РНК-полимераза II - RNA polymerase II
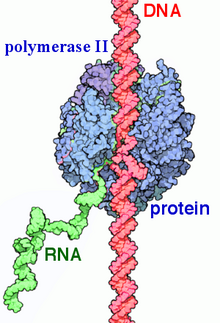
РНК-полимераза II (RNAP II и Pol II) это мультипротеиновый комплекс который расшифровывает ДНК в предшественников информационная РНК (мРНК) и большинство малая ядерная РНК (мяРНК) и микроРНК.[1][2] Это один из трех RNAP ферменты найден в ядре эукариотический клетки.[3] A 550 кДа комплекса из 12 субъединиц, РНКП II является наиболее изученным типом РНК-полимераза. Широкий спектр факторы транскрипции необходимы для связывания с вышестоящим геном промоутеры и начать транскрипцию.
Открытие

Ранние исследования предполагали наличие как минимум двух РНКП: один, который синтезировал рРНК в ядрышко, и тот, который синтезировал другую РНК в нуклеоплазма, часть ядра, но вне ядрышка.[5] В 1969 году ученые-экспериментаторы Роберт Рёдер и Уильям Раттер окончательно обнаружил дополнительную РНКП, которая отвечает за транскрипцию некоторого вида РНК в нуклеоплазме. Вывод был получен с использованием ионообменная хроматография через DEAE покрытый Сефадекс бусы. Методика разделяла ферменты в порядке соответствующих элюций,, ΙΙ,, путем увеличения концентрации сульфата аммония. Ферменты были названы в соответствии с порядком элюирования, RNAP I, RNAP II, RNAP IΙI.[3] Это открытие продемонстрировало, что в нуклеоплазме присутствует дополнительный фермент, который позволяет дифференцировать RNAP II и RNAP III.
Подразделения

В эукариотический Центральная РНК-полимераза II была сначала очищена с использованием анализов транскрипции.[7] Очищенный фермент обычно имеет 10–12 субъединиц (12 у человека и дрожжей) и не способен распознавать специфический промотор.[8] Известно много взаимодействий субъединица-субъединица.[9]
- ДНК-направленная субъединица РНК-полимеразы II RPB1 - ан фермент что у людей кодируется POLR2A ген а в дрожжах кодируется RPO21. RPB1 - самая крупная субъединица РНК-полимеразы II. Он содержит карбокси-концевой домен (CTD) состоит из 52 гептапептидных повторов (YSPTSPS), которые необходимы для активности полимеразы.[10] CTD был впервые обнаружен в лаборатории C.J. Ingles в Университете Торонто и JL Corden в Университет Джона Хопкинса. В сочетании с несколькими другими субъединицами полимеразы субъединица RPB1 образует ДНК-связывающий домен полимеразы, бороздку, в которой находится матрица ДНК. записано в РНК.[11] Он сильно взаимодействует с RPB8.[9]
- RPB2 (POLR2B ) - вторая по величине субъединица, которая в сочетании по крайней мере с двумя другими субъединицами полимеразы образует структуру внутри полимеразы, которая поддерживает контакт в активном сайте фермента между матрицей ДНК и вновь синтезированной РНК.[12]
- RPB3 (POLR2C ) - третья по величине субъединица. Существует как гетеродимер с другой субъединицей полимеразы, POLR2J формирование основного узла. RPB3 сильно взаимодействует с RPB1-5, 7, 10–12.[9]
- Субъединица B4 РНК-полимеразы II (RPB4) - закодировано POLR2D ген[13] является четвертой по величине субъединицей и может выполнять функцию защиты от стресса.
- RPB5 - У человека кодируется POLR2E ген. Две молекулы этой субъединицы присутствуют в каждой РНК-полимеразе II.[14] RPB5 сильно взаимодействует с RPB1, RPB3 и RPB6.[9]
- RPB6 (POLR2F ) - образует структуру по крайней мере с двумя другими субъединицами, которая стабилизирует транскрибирующую полимеразу на матрице ДНК.[15]
- RPB7 - закодировано POLR2G и может играть роль в регуляции функции полимеразы.[16] RPB7 сильно взаимодействует с RPB1 и RPB5.[9]
- RPB8 (POLR2H ) - взаимодействует с субъединицами RPB1-3, 5 и 7.[9]
- RPB9 - Канавка, в которой матрица ДНК транскрибируется в РНК, состоит из RPB9 (POLR2I ) и RPB1.
