дэн; Тонхофф, Джейсон Р .; Alsuliman, Abdullah S .; Шпалл, Элизабет Дж .; Резвани, Кэти; Аппель, Стэнли Х. (9 марта 2017 г.). «Регуляторные Т-лимфоциты пациентов с БАС дисфункциональны и коррелируют со скоростью и тяжестью прогрессирования заболевания». JCI Insight. 2 (5): e89530. Дои:10.1172 / jci.insight.89530. ЧВК 5333967. PMID 28289705.
- ^ Тонхофф, Джейсон Р .; Пиво, Дэвид Р .; Чжао, Вэйхуа; Плейтез, Мильвия; Симпсон, Эрика П .; Берри, Джеймс Д .; Cudkowicz, Merit E .; Аппель, Стэнли Х. (18 мая 2018 г.). «Расширенные инфузии аутологичных регуляторных Т-лимфоцитов при БАС». Неврология: нейроиммунология и нейровоспаление. 5 (4): e465. Дои:10.1212 / NXI.0000000000000465. ЧВК 5961523. PMID 29845093.
- ^ Цуда, Саяка; Накашима, Акитоши; Сима, Томоко; Сайто, Сигэру (2019). «Новая парадигма роли регуляторных Т-клеток во время беременности». Границы иммунологии. 10. Дои:10.3389 / fimmu.2019.00573. ISSN 1664-3224.
- ^ Ху, Минцзин; Эвистон, Дэвид; Сюй, Питер; Мариньо, Элиана; Чиджи, Энн; Сантнер-Нанан, Бриджит; Вонг, Калия; Ричардс, Джеймс Л .; Yap, Yu Anne; Кольер, Фиона; Куинтон, Энн (10.07.2019). «Снижение ацетата материнской сыворотки и нарушение развития тимуса плода и регуляторных Т-клеток при преэклампсии». Nature Communications. 10 (1): 3031. Дои:10.1038 / s41467-019-10703-1. ISSN 2041-1723.
- ^ Гуден М.Дж., де Бок Г.Х., Лефферс Н., Деймен Т., Нейман Х.В. (июнь 2011 г.). «Прогностическое влияние инфильтрирующих опухоль лимфоцитов на рак: систематический обзор с метаанализом». Британский журнал рака. 105 (1): 93–103. Дои:10.1038 / bjc.2011.189. ЧВК 3137407. PMID 21629244.
- ^ а б c Олейника К., Ниббс Р.Дж., Грэм Г.Дж., Фрейзер А.Р. (январь 2013 г.). «Подавление, подрыв и бегство: роль регуляторных Т-клеток в прогрессировании рака». Клиническая и экспериментальная иммунология. 171 (1): 36–45. Дои:10.1111 / j.1365-2249.2012.04657.x. ЧВК 3530093. PMID 23199321.
- ^ Плитас, Джордж; Руденский, Александр Юрьевич (09.03.2020). «Регуляторные Т-клетки при раке». Ежегодный обзор биологии рака. 4 (1): 459–477. Дои:10.1146 / annurev-Cancebio-030419-033428. ISSN 2472-3428.
- ^ а б Липпиц, Бодо Э (2013). «Цитокиновые паттерны у онкологических больных: систематический обзор». Ланцет онкологии. 14 (6): e218 – e228. Дои:10.1016 / с1470-2045 (12) 70582-х. PMID 23639322.
- ^ Марсон А., Кречмер К., Фрэмптон Г. М., Якобсен Е. С., Полански Дж. К., МакИзаак К. Д., Левин С. С., Френкель Е., фон Бёмер Н., Янг Р. А. (февраль 2007 г.). «Занятие Foxp3 и регуляция ключевых генов-мишеней во время стимуляции Т-клеток». Природа. 445 (7130): 931–5. Bibcode:2007Натура.445..931М. Дои:10.1038 / природа05478. ЧВК 3008159. PMID 17237765.
- ^ Эллис С.Д., Макговерн Дж. Л., ван Маурик А., Хоу Д., Эренштейн М. Р., Нотли, Калифорния (2014). «Индуцированные CD8 + FoxP3 + Treg-клетки при ревматоидном артрите модулируются фосфорилированием p38 и моноцитами, экспрессирующими мембранный фактор некроза опухоли α и CD86». Артрит Ревматол. 66 (10): 2694–705. Дои:10.1002 / арт.38761. PMID 24980778. S2CID 39984435.CS1 maint: несколько имен: список авторов (связь)
- ^ Wieczorek G, Asemissen A, Model F, Turbachova I, Floess S, Liebenberg V, Baron U, Stauch D, Kotsch K, Pratschke J, Hamann A, Loddenkemper C, Stein H, Volk HD, Hoffmüller U, Grützkau A, Mustea A , Huehn J, Scheibenbogen C, Olek S (январь 2009 г.). «Количественный анализ метилирования ДНК FOXP3 как новый метод подсчета регуляторных Т-клеток в периферической крови и твердых тканях». Исследования рака. 69 (2): 599–608. Дои:10.1158 / 0008-5472.CAN-08-2361. PMID 19147574.
