Proteaceae - Proteaceae
| Proteaceae | |
|---|---|
 | |
| Соцветие из Protea cynaroides | |
| Научная классификация | |
| Королевство: | Plantae |
| Clade: | Трахеофиты |
| Clade: | Покрытосеменные |
| Clade: | Eudicots |
| Заказ: | Proteales |
| Семья: | Proteaceae Юсс.[1] |
| Роды | |
Около 80 см. Текст | |
В Proteaceae /ˌпрoʊтяˈeɪsя/ площадь семья из цветущие растения преимущественно распространен в Южное полушарие. В семье 83 роды из них около 1660 известных разновидность.[2] Вместе с Platanaceae и Nelumbonaceae, они составляют заказ Proteales. Известные роды включают Protea, Banksia, Эмботриум, Гревиллея, Hakea и Макадамия. Такие виды, как варата Нового Южного Уэльса (Telopea speciosissima ), король протея (Protea cynaroides ), и различные виды Banksia, Гревиллея, и Leucadendron популярны срезанные цветы, в то время как орехи Macadamia integrifolia широко выращиваются и потребляются в коммерческих целях. Австралия и Южная Африка имеют наибольшую концентрацию разнообразия.
Этимология
Название Proteaceae от Антуан Лоран де Жюссьё в 1789 г. был основан на роде Protea, который в 1767 г. Карл Линней происходит от имени греческого бога Протей, божество, которое могло меняться между множеством форм.[3] Это подходящее изображение, поскольку семья известна своим удивительным разнообразием цветов и листьев.[нужна цитата ]
Описание

Роды Proteaceae очень разнообразны, с Banksia в частности, являясь ярким примером адаптивное излучение в растениях.[4] Эта изменчивость делает невозможным предоставление простого диагностического идентификационного ключа для семейства, хотя отдельные роды можно легко идентифицировать.
- Proteaceae варьируется от просторных кустарников до высоких лесов. деревья, высотой 40 м, обычно средней или низкой или многолетней кусты, за исключением некоторых Stirlingia виды, которые травы. Некоторые виды факультативно листопадные (Эмботрий кокцинеум ), редко остроконечные, стеблевидная часть воротника часто утолщена (лигнотуберкулез ). Indumentum из трехклеточных волосков, иногда железистые, редко отсутствуют, апикальная клетка обычно удлиненная, острая, иногда одинаково или неравномерно раздвоенная.
- Листья редко ароматные, обычно чередующиеся и по спирали, редко противоположные или мутовчатые; кожистые, редко мясистые или шиповатые, простые или сложные (непарипинатные, непарибипинатные или реже пальчатые или пальцевидные с перисто-рассеченными сегментами), весь край до (3-) перисто-рассеченного (придавая вид папоротника); редко делятся дихотомически, часто отдаленно зубчатые, зубчатые или зубчатые, сидящие или на ножках; то черешок часто с опухшим основанием, но редко с оболочкой (иногда в Синафея ), без прилистники; перистые, иногда дланчатые или параллельные жилкование, брохидодромные или сведенные к одной выступающей лопасти, вернация обычно кондуктор; анизофиллия часто возникает в разные периоды роста; листовая пластинка дорсивентральная, равносторонняя или центрированная; ткань мезофилла обычно со склеренхиматозным идиобласты, редкие секреторные полости. Брахи-парацитарный устьица (латероцитарный в Bellendena ).
Стебли растений с двумя типами радиусов: широким и многозубчатым или узким и однолучевым, флоэма стратифицированные или нет, трехлучевые узлы с тремя следы листьев (редко однолакунный с одним следом), склереиды частый; лаять с чечевицы часто увеличиваются по горизонтали, пробка камбий присутствует, обычно поверхностный. Корни боковые и короткие, часто сгруппированные в пучки (протеидные корни) с очень густыми корневыми волосками, редко с микориза.
- Растения обычно гермафродитные, реже однодомные, раздельнополые или андромодомные.
- Соцветия очень изменчивые, простые или сложные, пазушные или верхушечные, боковые цветки одиночные или парные, редко с верхушечным цветком, кистевидный, метель или конденсированный, обычно с прицветники, иногда превращаются в листья или чешуйчатые, образующие тип конуса, или с яркими цветами, образующие обертку или псевдантий, цветоносы и цветоножки иногда сокращается, уплотняется рахис, в некоторых случаях перегруженные соцветия образуют суперсоцветы (некоторые Аллоксилон ); очень редко цветки одиночные и пазушные на концах ветвей; у видов с лигнотуберами из них иногда вырастают цветы и проходят сквозь почву (геофиты).