- RPB10 - продукт гена POLR2L. Он взаимодействует с RPB1-3 и 5 и сильно с RPB3.[9]
- RPB11 - субъединица RPB11 у человека состоит из трех субъединиц: POLR2J (RPB11-a), POLR2J2 (RPB11-b) и POLR2J3[17] (RPB11-c).
- RPB12 - Также с RPB3 взаимодействует RPB12 (POLR2K ).[9]
сборка
RPB3 участвует в сборке РНК-полимеразы II.[18] Субкомплекс RPB2 и RPB3 появляется вскоре после синтеза субъединицы.[18] Этот комплекс впоследствии взаимодействует с RPB1.[18] RPB3, RPB5 и RPB7 взаимодействуют друг с другом с образованием гомодимеров, а RPB3 и RPB5 вместе могут связываться со всеми другими субъединицами RPB, кроме RPB9.[9] Только RPB1 прочно связывается с RPB5.[9] Субблок RPB1 также связывается с RPB7, RPB10 и более слабо, но наиболее эффективно с RPB8.[9] Как только RPB1 входит в комплекс, могут войти другие субъединицы, такие как RPB5 и RPB7, где RPB5 связывается с RPB6 и RPB8, а RPB3 вводит RPB10, RPB 11 и RPB12.[9] RPB4 и RPB9 могут войти, как только большая часть комплекса будет собрана. RPB4 образует комплекс с RPB7.[9]
Кинетика
Ферменты может катализировать до нескольких миллионов реакций в секунду. Нормы содержания ферментов зависят от условий раствора и концентрации субстрата. Как и другие ферменты, POLR2 имеет кривую насыщения и максимальную скорость (VМаксимум). Оно имеет Kм (концентрация субстрата требуется на половину VМаксимум) и kКот (количество молекул субстрата, обрабатываемых одним активным центром в секунду). Константа специфичности определяется выражением kКот/Kм. Теоретический максимум для константы специфичности - это предел диффузии около 108 до 109 (M−1s−1), где каждое столкновение фермента с его субстратом приводит к катализу. У дрожжей мутация в домене триггерной петли наибольшей субъединицы может изменить кинетику фермента.[19]
Бактериальная РНК-полимераза, родственница РНК-полимеразы II, переключается между инактивированным и активированным состояниями, перемещаясь назад и вперед вдоль ДНК.[20] Концентрации [NTP]экв = 10 мкМ GTP, 10 мкМ UTP, 5 мкМ ATP и 2,5 мкМ CTP, дают среднюю скорость элонгации, число оборотов ~ 1 п.н. (NTP)−1 для бактериальной РНКП, родственника РНК-полимеразы II.[20]

РНК-полимераза II претерпевает обширную котранскрипционную паузу во время элонгации транскрипции.[21][22] Эта пауза особенно выражена в нуклеосомах и частично возникает из-за того, что полимераза входит в транскрипционно некомпетентное состояние обратного отслеживания.[21] Продолжительность этих пауз колеблется от секунд до минут или дольше, и выходу из долгоживущих пауз могут способствовать факторы удлинения, такие как TFIIS.[23] В свою очередь, скорость транскрипции влияет на то, вытесняются ли гистоны транскрибируемых нуклеосом из хроматина или повторно вставляются за транскрибирующей полимеразой.[24]
Альфа-Аманитин
РНК-полимераза II ингибируется α-Аманитин[25] и другие аматоксины. α-Аманитин - очень ядовитое вещество, которое содержится во многих грибах.[5] Грибной яд по-разному влияет на каждую из РНК-полимераз: I, II, III. RNAP I полностью не реагирует на вещество и будет функционировать нормально, в то время как RNAP III имеет умеренную чувствительность. Однако RNAP II полностью подавляется токсином. Альфа-аманитин ингибирует RNAP II за счет сильного взаимодействия в «воронке», «щели» и ключевом «мостике» фермента. α-спираль "области субъединицы RPB-1.[26]
Холоэнзим
Холофермент РНК-полимеразы II это форма эукариотический РНК-полимераза II, которая рекрутируется на промоторы белок -кодирующие гены в живых клетках.[8] Он состоит из РНК-полимеразы II, подмножества общих факторы транскрипции, и регуляторные белки известные как белки SRB.
