Bovidae - Bovidae
| Bovidae | |
|---|---|
 | |
| Пример Bovidae (по часовой стрелке сверху слева) - аддакс, домашний скот, газель, импала, антилоп гну, и муфлон | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Млекопитающие |
| Заказ: | Парнокопытные |
| Инфразаказ: | Pecora |
| Семья: | Bovidae серый, 1821 |
| Тип род | |
| Bos | |
| Подсемейства | |
Aepycerotinae (1 род) | |
В Bovidae составляют биологическая семья из парнокопытный, жвачный позвоночные это включает зубр, Африканский буйвол, буйвол, антилопы, овца, козы, овцебык, и внутренние крупный рогатый скот. Член этой семьи называется бычий. С 143 сохранившийся видов и 300 известных вымерших видов, семейство Bovidae состоит из восьми основных подсемейства помимо спорных Пелейны и Pantholopinae. Семья возникла 20 миллионов лет назад, в начале Миоцен.
Крупные рогатые животные сильно различаются по размеру и шерсть окраска. За исключением некоторых одомашненные формы, у всех самцов есть два или более рога, и у многих видов самки тоже обладают рогами. Размер и форма рогов сильно различаются, но основная структура всегда представляет собой одну или несколько пар простых костных выступов без ветвей, часто имеющих спиральную, скрученную или рифленую форму, каждый из которых покрыт постоянной оболочкой из кератин. У большинства быков от 30 до 32 зубов.
Большинство быков дневной. Социальная активность и кормление обычно достигают пика на рассвете и в сумерках. Быковиды обычно отдыхают до рассвета, в полдень и после наступления темноты. У них разные методы социальной организации и социальное поведение, которые подразделяются на уединенное и стадное поведение. Бовиды используют разные формы вокального, обонятельного и осязаемого общения. Большинство видов в течение дня поочередно питаются и пережевывают. В то время как мелкие коровы кормятся в плотной и закрытой среде обитания, более крупные виды питаются растительностью с высоким содержанием клетчатки на открытых лугах. Большинство быков полигинный. Взрослые быки спариваются не реже одного раза в год, а более мелкие особи могут даже спариваться дважды. У некоторых видов новорожденные коровы остаются скрытыми от недели до двух месяцев, и их матери регулярно кормят грудью; у других видов новорожденные - это последователи, которые сопровождают своих маток, а не скрываются.
Наибольшее разнообразие полорогих встречается в Африка. Максимальная концентрация видов в саванны Восточной Африки. Другие виды быков также встречаются в Европе, Азии и Северной Америке. Bovidae включает три из пяти одомашненный млекопитающие, использование которых распространилось за пределы их первоначального ареала, а именно крупный рогатый скот, овцы и козы. Молочные продукты Такие как молоко, масло сливочное, и сыр в основном производятся из домашнего скота. Bovids также обеспечивают натуральная кожа, мясо, и шерсть.
Этимология
Название «Bovidae» дал британский зоолог. Джон Эдвард Грей в 1821 г.[1] Слово «Bovidae» - это комбинация приставки bov- (происходящих от латинский БОС, "бык", через Поздняя латынь бовинус) и суффикс -idae.[2]
Таксономия
В семья Bovidae помещается в порядок Парнокопытные (что включает в себя копытные ). Он включает 143 современных вида, что составляет почти 55% всех видов. копытные и 300 известных вымерших видов.[3]
Молекулярные исследования подтвердили монофилия в семействе Bovidae (группа организмов состоит из предков и всех их потомков).[4][5] Количество подсемейства in Bovidae оспаривается, предлагается целых десять и всего два подсемейства.[5] Однако молекулярная, морфологический и ископаемые свидетельства указывают на существование восьми отдельных подсемейств: Aepycerotinae (состоящий только из импалы), Alcelaphinae (бонтебок, гну, антилопа гну и родственники), Antilopinae (несколько антилоп, газелей и родственников), Bovinae (крупный рогатый скот, буйволы, бизоны и другие антилопы), Caprinae (козы, овцы, горные козлы, серы и родственники), Цефалофины (дукерс), Hippotraginae (аддакс, орикс и родственники) и Reduncinae (антилопы ридбак и коб). Кроме того, известны три вымерших подсемейства: Hypsodontinae (средний миоцен), Oiocerinae (Туролийский ) и подсемейство Tethytraginae, которое содержит Тетитрагус (средний миоцен).[6][7]
В 1992 году Алан У. Джентри из Музей естественной истории, Лондон разделил восемь основных подсемейств Bovidae на две основные клады на основе их эволюционной истории: Boodontia, которая включала только Bovinae, и Aegodontia, которая состояла из остальных подсемейств. Будонты несколько примитивный зубы, напоминающие бычьи, тогда как у эгодонтов больше передовой зубы, как у козла.[8]
Существуют разногласия по поводу признания Peleinae и Patholopinae, составляющих роды Пелеа и Пантолопс соответственно, как подсемейства. В 2000 году американский биолог Джордж Шаллер и палеонтолог Элизабет Врба предложил включить Пелеа в Редунцинах,[9] хотя серый рабок, единственный вид Пелеа, сильно отличается от кобов и редунцинов по морфологии.[10] Пантолопс, ранее классифицированный как Antilopinae, позже был помещен в собственное подсемейство Pantholopinae. Однако молекулярный и морфологический анализ поддерживает включение Пантолопс в Caprinae.[11]
Ниже представлена кладограмма, основанная на Gatesy et al. (1997) и Gentry et al. (1997)
| Bovidae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||






Эволюция
Ранний миоцен и раньше