- Цветы обычно идеальные, актиноморфные или зигоморфные, гипогинные, часто большие и эффектные. Плоские или косые, иногда образующие гинофор. Гипогинный диск присутствует, экстрастаминный или отсутствует. Околоцветник из (3-) 4 (-8) листочки околоцветника (иногда интерпретируется как димерный и дихламидный околоцветник), в 1 (-2) клапанных оборотах, иногда вытянутых в базальном мешочке, свободных или сросшихся по-разному (все слиты или даже один свободный и три базально до полностью слились) или даже попустительствуя гранично пересекающимся сосочкам, образующим трубку или двугубую структуру, зигоморфную, иногда открывающуюся латерально различными путями. Гаплостемонный андроэций, обычно изостемонный, противоположный (3-) 4 (-5) тычинки, все плодородные или некоторые превращены в стаминодии, обычно нитчатые, нити частично или полностью срослись с листочками околоцветника, реже свободны, с основанием пыльники аднатный, двустворчатый, четырехспорангиатный, иногда монокулярный и двояковыпуклая, прямая к латерному (редко), расширенная соединительнотканная, обычно с вершинкой, расхождение по продольным надрывам. Гипогинные железы (0-) 1-4, чешуйчатые или удлиненные, мясистые, свободные или сросшиеся, образующие полулунную или кольцевидную форму нектарник над сосудом. Гинецей верхний 1 (-2) апокарпный плодолистики сидячие или прилистниковые (с более или менее удлиненной гинофор ), иногда не до конца, стиль обычно развивается, клеймо небольшие или в форме терминального или субтерминального диска или даже бокового и косого, часто зубчатого, папиллярного, влажного или сухого, семяпочки 1-100 или более на плодолистик, анатропный, гемианатропный, амфитропный или ортотропный, в основном гемитропный, битегмический, крассинуцеллатный, Chalaza с кольцом сосудистых пучков, семяпровод иногда отсутствует и семяпочка срастается с плацентой, краевая плацентация с разным расположением или на вершине.
- Фрукты непристойный или непристойный, в семянка или ядро, фолликул, костянка (с одревесневшим эндокарпом) или ложно друпальным (с одревесневшим внутренним мезокарпом), иногда похожим на зерновка поскольку он срастается со стенкой яичника и семенником, часто одревесневший и серотиновый; плоды одного соцветия иногда срастаются, образуя синкарп.
- Семена 1-много, иногда крылатые, от плоских до округлых, с эндосперм отсутствует, присутствует в Беллендина, эндотеста с необычным слоем, содержащим кристаллы оксалат кальция редко отсутствует, хорошо дифференцированный эмбрион, прямые, двудольные, но часто с 3 и более (до 9) крупными семядолями, часто с ушками.
- Пыльца в монадах, треугольные в полярной проекции, (2-) 3 (-8) -апертурат, обычно изополярный и трипоратный, бипоратный в Эмботриум и племя Banksieae, кольпоидное в Beauprea, сферическая в Aulax и Франкландия или сильно анизополярный у некоторых видов Persoonia; открытие тетрад первого подчиняется закону Гарсайда.[требуется разъяснение ]
- Хромосомный номер: п = 5, 7, 10–14, 26, 28; размеры варьируются от очень маленьких (в среднем 1,0 мкм) до очень больших (в среднем 14,4 мкм) в зависимости от вида; Икс = 7, 12.
Цветы

Вообще говоря, диагностическим признаком Proteaceae является соединение Цветочная голова или, точнее, соцветие. У многих родов наиболее очевидной особенностью являются крупные и часто очень эффектные соцветия, состоящие из множества мелких цветы плотно упакованы в компактную головку или колосья. Однако даже этот признак встречается не у всех Proteaceae; Аденантос виды, например, имеют одиночные цветки. У большинства видов Proteaceae механизм опыления узкоспециализирован. Обычно это предполагает использование "пыльцы", области на стиль -конце, который представляет пыльца опылителю.[5]
Proteaceae состоит из четырех частей цветка. листочки околоцветника сплавлены в длинную узкую трубку с закрытой чашкой наверху, а нити четырех тычинки слиты с листочками околоцветника таким образом, что пыльники заключены в чашку. Первоначально пестик проходит по внутренней стороне трубки околоцветника, поэтому рыльце тоже находится внутри чашечки. По мере развития цветка пестик быстро разрастается. Поскольку рыльце защемлено, стигма должна изгибаться, чтобы удлиняться, и в конечном итоге она изгибается так далеко, что разделяет околоцветник по одному шву. Стиль продолжает развиваться, пока цветение, когда нектарники начать производить нектар. В это время околоцветник разделяется на составляющие листочки околоцветника, чашечка раскалывается, и пестик отпускается, чтобы выпрямиться более или менее вертикально.