- ^ Агарвал А., Сингх М., Чаттерджи Б.П., Чаухан А., Чакраборти А. (2014). «Взаимодействие Т-хелперов 17 с Tregs CD4 (+) CD25 (высокий) FOXP3 (+) в регуляции аллергической астмы у педиатрических пациентов». Международный журнал педиатрии. 2014: 636238. Дои:10.1155/2014/636238. ЧВК 4065696. PMID 24995020.
- ^ Лу Л.Ф., Линд Е.Ф., Гондек Д.К., Беннетт К.А., Глисон М.В., Пино-Лагос К., Скотт З.А., Койл А.Дж., Рид Дж. Л., Ван Сник Дж., Стром Т. Б., Чжэн ХХ, Ноэль Р. Дж. (Август 2006 г.). «Тучные клетки являются важными посредниками в регуляции толерантности Т-клеток». Природа. 442 (7106): 997–1002. Bibcode:2006Натура.442..997L. Дои:10.1038 / природа05010. PMID 16921386. S2CID 686654.
- ^ «Трегитоп: инструмент для иммуномодуляции». EpiVax.
- ^ Hui DJ, Basner-Tschakarjan E, Chen Y, Davidson RJ, Buchlis G, Yazicioglu M, Pien GC, Finn JD, Haurigot V, Tai A, Scott DW, Cousens LP, Zhou S, De Groot AS, Mingozzi F (сентябрь 2013 г. ). «Модуляция CD8 + Т-клеточных ответов на векторы AAV с эпитопами MHC класса II, полученными из IgG». Молекулярная терапия. 21 (9): 1727–37. Дои:10.1038 / мт.2013.166. ЧВК 3776637. PMID 23857231.
- ^ а б Де Гроот А.С., Моис Л., Макмерри Дж. А., Вамбре Е., Ван Овертвельт Л., Моингеон П., Скотт Д. В., Мартин В. (октябрь 2008 г.). «Активация естественных регуляторных Т-клеток пептидом, производным от Fc IgG» Трегитопы"". Кровь. 112 (8): 3303–11. Дои:10.1182 / кровь-2008-02-138073. ЧВК 2569179. PMID 18660382.
- ^ «Новое вливание средств NIH на сумму 2,25 миллиона долларов для Tregitope EpiVax, предложенное« лечение, изменяющее парадигму ». Жесткие биотехнологические исследования.
- ^ Су И, Росси Р., Де Гроот А.С., Скотт Д.В. (август 2013 г.). «Регуляторные Т-клеточные эпитопы (Трегитопы) в IgG индуцируют толерантность in vivo и не обладают иммуногенностью как таковой». Журнал биологии лейкоцитов. 94 (2): 377–83. Дои:10.1189 / jlb.0912441. ЧВК 3714563. PMID 23729499.
- ^ Cousens LP, Su Y, McClaine E, Li X, Terry F, Smith R, Lee J, Martin W, Scott DW, De Groot AS (2013). «Применение природных Treg-эпитопов, происходящих из IgG (трегитопов IgG), для индукции антиген-специфической толерантности на мышиной модели диабета 1 типа». Журнал исследований диабета. 2013: 1–17. Дои:10.1155/2013/621693. ЧВК 3655598. PMID 23710469.
- ^ Cousens LP, Mingozzi F, van der Marel S, Su Y, Garman R, Ferreira V, Martin W, Scott DW, De Groot AS (октябрь 2012 г.). «Обучение толерантности: новые подходы к заместительной ферментной терапии болезни Помпе». Человеческие вакцины и иммунотерапевтические препараты. 8 (10): 1459–64. Дои:10.4161 / hv.21405. ЧВК 3660767. PMID 23095864.
- ^ Cousens LP, Najafian N, Mingozzi F, Elyaman W., Mazer B, Moise L, Messitt TJ, Su Y, Sayegh M, High K, Khoury SJ, Scott DW, De Groot AS (январь 2013 г.). «Исследования in vitro и in vivo Treg-эпитопов (Tregitopes), полученных из IgG: новый многообещающий инструмент для индукции толерантности и лечения аутоиммунитета». Журнал клинической иммунологии. 33 Дополнение 1 (1): S43–9. Дои:10.1007 / s10875-012-9762-4. ЧВК 3538121. PMID 22941509.
- ^ Эльяман В., Хури С.Дж., Скотт Д.В., Де Гроот А.С. (2011). «Возможное применение трегитопов в качестве иммуномодулирующих агентов при рассеянном склерозе». Neurology Research International. 2011: 256460. Дои:10.1155/2011/256460. ЧВК 3175387. PMID 21941651.
- ^ Онлайн-менделевское наследование в человеке IPEX
- ^ ипекс в Национальные институты здравоохранения США /UW Генные тесты
внешняя ссылка
- Регуляторные + Т-клетки в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)