В раннем миоцене бычьи начали отходить от шейки матки (олень) и жирафы. Самые ранние полевые быки, присутствие которых в Африке и Евразии во второй половине раннего миоцена (20 Mya ), были мелкими животными, чем-то похожими на современных газели, и, вероятно, жил в лесной массив среды.[12] Эотрагус, самый ранний известный рогатый скот, весил 18 кг (40 фунтов) и был почти такого же размера, как и Газель Томпсона.[13] В начале своей эволюционной истории полорогие разделились на две основные клады: Boodontia (евразийского происхождения) и Aegodontia (африканского происхождения). Так рано расколоть между Boodontia и Aegodontia было приписано континентальному водоразделу между этими массивами суши. Когда эти континенты позже воссоединились, этот барьер был снят, и каждая группа расширилась на территорию другой.[14] Племена Бовини и Tragelaphini разошлись в раннем миоцене.[15] Известно, что Bovids достигли Америка в плейстоцене, пересекая Берингов мост.[13]
Современные роды Alcelaphinae появились в Плиоцен. Вымерший род Alcelaphine Paramularius, который был такого же размера, как и хищник, как полагают, возник в плиоцене, но вымер в середине Плейстоцен.[5] Несколько родов Hippotraginae известны с плиоцена и плейстоцена. Это подсемейство, по-видимому, отделилось от Alcelaphinae во второй половине раннего миоцена.[15] Считается, что у Bovinae расходился от остальных Bovidae в начале Миоцен.[16] Boselaphini вымерли в Африке в начале плиоцена; их последние окаменелости были раскопаны в Langebaanweg (Южная Африка) и Лотагам (Кения).[17]
Средний миоцен
Средний миоцен ознаменовал распространение быков в Китай и на Индийский субконтинент.[13] По словам Врба, радиация подсемейства Alcelaphinae возникли во второй половине среднего миоцена.[5] Племена Caprinae, вероятно, разошлись в раннем среднем миоцене. Caprini появились в среднем миоцене и, кажется, были заменены другими бычковыми и цервидными в Евразии.[18] Самые ранние окаменелости антилопин относятся к среднему миоцену, хотя исследования показывают, что это подсемейство относится к раннему миоцену. Видообразование произошло в племя Антилопини в среднем или верхнем миоцене, в основном в Евразии. Племя Neotragini По всей видимости, он появился в Африке к концу миоцена и получил широкое распространение к плиоцену.[15]
Поздний миоцен
К концу миоцена, примерно к 10 млн лет назад, половые животные быстро разнообразный, что привело к созданию 70 новых родов.[13] Эта радиация в позднем миоцене отчасти была вызвана тем, что многие быки приспособились к более открытым местам обитания на лугах.[12] Впервые Aepycerotinae появились в позднем миоцене, и существенной разницы в размерах примитивной и современной импалы замечено не было.[19] Окаменелости яйцеклеток, племени Caprinae, в Африке восходят к позднему миоцену.[15] Самые ранние окаменелости Hippotragine относятся к позднему миоцену и были раскопаны в таких местах, как Лотагам и Awash Valley.[15] Первые африканские окаменелости Reduncinae датируются 6-7 млн лет назад.[20] Расхождение Reduncinae и Peleinae, вероятно, произошло в середине миоцена.[5]
Характеристики

У всех крупного рогатого скота схожая основная форма - тупой конец морды, одна или несколько пар рогов (обычно присутствующих у самцов) сразу после овальных или заостренных ушей, отчетливая шея и конечности, а также хвост, различающийся по длине и густоте. виды.[21] Большинство быков выставляются половой диморфизм, причем самцы обычно крупнее и тяжелее самок. Половой диморфизм более выражен у средних и крупных рогатых животных. У всех крупного рогатого скота по четыре пальца на каждой ступне - они ходят на двух центральных ( копыта ), а два внешних ( dewclaws ) намного меньше по размеру и редко касаются земли.[3]
Крупные рогатые животные сильно различаются по размеру: гаур может весить более 1500 кг (3300 фунтов) и достигать высоты 2,2 м (87 дюймов) в плече.[22] В королевская антилопа, напротив, его высота составляет всего 25 см (9,8 дюйма), а вес - не более 3 кг (6,6 фунта).[23] В клипспрингер, еще одна небольшая антилопа, имеет высоту 45–60 см (18–24 дюйма) в плече и весит всего 10–20 кг (22–44 фунта).[24]
Различия возникают в шерсть окраска в диапазоне от бледно-белого (как в Арабский орикс )[25] в черный (как в черный гну ).[26] Однако только промежуточные оттенки, такие как коричневый и красновато-коричневый (как в Reedbuck ), обычно наблюдаются.[27] У некоторых видов самки и молодые особи имеют светлую шерсть, а у самцов с возрастом темнеют. Как в антилоп гну шерсть может быть отмечена заметными или слабыми полосами. У некоторых видов, таких как аддакс, окрас шерсти может меняться в зависимости от сезона.[28] Ароматические железы и сальные железы часто присутствуют.[21]

Некоторые виды, такие как гемсбок, соболиная антилопа, и Газель Гранта, находятся замаскированный с сильно разрушительный отметины на лице, скрывающие очень узнаваемый глаз.[30] Многие виды, такие как газели, могут выглядеть плоскими и, следовательно, сливаться с фоном, с помощью затенение.[31] Очертания многих крупных рогатых животных разделены яркой разрушительной окраской, а резко контрастирующие узоры помогают задержать распознавание хищниками.[32] Однако все Hippotraginae (включая гемсбока) имеют бледные тела и лица с заметными отметинами. Зоолог Тим Каро описывает это как трудно объяснимое, но, учитывая, что эти виды ведут дневной образ жизни, он предполагает, что отметины могут функционировать при общении. Сильно контрастная окраска ног обычна только у Bovidae, где, например, Bos, Овис, бонтебок а у гемсбока белые чулки. Опять же, общение - это вероятная функция.[29]