Экология

Многие из Proteaceae специализировались протеидные корни, массы боковых корней и волосков, образующих радиальную впитывающую поверхность, образуются в слое опада листьев во время сезонного роста и обычно сморщиваются в конце сезона роста. Они являются адаптацией к росту в бедных почвах с дефицитом фосфора, значительно увеличивая доступ растений к дефицитной воде и питательным веществам за счет выделения карбоксилатов, которые мобилизуют ранее недоступный фосфор. Они также увеличивают абсорбционную поверхность корня, но это второстепенная особенность, так как это также увеличивает конкуренцию за питательные вещества с собственными корневыми кластерами.[6] Однако такая адаптация делает их очень уязвимыми для отмирания, вызванного Phytophthora cinnamomi водяная плесень и вообще нетерпимы к оплодотворение. Благодаря этим специализированным корням протеидов Proteaceae - одно из немногих семейств цветковых растений, которые не образуют симбиоза с арбускулярный микоризный грибы. Они выделяют большое количество органических кислот (лимонная кислота и яблочная кислота ) каждые 2–3 дня, чтобы способствовать мобилизации и абсорбции фосфатов. Многие виды приспособлены к огню (пирофиты ), что означает, что у них есть стратегии выживания в пожарах, охватывающих их среду обитания. Некоторые респираторы, и иметь толстый подвой, закопанный в землю, который дает новые стебли после пожара, а другие исследователи Это означает, что взрослые растения погибают от огня, но рассыпают свои семена, которые стимулируются дымом, чтобы пустить корни и прорасти. Ранее считалось, что тепло стимулирует рост, но теперь было показано, что химические вещества в дыме вызывают его.
Выделяют четыре раздельнополых рода (Aulax, Дилобея, Гелиопсис и Leucadendron ), У 11 родов андромоноэцитов и некоторых других родов есть виды, которые являются криптическими андромоноэцитами: два вида бесплодны и размножаются только вегетативно (Ломатия тасманика, Hakea pulvinifera). Виды различаются между автосовместимостью и автонесовместимостью с промежуточными ситуациями; такие ситуации иногда возникают у одного и того же вида. Цветки обычно выступающие. Непосредственно перед цветением пыльники выпускают свои пыльца, откладывая его на рыльце, которое во многих случаях имеет увеличенную мясистую область специально для отложения собственной пыльцы. Нектародобывающие вряд ли соприкоснутся с самими пыльниками, но вряд ли могут избежать контакта с рыльцем; таким образом, стигма действует как пыльца, гарантируя, что питатели нектара действуют как опылители. Обратной стороной этой стратегии опыления является то, что вероятность самоопыления значительно увеличивается; многие Proteaceae противодействуют этому с помощью таких стратегий, как протандрия, несовместимость с самими собой или преимущественное прерывание самоопыления семян. Системы представления пыльцы обычно очень разнообразны, что соответствует разнообразию опылителей. Опыление осуществляется пчелы, жуки, мухи, моль, птицы (медоеды, солнечные птицы, сахарные птицы и колибри ) и млекопитающих (грызуны, мелкие сумчатые, слоновьи землеройки и летучие мыши. Последние два средства были эволюционно выведены из энтомофилия в разных, независимых мероприятиях. Распространение некоторых видов демонстрирует любопытный феномен серотины, что связано с их пирофитным поведением: эти деревья накапливают на своих ветвях плоды, внешние слои или защитные структуры которых (прицветники ) сильно одревесневшие и огнестойкие. Плоды высвобождают семена только после того, как они сгорели и когда земля удобрена золой от огня и свободна от конкурентов. Многие виды имеют семена с элайосомы которые рассредоточены муравьи; Семена с крыльями или чертополох выставляют анемохория, в то время как костянки и другие мясистые плоды демонстрируют эндозоохория поскольку их глотают млекопитающие и птицы. Известно, что некоторые африканские и австралийские грызуны накапливают плоды и семена этих растений в своих гнездах, чтобы питаться ими, хотя некоторым удается прорасти.