Часть сборки холофермента упоминается как преинициативный комплекс, поскольку его сборка происходит на ген промоутер до начала транскрипция. В посреднический комплекс действует как мост между РНК-полимеразой II и факторами транскрипции.
Контроль по структуре хроматина
Это схема примерного механизма дрожжевых клеток, с помощью которого хроматин структура и гистон посттрансляционная модификация помочь регулировать и записывать транскрипция из гены РНК-полимеразой II.
Этот путь дает примеры регуляции в следующих точках транскрипции:
- Предварительная инициация (продвижение Bre1, модификация гистона)
- Инициирование (продвижение посредством TFIIH, модификация Pol II И продвижение посредством КОМПАСА, модификация гистона)
- Удлинение (продвижение Set2, модификация гистонов)
Обратите внимание, что это относится к различным этапам процесса как к этапам регулирования. Не было доказано, что они используются для регулирования, но весьма вероятно, что это так.
Промоторы элонгации РНК Pol II можно разделить на 3 класса.
- Факторы, влияющие на арест лекарственного средства / последовательности (различные интерферирующие белки)
- Факторы, ориентированные на структуру хроматина (посттранскрипционные модификаторы гистонов, например, метилтрансферазы гистонов)
- Факторы, улучшающие катализ РНК Pol II (различные мешающие белки и кофакторы Pol II; см. РНК-полимераза II).
Механизмы транскрипции
- Факторы, ориентированные на структуру хроматина:
(HMTs (ЧАСistone MэтилТрансферазы)):
КОМПАС§ † - (COMсплетение пroteins В КАЧЕСТВЕобщался с Set1) - Метилирует лизин 4 гистона H3: отвечает за репрессию / подавление транскрипции. Нормальная часть регуляции роста клеток и транскрипции в RNAP II.[27] - Set2 - метилирует лизин 36 гистона H3: Set2 участвует в регуляции элонгации транскрипции посредством прямого контакта с CTD.[28]
(интересный не относящийся к делу пример: Dot1 * ‡ - метилирует лизин 79 гистона H3.) - Bre1 - Убихинаты (добавляет убиквитин к) лизин 123 гистона H2B. Связан с пре-инициацией и позволяет связываться с РНК Pol II.
CTD РНК-полимеразы
С-конец RPB1 присоединяется для образования С-концевого домена (CTD). Карбоксиконцевой домен РНК-полимеразы II обычно состоит из 52 повторов последовательности Tyr-Ser-Pro-Thr-Ser-Pro-Ser.[29] Домен простирается от ядра фермента RNAPII до выходного канала, это размещение эффективно из-за индукции «реакций процессинга РНК посредством прямого или косвенного взаимодействия с компонентами механизма процессинга РНК».[30] Домен CTD не существует в РНК-полимеразе I или РНК-полимеразе III.[3] CTD РНК-полимеразы был впервые обнаружен в лаборатории К. Дж. Инглза в Университете Торонто, а также в лаборатории Дж. Кордена в Университете Джона Хопкинса во время процессов секвенирования ДНК, кодирующей субъединицу RPB1 РНК-полимеразы дрожжей и мышей соответственно. Другие белки часто связывают С-концевой домен РНК-полимеразы, чтобы активировать полимеразную активность. Именно белковый домен участвует в инициация транскрипции, укупорка из Транскрипт РНК, и привязанность к сплайсосома за Сплайсинг РНК.[10]
Фосфорилирование домена CTD