За исключением некоторых одомашненных форм, у всех самцов быков есть рога, и у многих видов самки тоже. Размер и форма рогов сильно различаются, но основная структура представляет собой пару простых костных выступов без ветвей, часто имеющих спиральную, витую или рифленую форму, каждый из которых покрыт постоянной оболочкой из кератина. Хотя рога встречаются в одной паре почти у всех видов рогатых животных, есть исключения, такие как четырехрогая антилопа[33] и Овца иакова.[34][35] Уникальное строение рогов - единственная недвусмысленная морфологическая особенность коров, которая отличает их от других. Pecorans.[36][37] Между морфологией рогов и боевым поведением особи существует высокая корреляция. Например, длинные рога предназначены для борьбы и фехтования, а изогнутые - для тарана.[38] Самцы с направленными внутрь рогами моногамный и одинокие, тогда как те, у кого рога направлены наружу, как правило, полигинный. Эти результаты не зависели от размера тела.[39]
Развитие мужского рога связано с половой отбор,[40][41] Рога - это маленькие шипы у моногамных дукеров и других мелких антилоп, тогда как у полигинных они большие и замысловато сформированные (например, в спиральной структуре, как у гигантская канна ). Таким образом, в некоторой степени рога отображают степень конкуренция среди самцов вида.[27] Однако наличие рогов у самок, вероятно, связано с естественный отбор.[40][42] Рога у самок обычно меньше, чем у самцов, а иногда и иной формы. Считается, что рога самок крупного рогатого скота эволюционировали для защиты от хищников или для выражения территориальности, поскольку самки, не имеющие территориальной принадлежности, могут использовать крипсида для защиты от хищников часто не имеют рогов.[42] Самки имеют рога только у половины родов полукровок, причем самки этих родов тяжелее остальных. Самки используют рога в основном для колющих ударов.[43]

Анатомия
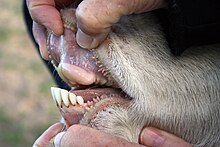
У крупного рогатого скота третий и четвертый метаподиалы объединены в берцовая кость. В локтевая кость и малоберцовая кость уменьшаются и срастаются с лучевой и большеберцовой костью соответственно. Длинный лопатки присутствуют, а ключицы отсутствуют. Существование жвачные животные, желудок состоит из четырех камер: рубец (80%), омазум, то сеточка, а сычуг. В инфузории и бактерии рубца брожение комплекс целлюлоза в более простой жирные кислоты, которые затем всасываются через стенку рубца. У быков долгая тонкий кишечник; длина тонкой кишки в крупный рогатый скот составляет 29–49 м (95–161 футов). Температура тела колеблется в течение дня; например, у коз температура может незначительно изменяться от почти 37 ° C (99 ° F) ранним утром до 40 ° C (104 ° F) днем. У крупного рогатого скота температура регулируется потоотделением, тогда как козы используйте одышку для того же. Право легкое, состоящий из четырех-пяти доли, примерно в 1,5 раза больше, чем левая, имеющая три доли.[3][21]
Зубы
У большинства быков от 30 до 32 зубов.[27] Пока верхний резцы отсутствуют, верхние клыки либо уменьшены, либо отсутствуют. Вместо верхних резцов у быков толстый и прочный слой ткани, называемый зубная прокладка, которая обеспечивает поверхность для захвата травы и листвы. Они есть гипсодонт и селенодонт, поскольку коренные зубы и премоляры низкие-увенчанный и в форме полумесяца куспиды. Нижние резцы и клыки выступают вперед. За резцами следует длинная беззубая щель, известная как диастема.[44] Генерал зубная формула для коров это 0.0.2-3.33.1.3.3. Большинство членов семьи травоядный, но большинство дукеров всеядный. Как и у других жвачных животных, у крупного рогатого скота есть желудки с четырьмя камерами, которые позволяют им переваривать растительный материал, такой как трава, который не может использоваться многими другими животными. Жвачные (и некоторые другие любят кенгуру, кролики, и термиты ) умеют использовать микроорганизмы живя в их кишках, чтобы расщепить целлюлозу ферментация.[3]
Экология и поведение


У коров есть различные методы социальной организации и социального поведения, которые подразделяются на одиночное и стадное поведение. Кроме того, каждый из этих типов можно разделить на территориальное и нетерриториальное поведение.[27] Мелкие рогатые, такие как клипспрингер, Ориби, и Steenbok обычно одинокие и территориальные. Они занимают небольшие территории, на которые другим представителям вида не разрешается входить. Эти антилопы образуют моногамный пары. Многие виды, такие как дик-дик использовать феромон выделения из предглазничные железы а иногда и навоз, чтобы пометить свою территорию.[45] Потомство расселяется в подростковом возрасте, и самцы должны осваивать территории до спаривания.[3] В кустарник это единственный крупный рогатый скот, который одновременно обособлен и не территориален. Эта антилопа почти не проявляет агрессии и имеет тенденцию изолироваться или образовывать рыхлые стада, хотя в благоприятной среде обитания можно встретить несколько кустарников довольно близко друг к другу.[46]
За исключением цефалофинов (дуикеров), трагелафинов (спиралевидных антилоп) и неотрагин, большинство африканских бычков стайно и территориально. Самцы вынуждены рассредоточиться по достижении половой зрелости и должны образовывать свои собственные территории, в то время как самки не обязаны этого делать. Самцы, не владеющие территориями, образуют холостяцкие стада. Мужчины соревнуются за доминирование, и драки, как правило, проходят более жестко в ограниченном количестве. сезоны гона. За исключением мигрирующих самцов, самцы обычно занимают одну и ту же территорию на протяжении всей своей жизни.[27] В козлах некоторые особи самцов, известные как «самцы-спутники», могут быть допущены на территорию других самцов и должны ждать, пока владелец состарится, чтобы они могли завладеть его территорией.[47] Лек вязка, где самцы собираются вместе и соревнуются друг с другом, как известно, существует среди топис, Кобс, и Лехве.[48] Трагелафины, крупный рогатый скот, овцы и козы стадны, а не территориальны. У этих видов самцы должны получить абсолютное доминирование над всеми остальными самцами, и драки не ограничиваются территориями. Поэтому самцы тратят годы на рост тела.[27]