Распределение
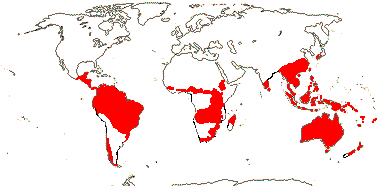
Proteaceae - это в основном семейство южного полушария с основными центрами разнообразия в Австралии и Южной Африке. Встречается также в Центральной Африке, юг и Центральная Америка, Индия, восточная и юго-восточная Азия, и Океания.[6] Только два вида известны из Новой Зеландии, хотя свидетельства ископаемой пыльцы предполагают, что раньше их было больше.[7]
Это хороший пример Гондванец семейство, таксоны встречаются практически на каждом участке суши, считающемся остатком древних суперконтинент Гондвана, кроме Антарктида. Считается, что семья и подсемейства диверсифицировались задолго до фрагментации Гондваны, а это значит, что всем им более 90 миллионов лет. Доказательства этого включают множество протеиновых пыльца найдено в Меловой каменный уголь депозиты Южный остров из Новая Зеландия. Считается, что своего нынешнего распространения он получил в основном благодаря Континентальный дрифт а не рассредоточение через океанские пропасти.[8]
Фитохимия

Никаких убедительных исследований химических веществ, присутствующих в этом большом семействе, не проводилось. Роды Protea и Faurea необычны, поскольку они используют ксилоза в качестве основного сахара в их нектаре и поскольку они имеют высокую концентрацию полигалактола, в то время как сахароза это основной сахар, присутствующий в Гревиллея. Цианогенные гликозиды, происходит от тирозин, часто присутствуют, как и проантоцианидины (дельфинидин и цианидин ), флавонолы (кемпферол, кверцетин и мирицетин ) и арбутин. Алкалоиды обычно отсутствуют. Иридоиды и эллаговая кислота также отсутствуют. Сапонины и сапогенины могут присутствовать или отсутствовать у разных видов. Накапливаются многие виды алюминий.

Использование и выращивание

Многие традиционные культуры использовали Proteaceae в качестве корма, лекарства, для обработки шкур животных, в качестве источника красителей, дров и древесины для строительства. Австралийские аборигены едят плоды Persoonia, а также семена видов других родов, в том числе Гевуина и Макадамия, являются частью рациона коренных народов, но также продаются по всему миру. Нежные ростки Helicia виды используются на Яве, а нектар из соцветий ряда видов пьют в Австралии. Традиционные лекарства могут быть получены из настоев корней, коры, листьев или цветов многих видов, которые используются в качестве местного применения при кожных заболеваниях или для внутреннего применения в качестве тонизирующих средств, афродизиаков и галактогенов для лечения головных болей, кашля, дизентерии, диареи, несварения желудка, язвы желудка и болезни почек. Древесина деревьев этого семейства широко используется в строительстве и для внутренних работ, например, для украшения; дерево из пород Protea, Leucadendron и Гревиллея особенно популярен. Много разновидность используются в садоводстве, особенно виды Banksia, Эмботриум, Гревиллея, и Телопея. К сожалению, это использование привело к появлению экзотических видов, которые стали инвазивными; примеры включают иву хакея (Хакея салицифолистная ) и шелковистой хакеи (Hakea sericea ) в Португалии. Два вида Макадамия выращиваются в коммерческих целях ради съедобных орехов.
Гевуина авельяна (Чилийский лещина) выращивают ради съедобных орехов в Чили и Новая Зеландия, и они также используются в фармацевтической промышленности из-за их увлажняющих свойств и в качестве ингредиента в солнцезащитные кремы. Это самое устойчивое к холоду из семейств деревьев, производящих орехи. Его также высаживают в Британские острова и на тихоокеанском побережье Соединенные Штаты за его тропический вид и способность расти в более прохладный климат.
Многие виды Proteaceae культивируются питомник промышленность в качестве барьерных растений и их выдающихся и отличительных цветов и листвы. Некоторые виды важны для срезанная цветочная промышленность, особенно некоторые Banksia и Protea разновидность. Два вида рода Макадамия выращиваются в коммерческих целях для получения съедобных орехов.