РНК-полимераза II существует в двух нефосфорилированных и фосфорилированных формах, IIA и IIO соответственно.[5][3] Переход между двумя формами способствует различным функциям транскрипции. В фосфорилирование CTD катализируется одним из шести общие факторы транскрипции, TFIIH. TFIIH служит двум целям: первая - раскручивать ДНК в стартовом сайте транскрипции, а вторая - фосфорилировать. Полимераза формы IIA присоединяется к преинициативному комплексу, это предполагается, потому что IIA связывается с более высоким сродством с TBP (Связывающий белок ТАТА-бокса ), субъединица общего фактора транскрипции TFIID, чем форма полимеразы IIO. Полимераза формы IIO способствует удлинению цепи РНК.[5] Метод инициации элонгации осуществляется путем фосфорилирования Серин в положении 5 (Ser5) через TFIIH. Вновь фосфорилированный Ser5 рекрутирует ферменты, которые блокируют 5'-конец вновь синтезированной РНК, и факторы процессинга «3», чтобы поли (А) места".[30] Как только второй серин фосфорилируется, Ser2, элонгация активируется. Для прекращения удлинения должно произойти дефосфорилирование. После того, как домен полностью дефосфорилирован, фермент RNAP II «возвращается в цикл» и катализирует тот же процесс с другим сайтом инициации.[30]
Рекомбинационная репарация, связанная с транскрипцией
Окислительное повреждение ДНК может блокировать РНК-полимеразу II транскрипция и вызвать обрыв пряди. Был описан процесс рекомбинации, связанный с транскрипцией, который может защитить от повреждения ДНК.[31] На этапах G1 / G0 клеточный цикл, клетки демонстрируют сборку гомологичных факторов рекомбинации на двухцепочечных разрывах в активно транскрибируемых областях. Похоже, что транскрипция связана с репарацией двухцепочечных разрывов ДНК с помощью шаблонной гомологичной рекомбинации РНК. Этот процесс ремонта эффективно и точно восстанавливает двухнитевые разрывы в гены активно транскрибируется РНК-полимеразой II.
Смотрите также
- Эукариотическая транскрипция
- Посттранскрипционная модификация
- РНК-полимераза I
- Холофермент РНК-полимеразы II
- РНК-полимераза III
- Транскрипция (генетика)
Рекомендации
- ^ Корнберг Р.Д. (декабрь 1999 г.). «Контроль транскрипции эукариот». Тенденции в клеточной биологии. 9 (12): M46–9. Дои:10.1016 / S0962-8924 (99) 01679-7. PMID 10611681.
- ^ Симс Р.Дж., Мандал С.С., Рейнберг Д. (июнь 2004 г.). «Последние основные моменты транскрипции, опосредованной РНК-полимеразой-II». Текущее мнение в области клеточной биологии. 16 (3): 263–71. Дои:10.1016 / j.ceb.2004.04.004. PMID 15145350.
- ^ а б c d Янг, Ричард А. (28 ноября 2003 г.). «РНК-полимераза II». Ежегодный обзор биохимии. 60 (1): 689–715. Дои:10.1146 / annurev.bi.60.070191.003353. PMID 1883205.
- ^ Meyer PA, Ye P, Zhang M, Suh MH, Fu J (июнь 2006 г.). «Фазирование РНК-полимеразы II с использованием внутренне связанных атомов Zn: обновленная структурная модель». Структура. 14 (6): 973–82. Дои:10.1016 / j.str.2006.04.003. PMID 16765890.
- ^ а б c d 1942-, Уивер, Роберт Франклин (01.01.2012). Молекулярная биология. Макгроу-Хилл. ISBN 9780073525327. OCLC 789601172.CS1 maint: числовые имена: список авторов (связь)
- ^ Армаш, Карим-Жан; Миттервегер, Симона; Мейнхарт, Антон; Крамер, Патрик (2019). «Структуры полной РНК-полимеразы II и ее субкомплекса, Rpb4 / 7» (PDF). Журнал биологической химии. 280 (8): 7131–1734. Дои:10.2210 / pdb1wcm / PDB. PMID 15591044.