Мероприятия

Большинство быков ведут дневной образ жизни, хотя некоторые из них, такие как буйвол, бушбак, тростник и грызбок, являются исключением. Социальная активность и кормление обычно достигают пика на рассвете и в сумерках. Полководцы обычно отдыхают перед рассветом, в полдень и после наступления темноты. Уход обычно заключается в облизывании языком. Антилопы редко катаются в грязи или пыли. Антилопы гну и буйволы обычно валяются в грязи, тогда как хищники и топи протирают грязью головы и рога, а затем размазывают ею свои тела. Бовиды используют разные формы вокального, обонятельного и осязаемого общения. Они включают в себя различные позы шеи, головы, рогов, волос, ног и ушей, чтобы передать сексуальное возбуждение, эмоциональное состояние или тревогу. Одним из таких выражений является ответ флемена. Боровы обычно стоят неподвижно, с высоко поднятой головой и пристальным взглядом, когда чувствуют опасность. Некоторые, такие как импала, куду и канна, могут даже прыгать на высоту нескольких футов.[27] Бычьи животные могут рычать или кряхтеть, чтобы предупредить других и предупредить хищников.[3] Бычьи, такие как газели стот или пронк в ответ на хищников, совершая высокие прыжки на окоченевших ногах, показывая честно и то, что хищник был замечен, и что стоящая особь сильна и не стоит того, чтобы ее преследовать.[49]

В брачный период самцы издают рев, чтобы самки узнали о своем присутствии. Овцебыки ревет во время схваток самцов с самцами, а самцы сайгаков проталкивают воздух через нос, издавая рев, чтобы отпугнуть соперничающих самцов и привлечь самок. Матери также используют голосовое общение, чтобы определить местонахождение своих телят, если они разлучены. Во время борьбы за доминирование мужчины склонны отображать в вертикальном положении с ровной мордой.[50][51]
Техника боя у коров различается, а также зависит от их телосложения. В то время как самые хищные дерутся на коленях, другие обычно дерутся на четвереньках. Газели разных размеров используют разные приемы боя. Газели обычно боксируют, а в серьезных боях могут столкнуться и фехтовать, состоящие из жестких ударов с близкого расстояния. Самцы козерога, козы и овцы стоят вертикально и сталкиваются друг с другом вниз. Гну используют мощные удары головой в агрессивных столкновениях. Если рога запутались, противники двигаются по кругу, чтобы разблокировать их. Овцебыки будут врезаться друг в друга на большой скорости. Как правило, только два быка одинакового телосложения и уровня защиты участвуют в бою, который предназначен для определения лучшего из них. Люди, которые явно уступают другим, скорее убегут, чем сражаются; например, неполовозрелые самцы не борются со взрослыми быками. Как правило, быки направляют свои атаки на голову противника, а не на его тело. S-образные рожки, такие как импала, имеют различные секции, которые помогают таранить, удерживать и колоть. Серьезные драки, приводящие к травмам, случаются редко.[27][50][52]
Рацион питания

Большинство рогатых животных в течение дня попеременно кормятся и пережевывают. В то время как те, кто питается концентрированным кормом и переваривает их за короткие промежутки времени, кормушки для грубых кормов используют более длительные интервалы. Лишь мелкие виды, такие как дукер, пасутся в течение нескольких часов днем или ночью.[27] Привычки питания зависят от размера тела; в то время как мелкие коровы кормятся в плотной и закрытой среде обитания, более крупные виды питаются растительностью с высоким содержанием клетчатки на открытых лугах. Подсемейства демонстрируют разные стратегии питания. В то время как виды Bovinae активно пасутся на свежей траве и разбросанных кормах, виды Cephalophinae (за исключением Сильвикапра ) в первую очередь потребляют фрукты.[3] Виды Reduncinae и Hippotraginae зависят от нестабильных источников пищи, но последние специально приспособлены к засушливым районам. Представители Caprinae, будучи гибкими кормушками, кормятся даже на участках с низкой продуктивностью. Племена Alcelaphini, Hippotragini и Reduncini имеют высокую долю однодольные в их диетах. Напротив, Tragelaphini и Neotragini (за исключением Оребия ) активно питаются двудольные.[53] Нет заметной связи между размером тела и потреблением однодольных.[54]
Сексуальность и размножение

Большинство полигинных животных. У некоторых видов особи моногамны, что приводит к минимальной агрессии самцов по отношению к самцам и сокращению отбора самцов в отношении больших размеров тела. Таким образом, половой диморфизм практически отсутствует. Самки могут быть немного крупнее самцов, возможно, из-за конкуренции между самками за владение территориями. Так обстоит дело с дукерами и другими мелкими рогатками.[55][56] Время, необходимое для достижения половая зрелость в зависимости от пола широко варьируется среди рогатых животных. Половая зрелость может даже предшествовать спариванию или следовать за ним. Например, самцы импала, хотя и достигают половой зрелости к году, могут спариваться только после четырехлетнего возраста.[57] Напротив варварская овца самки могут производить потомство даже до достижения половой зрелости.[58] Задержка полового созревания самцов более заметна у половодиморфных видов, особенно у редунцинов, вероятно, из-за конкуренции между самцами.[3] Например, синий гну самки становятся способными к воспроизводству в течение года или двух после рождения, тогда как самцы становятся зрелыми только в возрасте четырех лет.[26]