Сахарные кусты (Protea ), подушечки для булавок (Лейкоспермум ) и шишки (Leucadendron ), а также другие подобные пагоды (Мимет ), Aulax и краснеющие невесты (Серрурия ), составляют одну из трех основных групп растений финбос, который является частью Мыс Цветочное Королевство, самое маленькое, но самое богатое царство растений для своего размера и единственное царство, находящееся в пределах одной страны. Другими основными группами растений финбоса являются Вересковые и Restionaceae. Таким образом, южноафриканские протеи широко культивируются из-за их разнообразных форм и необычных цветов. Они популярны в Южной Африке своей красотой и полезностью в сады дикой природы для привлечения птиц и полезных насекомых.
В качестве декоративных растений наиболее ценятся деревья, растущие в южных широтах, так как они создают ландшафты в умеренный климат тропический вид; Lomatia ferruginea (Fuinque), Lomatia Hirsuta (Radal) были введены в Западной Европа и на запад Соединенные Штаты. Эмботрий кокцинеум (Чилийский Firetree или Notro) высоко ценится на Британских островах за свои темно-красные цветы и может быть найден на севере до Фарерские острова на широте 62 ° северной широты.
Среди Банкиас, многие из которых произрастают в умеренном и средиземноморском климате, подавляющее большинство - кустарники; лишь некоторые из них ценятся за свою высоту. Среди самых высоких видов: Б. Integrifolia со своими подвидами Б. Integrifolia subsp. Монтикола, который примечателен тем, что растения, входящие в подвид, являются самыми высокими деревьями банксии и более морозостойки, чем другие банки, Б. Семинуда, Б. литоралис, B. serrata; среди тех, что можно считать небольшими деревьями или большими кустарниками: B. grandis, Б. прионоты, B. marginata, B. coccinea и Б. speciosa; все они из-за своего размера высажены в парках и садах и даже вдоль дорог. Остальные виды этого рода, около 170 видов, являются кустарниками, хотя некоторые из них ценятся за свои цветы.
Другой вид, который культивируется в некоторых частях света, хотя и меньше по размеру, - это Telopea speciosissima (Варатах), с гор Новый Южный Уэльс, Австралия.
Некоторые виды с умеренным климатом выращиваются в Австралии из-за их привлекательного внешнего вида: Persoonia pinifolia (сосноволистный гибунг ) ценится за ярко-желтые цветы и плоды, напоминающие виноград. Аденантос серцеус (пушистый куст) выращивают из-за его привлекательных мягких листьев и маленьких красных или оранжевых цветов. Hicksbeachia pinnatifolia (говяжий орех, красный орех) обычно сажают из-за листвы и съедобных орехов.
Паразиты

Proteaceae особенно восприимчивы к определенным паразитам, в частности к паразитам. оомицет Phytophthora cinnamomi, которая вызывает сильную корневую гниль у растений, произрастающих в средиземноморском климате. Fusarium oxysporum вызывает заболевание корней, называемое фузариозом, которое вызывает пожелтение и увядание с серьезным экологическим ущербом для лесных растений и экономическим ущербом для растений, представляющих коммерческий интерес. Другие распространенные инфекции вызываются видами Ботриосфаерия, Ризоктония, Армиллярия, Ботритис, Калонектрия и другие грибы.
Статус сохранения
В МСОП[9] считает, что 47 видов Proteaceae находятся под угрозой исчезновения, из которых один вид, Stenocarpus dumbeensis Guillaumin, 1935, из Новой Каледонии, считается вымершим. Виды этого семейства особенно подвержены разрушению или фрагментации своих среда обитания, пожар, паразитарные болезни, конкуренция из интродуцированных растений, деградация почвы и другой ущерб, причиненный людьми и их домашними животными. Вид также подвержен влиянию изменение климата.
Окаменелости

Proteaceae имеет богатую летопись окаменелостей, несмотря на присущие им трудности в идентификации останков, не показывающих диагностических характеристик. Идентификация обычно происходит по сочетанию брахи-парацитарных устьиц и необычного трихома основания или, в других случаях, необычное строение пыльцевых тетрад. Окаменелости, относящиеся к этому семейству, были найдены на большинстве территорий, которые сформировали Гондвана суперконтинент. Широкое разнообразие пыльцы этого семейства, восходящее к Верхнему Меловой (Кампанский -Маастрихтский ) с юго-востока Австралии и пыльца из среднего мела (Сеноманский -Туронский ) из Северной Африки и Перу описывается как Triorites africaensis. Первые макрофоссилии появляются двадцать миллионов лет спустя в Палеоцен Южной Америки и северо-востока Австралии. Летопись окаменелостей в некоторых областях, таких как Новая Зеландия и Тасмания, показывает большее биоразнообразие Proteaceae, чем существует в настоящее время, что подтверждает тот факт, что распространение многих таксонов резко изменилось с течением времени и что семейство пострадало от общей упадка, включая высокие уровни исчезновения во время Кайнозойский.