- ^ Савадого М., Сентенак А (1990). «РНК-полимераза B (II) и общие факторы транскрипции». Ежегодный обзор биохимии. 59: 711–54. Дои:10.1146 / annurev.bi.59.070190.003431. PMID 2197989.
- ^ а б Майер В.Е., Янг Р.А. (октябрь 1998 г.). «Холоферменты и субкомплексы РНК-полимеразы II». Журнал биологической химии. 273 (43): 27757–60. Дои:10.1074 / jbc.273.43.27757. PMID 9774381.
- ^ а б c d е ж грамм час я j k л м Акер Дж., Де Грааф М., Чейнел И., Хазак В., Кединджер С., Виньерон М. (июль 1997 г.). «Взаимодействие между субъединицами РНК-полимеразы II человека». Журнал биологической химии. 272 (27): 16815–21. Дои:10.1074 / jbc.272.27.16815. PMID 9201987.
- ^ а б Брикки В.Дж., Гринлиф А.Л. (июнь 1995 г.). «Функциональные исследования карбоксиконцевого повторяющегося домена РНК-полимеразы II дрозофилы in vivo». Генетика. 140 (2): 599–613. ЧВК 1206638. PMID 7498740.
- ^ «Ген Entrez: полипептид А полимеразы POLR2A (РНК) II (ДНК-направленный), 220 кДа».
- ^ "Ген Entrez: полипептид B полимеразы (РНК) II (ДНК-направленный) POLR2B, 140 кДа".
- ^ Khazak V, Estojak J, Cho H, Majors J, Sonoda G, Testa JR, Golemis EA (апрель 1998 г.). «Анализ взаимодействия новой субъединицы РНК-полимеразы II (pol II) hsRPB4 с ее партнером hsRPB7 и с pol II». Молекулярная и клеточная биология. 18 (4): 1935–45. Дои:10.1128 / mcb.18.4.1935. ЧВК 121423. PMID 9528765.
- ^ «Ген Entrez: полипептид E полимеразы POLR2E (РНК) II (ДНК-направленный), 25 кДа».
- ^ «Ген Entrez: полипептид F полимеразы (РНК) II (ДНК-направленный) POLR2F».
- ^ «Ген Entrez: полипептид G полимеразы (РНК) II (ДНК-направленный) POLR2G».
- ^ «Полипептид J3 полимеразы (РНК) II (ДНК-направленный) POLR2J3».
- ^ а б c Колоджей П.А., Молодой Р.А. (сентябрь 1991 г.). «Мутации в трех крупнейших субъединицах дрожжевой РНК-полимеразы II, которые влияют на сборку ферментов». Молекулярная и клеточная биология. 11 (9): 4669–78. Дои:10.1128 / mcb.11.9.4669. ЧВК 361357. PMID 1715023.
- ^ Каплан CD, Цзинь Х., Чжан И.Л., Белянин А (12 апреля 2012 г.). «Рассечение функции триггерной петли Pol II и зависимый от активности Pol II контроль выбора стартового сайта in vivo». PLOS Genetics. 8 (4): e1002627. Дои:10.1371 / journal.pgen.1002627. ЧВК 3325174. PMID 22511879.
- ^ а б Abbondanzieri EA, Greenleaf WJ, Shaevitz JW, Landick R, Block SM (ноябрь 2005 г.). «Прямое наблюдение шагового перехода пар оснований с помощью РНК-полимеразы». Природа. 438 (7067): 460–5. Bibcode:2005Натура.438..460А. Дои:10.1038 / природа04268. ЧВК 1356566. PMID 16284617.
- ^ а б Ходжес, Кортни; Бинту, Лакрамиоара; Лубковская, Люцина; Кашлев Михаил; Бустаманте, Карлос (31.07.2009). «Нуклеосомные колебания определяют динамику транскрипции РНК-полимеразы II». Наука. 325 (5940): 626–628. Bibcode:2009Sci ... 325..626H. Дои:10.1126 / science.1172926. ISSN 1095-9203. ЧВК 2775800. PMID 19644123.