Все быки спариваются не реже одного раза в год, а более мелкие виды могут даже спариваться дважды. Сезон спаривания у большинства рогатых обычно приходится на дождливые месяцы. Таким образом, пик размножения может достигаться дважды в экваториальных регионах. Овцы и козы демонстрируют замечательную сезонность воспроизводства, при определении которой годовой цикл суточного фотопериод играет ключевую роль. Другие факторы, которые оказывают значительное влияние на этот цикл, включают температуру окружающей среды, пищевой статус, социальные взаимодействия, дату родов и период лактации. Исследование этого явления пришло к выводу, что козы и овцы заводчики короткого дня. Спаривание у большинства пород овец начинается летом или ранней осенью.[59] На спаривание у овец также влияет мелатонин, который приближает начало сезона размножения;[60] и тироксин, что завершает сезон размножения.[61] Течка длится не более суток у крупного рогатого скота, за исключением крупного рогатого скота и трагелафин. За исключением хартебеста и топи, все коровы могут определять течку у самок, анализируя мочу с помощью вомероназальный орган.[27] Как только самец уверен, что самка находится в течке, он начинает ухаживать; эти проявления сильно различаются: от замысловатых маршей среди стайных видов до страстного облизывания женских гениталий среди одиночных видов. Самки, изначально невосприимчивые, в конечном итоге спариваются с самцом, который добился господства над другими. Восприимчивость выражается разрешением самца садиться на него и откладывать хвост самкой. Копуляция обычно занимает несколько секунд.[27][55]
Период беременности варьируется среди рогатых животных - в то время как беременность дукеров колеблется от 120 до 150 дней, беременность африканского буйвола колеблется от 300 до 330 дней. Обычно рождается одинокий потомок (близнецы реже), и он способен самостоятельно стоять и бегать в течение часа после рождения. У моногамных видов самцы помогают защитить своих детенышей, но это не относится к полигамным видам. Большинство новорожденных телят остаются скрытыми от недели до двух месяцев, и их матери регулярно кормят грудью. У некоторых видов быков новорожденные начинают следить за своей матерью сразу или в течение нескольких дней, как у импалы.[57] У разных быков разные стратегии защиты молоди. Например, в то время как матери гну защищают исключительно своих детенышей, буйволы демонстрируют коллективную защиту. Отлучение от груди может происходить уже через два месяца (как у королевской антилопы) или в течение года (как у королевской антилопы). овцебык ).[55][56]
Срок жизни
Большинство диких быков живут от 10 до 15 лет. Более крупные виды обычно живут дольше;[3] например, Американский бизон могут жить до 25 лет, а гаур до 30 лет.Средняя продолжительность жизни одомашненных особей составляет почти десять лет. Например, средняя продолжительность жизни домашних коз составляет 12 лет. Обычно у самцов, в основном у полигинных видов, продолжительность жизни короче, чем у самок. Это может быть связано с несколькими причинами: раннее расселение молодых самцов, агрессивные схватки самцов с самцами, уязвимость перед хищниками (особенно когда самцы менее подвижны, как в куду) и недоедание (будучи большими по размеру, самец имеет высокую питательную ценность. требования, которые могут не быть выполнены).[62][63] Ричард Деспар Эстес предположили, что самки имитируют вторичные половые признаки самцов, такие как рога, чтобы защитить свое потомство от доминирующих самцов. Эта функция, по-видимому, была тщательно выбрана для предотвращения мужской смертности и несбалансированности соотношение полов из-за нападений агрессивных самцов и вынужденного расселения молодых самцов во время юность.[64]
Распределение

Большинство разнообразных видов быков встречается в Африке. Максимальная концентрация находится в саванны Восточной Африки. В зависимости от привычки питания у нескольких видов излученный на больших участках суши, и, следовательно, наблюдается несколько вариаций морфологии зубов и конечностей. Дукеры населяют экваториальные тропические леса, ситатунга, и lechwe встречаются у болот, канна населяет луга, газель и орикс встречаются в пустынях, бонго и аноа живут в густых лесах, а горные козлы и такин живут на больших высотах.[27] Несколько видов быков также встречаются в Европе, Азии и Северной Америке. Овцы и козы водятся в основном в Евразии, хотя берберийские овцы и горные козлы являются частью африканской фауны. Овцебык приурочен к арктическая тундра. Некоторые виды быков были одомашнены людьми. Приручение коз и овец началось 10 тысяч лет назад, а крупный рогатый скот - около 7,5 тысяч лет назад.[3][55]
Взаимодействие с людьми
Домашние животные

В приручение bovids способствовал смещению зависимости людей от охоты и собирательства к сельскому хозяйству. К Bovidae относятся три из пяти крупных домашних травоядных, использование которых распространилось за пределы их первоначального ареала, а именно крупный рогатый скот, овцы и козы; все они из Евразии и теперь встречаются по всему миру. Два других вида - это лошадь и свинья. Другими крупными рогатками, которые были одомашнены, но которые остались в пределах ареалов своих диких предков, являются буйвол (от Индийский буйвол ), домашний як (от дикий як ), зебу (от Индийские зубры ), гаял (от гаур ) и Балийский скот (от Бантенг ).[55]Некоторые антилопы были одомашнены, в том числе Орикс, аддакс, канна и вымершие Bubal Hartebeest. В Древний Египет Ориксы, аддаксы и бубальные хищники изображены на резных стенах.