Таксономия

Впервые описан французским ботаником Антуан Лоран де Жюссьё, семейство Proteaceae - довольно большое, насчитывающее около 80 родов, но менее 2000 видов. Это признают практически все систематики. Прочно утвердился в Линнеевская таксономия, это также признается кладистика -основан ПНГ и APG II системы. Ставится в заказ Proteales, чье размещение менялось.
Классификация родов Proteaceae была произведена Лори Джонсон и Барбара Бриггс[10] в своей влиятельной монографии 1975 г. "О Proteaceae: эволюция и классификация южного семейства ",[11] пока он не был в значительной степени вытеснен молекулярными исследованиями Питера Х. Вестона и Найджела Баркера в 2006 году. Proteaceae теперь разделен на пять подсемейств: Bellendenoideae, Persoonioideae, Symphionematoideae, Proteoideae и Grevilleoideae.[11] В 2008 году Маст и его коллеги обновили Макадамия и родственные роды в трибе Macadamieae. Более того, Орит мегакарпус было обнаружено, что не принадлежит к роду Оритс, ни в трибе Roupaleae, а в трибе Macadamieae, отсюда и новое название вида Nothorites megacarpus.[12] Полная договоренность, согласно Weston and Barker (2006) с обновлениями родов от Mast et al. (2008), выглядит следующим образом:

- Семья Proteaceae
- Подсемейство Bellendenoideae
- Подсемейство Persoonioideae
- Племя Placospermeae
- Племя Persoonieae
- Подсемейство Symphionematoideae
- Подсемейство Proteoideae
- incertae sedis
- Эйдотея — Beauprea — Бопреопсис — Дилобея — Кенаррен — Франкландия
- Племя Conospermeae
- Subtribe Stirlingiinae
- Subtribe Conosperminae
- Племя Petrophileae
- Племя Простейшие
- Племя Leucadendreae
- Subtribe Isopogoninae
- Subtribe Аденантины
- Subtribe Leucadendrinae
- Leucadendron — Серрурия — Параномус — Vexatorella — Сороцефал — Spatalla — Лейкоспермум — Мимет — Диастелла — Оротамнус
- incertae sedis
- Подсемейство Grevilleoideae
- incertae sedis
- Племя Roupaleae
- incertae sedis
- Subtribe Roupalinae
- Subtribe Lambertiinae
- Subtribe Heliciinae
- Subtribe Floydiinae
- Племя Banksieae
- Subtribe Musgraveinae
- Subtribe Banksiinae
- Племя Embothrieae
- Subtribe Lomatiinae
- Subtribe Embothriinae
- Эмботриум — Oreocallis — Аллоксилон — Телопея
- Subtribe Stenocarpinae
- Subtribe Hakeinae
- Опистиолепис — Букингемия — Hakea — Гревиллея — Finschia
- Племя Macadamieae
- Subtribe Macadamiinae
- Subtribe Малагасийны
- Subtribe Virotiinae
- Subtribe Gevuininae
- Кардвеллия — Слюмеродендрон — Euplassa — Гевуина — Bleasdalea — Hicksbeachia — Kermadecia — Turrillia
Рекомендации
- ^ Группа филогении покрытосеменных (2009). «Обновленная классификация филогенетической группы покрытосеменных для порядков и семейств цветковых растений: APG III». Ботанический журнал Линнеевского общества. 161 (2): 105–121. Дои:10.1111 / j.1095-8339.2009.00996.x.
- ^ Кристенхус, М. Дж. М. и Бинг, Дж. У. (2016). «Число известных видов растений в мире и его ежегодный прирост». Фитотакса. 261 (3): 201–217. Дои:10.11646 / phytotaxa.261.3.1.