- ^ Черчман, Л. Стирлинг; Вайсман, Джонатан С. (20 января 2011 г.). «Зарождающееся секвенирование транскриптов визуализирует транскрипцию при разрешении нуклеотидов». Природа. 469 (7330): 368–373. Bibcode:2011Натура.469..368C. Дои:10.1038 / природа09652. ISSN 1476-4687. ЧВК 3880149. PMID 21248844.
- ^ Galburt, Eric A .; Гриль, Стефан В .; Видманн, Анна; Лубковская, Люцина; Чой, Джейсон; Ногалес, Ева; Кашлев Михаил; Бустаманте, Карлос (2007-04-12). «Обратное отслеживание определяет силовую чувствительность RNAP II в зависимости от факторов». Природа. 446 (7137): 820–823. Bibcode:2007Натура.446..820Г. Дои:10.1038 / природа05701. ISSN 1476-4687. PMID 17361130. S2CID 4310108.
- ^ Бинту, Лакрамиоара; Копачинская, Марта; Ходжес, Кортни; Лубковская, Люцина; Кашлев Михаил; Бустаманте, Карлос (13 ноября 2011 г.). «Скорость удлинения РНК-полимеразы определяет судьбу транскрибируемых нуклеосом». Структурная и молекулярная биология природы. 18 (12): 1394–1399. Дои:10.1038 / nsmb.2164. ISSN 1545-9985. ЧВК 3279329. PMID 22081017.
- ^ Каплан CD, Ларссон К.М., Корнберг Р.Д. (июнь 2008 г.). «Триггерная петля РНК-полимеразы II участвует в выборе субстрата и напрямую нацелена на альфа-аманитин». Молекулярная клетка. 30 (5): 547–56. Дои:10.1016 / j.molcel.2008.04.023. ЧВК 2475549. PMID 18538653.
- ^ Gong, Xue Q .; Недиалков, Юрий А .; Бертон, Захари Ф. (2004-06-25). «-Аманитин блокирует транслокацию человеческой РНК-полимеразы II». Журнал биологической химии. 279 (26): 27422–27427. Дои:10.1074 / jbc.M402163200. ISSN 0021-9258. PMID 15096519.
- ^ Бриггс, Скотт Д.; Брик, Мэри; Strahl, Brian D .; Cheung, Wang L .; Дэви, Джудит К .; Dent, Sharon Y. R .; Уинстон, Фред; Аллис, К. Дэвид (2001-12-15). «Метилирование гистона H3 по лизину 4 опосредуется Set1 и требуется для роста клеток и подавления рДНК у Saccharomyces cerevisiae». Гены и развитие. 15 (24): 3286–3295. Дои:10.1101 / gad.940201. ISSN 0890-9369. ЧВК 312847. PMID 11751634.
- ^ Ли, Бинг; Хау, Лиэнн; Андерсон, Скотт; Йейтс, Джон Р .; Уоркман, Джерри Л. (14 марта 2003 г.). "Гистон-метилтрансфераза Set2 функционирует через фосфорилированный карбоксиконцевой домен РНК-полимеразы II". Журнал биологической химии. 278 (11): 8897–8903. Дои:10.1074 / jbc.M212134200. ISSN 0021-9258. PMID 12511561.
- ^ Мейнхарт А., Крамер П. (июль 2004 г.). «Распознавание карбоксиконцевого домена РНК-полимеразы II факторами процессинга 3'-РНК». Природа. 430 (6996): 223–6. Bibcode:2004Натура 430..223М. Дои:10.1038 / природа02679. PMID 15241417. S2CID 4418258.
- ^ а б c Эглофф, Сильвен; Мерфи, Шона (2008). «Взлом кода CTD РНК-полимеразы II». Тенденции в генетике. 24 (6): 280–288. Дои:10.1016 / j.tig.2008.03.008. PMID 18457900.
- ^ Вэй Л., Левин А.С., Лан Л. (2016). «Транскрипционно-связанная гомологичная рекомбинация после окислительного повреждения». Ремонт ДНК (Amst.). 44: 76–80. Дои:10.1016 / j.dnarep.2016.05.009. PMID 27233112.
внешняя ссылка
(Wayback Machine копия)
- РНК + Полимераза + II в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)