Самые ранние свидетельства одомашнивания крупного рогатого скота относятся к 8000 г. до н.э., что позволяет предположить, что этот процесс начался в Кипр и Евфрат бассейн.[65]
Животные продукты

Молочные продукты Такие как молоко, масло сливочное, топленое масло, йогурт, пахта и сыр в основном производятся из домашнего скота, хотя молоко овец, коз, яков и буйволов также используется в некоторых частях мира для производства деликатесов. Например, из буйволиного молока делают моцарелла в Италии и Гулаб Джамун десерт в Индии,[66] в то время как овечье молоко используется для получения синего Рокфор сыр во Франции.[67]Говядина отличный источник цинк, селен, фосфор, утюг, и Витамины группы B.[68] В мясе бизона меньше жира и холестерина, чем в говядине, но в нем больше белка.[69]
Бовид натуральная кожа прочный и долговечный, с дополнительным преимуществом: из него можно изготавливать кожу различной толщины - от мягкой одежды для одежды до жесткой кожи для обуви. Кожа козла и крупного рогатого скота имеет широкое применение, овчина подходит только для одежды.[70] Шерсть из Меринос кабачки самый лучший и самый ценный. мериносовой шерсти 3–5 дюймов (7,6–12,7 см) в длину и очень мягкие. Грубая шерсть, будучи прочной и устойчивой к пиллинг, используются для изготовления жесткой одежды[71] и ковры.

Костная мука это важный удобрение богат кальцием, фосфором и азот, эффективно удаляет кислотность почвы.[72] Рога крупного рогатого скота использовались как сосуды для питья с древности.[73]
В человеческой культуре
Bovidae фигурируют в рассказах, по крайней мере, со времен Басни Эзопа из Древняя Греция около 600 г. до н. э. Басни Эзопа включают Ворона и овца, Лягушка и Бык, и Волк и ягненок.[74] Мифологическое существо Химера, изображавшееся в виде льва, с головой козла, выступающей из спины, и хвостом, который мог заканчиваться головой змеи, было одним из потомков Тифон и Ехидна и брат таких монстров, как Цербер и Лернейская гидра.[75] Овца, синоним козел в китайской мифологии это восьмое животное из китайский гороскоп, и символ сыновней почтительности.[76]
Рекомендации
- ^ Грабб, П. (2005). «Семья Bovidae». В Уилсон, Д.; Ридер, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Издательство Университета Джона Хопкинса. С. 637–722. ISBN 978-0-8018-8221-0. OCLC 62265494.
- ^ "Bovidae". Онлайн-словарь Merriam-Webster. Получено 7 октября 2014.
- ^ а б c d е ж грамм час я j Gomez, W .; Паттерсон, Т. А .; Swinton, J .; Берини, Дж. «Bovidae: антилопы, крупный рогатый скот, газели, козы, овцы и родственники». Сеть разнообразия животных. Музей зоологии Мичиганского университета. Получено 7 октября 2014.
- ^ Gatesy, J .; Amato, G .; Vrba, E .; Шаллер, Г. (1997). «Кладистический анализ митохондриальной рибосомной ДНК от Bovidae». Молекулярная филогенетика и эволюция. 7 (3): 303–19. Дои:10.1006 / mpev.1997.0402. PMID 9187090.
- ^ а б c d е Fernández, M. H .; Врба, Э. С. (2005). «Полная оценка филогенетических отношений у Ruminantia: датированное наддерево на уровне видов современных жвачных». Биологические обзоры. 80 (2): 269–302. Дои:10.1017 / S1464793104006670. PMID 15921052. S2CID 29939520.
- ^ Харрисон, Т. (2011). Палеонтология и геология эволюции человека Лаэтоли в контексте. Дордрехт: Спрингер. С. 363–465. ISBN 978-9048-199-624.
- ^ Demiguel, D .; Sánchez, I.M .; Альба, Д. М .; Галиндо, Дж .; Роблес, Дж. М .; Моя-Сола, С. (2012). "Первые свидетельства существования Azanza and Morales, 1994 (Ruminantia, Bovidae), в миоцене бассейна Валлес-Пенедес (Испания)". Журнал палеонтологии позвоночных. 32 (6): 1457–62. Дои:10.1080/02724634.2012.696082. S2CID 86664298.
- ^ Харрисон, Т. (1997). Неогеновая палеонтология долины Манонга, Танзания: окно в эволюционную историю Восточной Африки. Нью-Йорк: Пленум Пресс. п. 113. ISBN 978-0-306-45471-4.