- ^ Жасси, Антуан Лоран де, Genera plantarum secundum ordines naturales disposita, juxta methodum в Horto Regio Parisiensi exaratum, anno M.DCC.LXXIV. Париж, 1789 год. стр. 78-80.
- ^ Маст, А. Р. и Гивниш, Т. Дж. (2002). «Историческая биогеография и происхождение устьичных распределений у Banksia и Dryandra (Proteaceae) на основе их филогении хпДНК». Американский журнал ботаники. 89 (8): 1311–1323. Дои:10.3732 / ajb.89.8.1311. PMID 21665734. S2CID 2835600.
- ^ Уотсон, Л. и Даллвиц, М. Дж. (1992). "Proteaceae". Семейства цветковых растений: описания, иллюстрации, идентификация, поиск информации. Получено 26 июн 2006.
- ^ а б Орчард, Энтони Э. (ред.). "Proteaceae". Флора Австралии, том 16: Elaeagnaceae, Proteaceae 1. Мельбурн: Австралийское исследование биологических ресурсов / публикация CSIRO. Архивировано из оригинал 31 октября 2006 г.. Получено 26 июн 2006.CS1 maint: дополнительный текст: список авторов (связь)
- ^ Полюс М (1998). «Рекорд Proteaceae в Новой Зеландии». Австралийская систематическая ботаника. 11 (4): 343–372. Дои:10.1071 / SB97019.
- ^ Уэстон, П. Х. и Крисп, М. Д. (1996). «Транстихоокеанские биогеографические закономерности в Proteaceae». В Кисте А.И Миллер, С. Э. (ред.). Происхождение и эволюция биоты островов Тихого океана, от Новой Гвинеи до восточной Полинезии: закономерности и процессы. Амстердам: SPB Academic Publishing. С. 215–232. ISBN 978-90-5103-136-2.
- ^ МСОП 2006 г. Красный список МСОП 2006 г., находящихся под угрозой исчезновения.
. Загружено 22 февраля 2007 г. - ^ Л. А. С. Джонсон и Бриггс, Б.Г. (1975). «О Proteaceae: эволюция и классификация южного семейства». Журнал Линнеевского общества Лондона. Ботаника. 70 (2): 83–182. Дои:10.1111 / j.1095-8339.1975.tb01644.x.
- ^ а б Уэстон, Питер Х .; Баркер, Найджел П. (2006). «Новая надродовая классификация Proteaceae с аннотированным списком родов» (PDF). Телопея. 11 (3): 314–44. Дои:10.7751 / telopea20065733. Получено 30 марта 2010.
- ^ Mast, Austin R .; Willis, Crystal L .; Джонс, Эрик H .; Даунс, Кэтрин М .; Уэстон, Питер Х. (июль 2008 г.). "Меньший Макадамия от более блуждающего племени: вывод о филогенетических отношениях, временах расхождения и эволюции диаспор в Макадамия и родственники (триба Macadamieae; Proteaceae) ". Американский журнал ботаники. 95 (7): 843–870. Дои:10.3732 / ajb.0700006. ISSN 1537-2197. PMID 21632410. Получено 4 апреля 2013.
- Уэстон, П. (2007). «Proteaceae». Кубицки К. (Редактор). Семейства и роды сосудистых растений. IX. Цветущие растения - Eudicots. Springer-Verlag: Берлин. ISBN 978-3-540-32214-6.
- Хут, С. И Дуглас, А. (1998). «Филогения Proteaceae на основе атпГруппа атпB-rbcПоследовательности межгенной спейсерной области L ". Австралийская систематическая ботаника. 11 (4): 301–320. Дои:10.1071 / sb98027.
- Рэмси, H.P .. (1963). «Число хромосом у протейных». Австралийский журнал ботаники. 11 (1): 1–20. Дои:10.1071 / BT9630001.
- Уотсон, Л. и Даллвиц, М.Дж. (1992). «Семейства цветковых растений: описания, иллюстрации, идентификация и поиск информации. Версия: 29 июля 2006 г.». Получено 31 января 2007.
- Браун, Р. О Proteaceae Жасси Труды Линнеевского общества 10:15-216.
внешняя ссылка
 Данные, относящиеся к Proteaceae в Wikispecies
Данные, относящиеся к Proteaceae в Wikispecies СМИ, связанные с Proteaceae в Wikimedia Commons
СМИ, связанные с Proteaceae в Wikimedia Commons- Изображения Proteaceae из Австралийского национального ботанического сада
- карта
{kind=link}