- ^ Vrba, E. S .; Шаллер, Г. (2000). Антилопы, олени и родственники: летопись окаменелостей, поведенческая экология, систематика и сохранение. Нью-Хейвен: издательство Йельского университета. ISBN 978-0300-081-428.
- ^ Грабб, П. (2005). "Отряд парнокопытных". В Уилсон, Д.; Ридер, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Издательство Университета Джона Хопкинса. п. 719. ISBN 978-0-8018-8221-0. OCLC 62265494.
- ^ Грабб, П. (2005). "Отряд парнокопытных". В Уилсон, Д.; Ридер, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Издательство Университета Джона Хопкинса. п. 699. ISBN 978-0-8018-8221-0. OCLC 62265494.
- ^ а б Savage, R.J.G .; Лонг, М.Р. (1986). Эволюция млекопитающих: иллюстрированное руководство. Нью-Йорк: факты в файле. стр.232–5. ISBN 978-0-8160-1194-0.
- ^ а б c d Prothero, D. R .; Шох, Р. М. (2002). Рога, клыки и ласты: эволюция копытных млекопитающих. Балтимор: Издательство Университета Джона Хопкинса. С. 87–90. ISBN 978-0-8018-7135-1.
- ^ Hassanin, D .; Дузери, Э.Дж. (1999). "Племенное излучение семейства Bovidae (Artiodactyla) и эволюция митохондриального цитохрома б ген" (PDF). Молекулярная филогенетика и эволюция. 13 (2): 227–43. Дои:10.1006 / mpev.1999.0619. PMID 10603253. Архивировано из оригинал (PDF) на 2011-07-20.
- ^ а б c d е Gilbert, W.H .; Асфау, Б. (2008). Homo Erectus: свидетельства плейстоцена из Среднего Аваша, Эфиопия. Беркли: Калифорнийский университет Press. С. 45–84. ISBN 978-0-520-25120-5.
- ^ Биби, Ф. (август 2013 г.). «Многокалиброванная митохондриальная филогения современных Bovidae (Artiodactyla, Ruminantia) и важность летописи окаменелостей для систематики». BMC Эволюционная биология. 13: 166. Дои:10.1186/1471-2148-13-16. ЧВК 3607928. PMID 23339550.CS1 maint: использует параметр авторов (связь)
- ^ Geraads, D .; El Boughabi, S .; Зухри, С. (2012). «Новый бычий каприн (Mammalia) из позднего миоцена Марокко». Palaeontologica Africana (47): 19–24. ISSN 0078-8554.
- ^ Кингдон, Дж. (1989). Восточноафриканские млекопитающие: атлас эволюции в Африке (том III, часть C). Чикаго: Пресса Чикагского университета. С. 1–33. ISBN 978-0-226-43724-8.
- ^ Стэнли, С. М .; Элдридж, Н. (1984). «Эволюционная модель и процесс в сестринской группе Alcelaphini-Aepycerotini (Mammalia: Bovidae)». Живые ископаемые. Springer. С. 62–79. ISBN 978-1461-382-737.
- ^ Vrba, E. S .; Burckle, L.H .; Partridge, T. C .; Дентон, Г. Х. (1995). Палеоклимат и эволюция с упором на происхождение человека. Нью-Хейвен: издательство Йельского университета. С. 24–5. ISBN 978-0300-063-486.
- ^ а б c Уолтон, Д.В. (1989). Фауна Австралии (Том 1B). Канберра: Служба публикаций правительства Австралии. С. 1–14. ISBN 978-0644-060-561.
- ^ Хасанин, А. (2015). «Систематика и филогения крупного рогатого скота». В Гаррик, Д.; Рувинский, А. (ред.). Генетика крупного рогатого скота (Второе изд.). Оксфордшир, Бостон: Каби. С. 1–18. ISBN 9781780642215.
- ^ Хаффман, Б. "Королевская антилопа". Совершенное копытное животное. Архивировано из оригинал 16 декабря 2014 г.. Получено 8 октября 2014.
- ^ Хилдьярд, А. (2001). Вымирающие виды дикой природы и растений мира. Нью-Йорк: Маршалл Кавендиш. С. 769–70. ISBN 978-0-7614-7200-1.
- ^ "Oryx leucoryx" на Энциклопедия жизни
- ^ а б Lundrigan, B .; Бидлингмейер, Дж. (2000). "Connochaetes gnou: черный гну ". Сеть разнообразия животных. университет Мичигана. Получено 2013-08-21.
- ^ а б c d е ж грамм час я j k л Эстес, Р. Д. (2004). Руководство по поведению африканских млекопитающих: включая копытных, плотоядных и приматов (4-е изд.). Беркли: Калифорнийский университет Press. стр.7–25. ISBN 978-0-520-08085-0.
- ^ Krausman, P.R .; Кейси, А.Л. (2012). "Addax nasomaculatus". Виды млекопитающих. 807: 1–4. Дои:10.1644/807.1.
- ^ а б Каро, Тим (2009). «Контрастная окраска у наземных млекопитающих». Философские труды Королевского общества B. 364 (1516): 537–548. Дои:10.1098 / rstb.2008.0221. ЧВК 2674080. PMID 18990666.
- ^ Котт, Х. (1940). Адаптивная окраска животных. Лондон: Метуэн. С. 88 и лист 25.
- ^ Килти, Р.А. (Январь 1998 г.). «Противозатушевка: универсально обманчивая или обманчиво универсальная?». Тенденции в экологии и эволюции. 3 (1): 21–23. Дои:10.1016/0169-5347(88)90079-1. PMID 21227055.
- ^ Котт, Х. (1940). Адаптивная окраска животных. Лондон: Метуэн. п.53.
- ^ Лесли, Д. М .; Шарма, К. (25 сентября 2009 г.). «Tetracerus quadricornis (Artiodactyla: Bovidae)». Виды млекопитающих. 843: 1–11. Дои:10.1644/843.1.
- ^ E.C., Мунгалл (2007). Полевой справочник экзотических животных: неродные копытные в Соединенных Штатах (1-е изд.). Колледж-Стейшн: издательство Техасского университета A&M. п. 197. ISBN 978-1-58544-555-4.
- ^ Сохранение американских пород домашнего скота (2009). "Джейкоб Баран". Питтсборо, Северная Каролина: Охрана американских пород домашнего скота. Получено 2011-05-05.
- ^ Биби, Ф .; Бухсианидзе, М .; Gentry, A .; Geraads, D .; Kostopoulos, D .; Врба, Э. (2009). «Летопись окаменелостей и эволюция Bovidae: состояние поля». Palaeontologia Electronica. 12 (3): 10А.
- ^ Gatesy, J .; Yelon, D .; DeSalle, R .; Врба, Э. (1992). «Филогения Bovidae (Artiodactyla, Mammalia) на основе последовательностей митохондриальной рибосомной ДНК». Молекулярная биология и эволюция. 9 (3): 433–446. Дои:10.1093 / oxfordjournals.molbev.a040734. PMID 1584013.
- ^ Лундриган, Б. (1996). «Морфология рогов и боевое поведение в семействе Bovidae». Журнал маммологии. 77 (2): 462–75. Дои:10.2307/1382822. JSTOR 1382822.
- ^ Caro, T. M .; Graham, C.M .; Стоунер, С. Дж .; Флорес, М. М. (2003). «Корреляты формы рога и рога у крупного рогатого скота и шейки матки». Поведенческая экология и социобиология. 55 (1): 32–41. Дои:10.1007 / s00265-003-0672-6. S2CID 12030618.
- ^ а б Бро-Йоргенсен, Дж. (2007). «Интенсивность полового отбора определяет размер оружия у самцов крупного рогатого скота». Эволюция. 61 (6): 1316–1326. Дои:10.1111 / j.1558-5646.2007.00111.x. PMID 17542842. S2CID 24278541.
- ^ Эзенва, В. (2008). «Рога честно рекламируют заражение паразитами самцов и самок африканских буйволов». Поведение животных. 75 (6): 2013–21. Дои:10.1016 / j.anbehav.2007.12.013. S2CID 49240459.
- ^ а б Станкович, Т .; Каро, Т. (2009). «Эволюция вооружения у самок крупного рогатого скота». Труды Королевского общества B: биологические науки. 276 (1677): 4329–34. Дои:10.1098 / rspb.2009.1256. ЧВК 2817105. PMID 19759035.
- ^ Пакер, К. (1983). «Половой диморфизм: рога африканских антилоп». Наука. 221 (4616): 1191–3. Bibcode:1983Научный ... 221.1191П. Дои:10.1126 / science.221.4616.1191. PMID 17811523. S2CID 37000507.
- ^ Янис, Ц .; Джарман, П. (1984). Макдональд, Д. (ред.). Энциклопедия млекопитающих. Нью-Йорк: факты в файле. стр.498–9. ISBN 978-0-87196-871-5.
- ^ Вятт, Т. Д. (2003). Феромоны и поведение животных: общение через запах и вкус. Кембридж: Издательство Кембриджского университета. п.97. ISBN 978-0-521-48526-5.
- ^ Чишек, Д. "Бушбак". Сеть разнообразия животных. Музей зоологии Мичиганского университета. Получено 28 октября 2014.
- ^ Т. Л., Ньюэлл. "Уотербак". Сеть разнообразия животных. Музей зоологии Мичиганского университета. Получено 28 октября 2014.
- ^ Лотт, Дейл Ф. (1991). Внутривидовая изменчивость социальных систем диких позвоночных. Издательство Кембриджского университета. стр.37. ISBN 978-0521370240.
- ^ Бигалке, Р. (1972). "Наблюдения за поведением и привычками кормления спрингбока. Antidorcas marsupialis". Zoologica Africana. 7 (1): 333–359. Дои:10.1080/00445096.1972.11447448.
- ^ а б Паркер, С.П. (1990). Энциклопедия млекопитающих Гржимека (том 5) (1-е изд.). Нью-Йорк: McGraw-Hill Publishing. стр.288–324, 338–9, 354–5, 432–3, 444–5, 460–1, 482–3. ISBN 9780079095084.
- ^ Czaplewski, N.J .; Райан, Дж. М .; Воан, Т.А. (2011). Маммология (5-е изд.). Садбери: издательство «Джонс и Бартлетт». ISBN 9780763762995.
- ^ Post, E .; Форчхаммер М.С. (июль 2008 г.). «Изменение климата снижает репродуктивный успех арктических травоядных из-за трофического несоответствия». Философские труды Королевского общества B: биологические науки. 363 (1501): 2367–2373. Дои:10.1098 / rstb.2007.2207. ЧВК 2606787. PMID 18006410.
- ^ Gagnon, M .; Chew, A.E. (май 2000 г.). «Диетические предпочтения современных африканских быков». Журнал маммологии. 81 (2): 490–511. Дои:10.1644 / 1545-1542 (2000) 081 <0490: DPIEAB> 2.0.CO; 2.
- ^ Sponheimer, M .; Lee-Thorp, J.A .; ДеРуитер, Д.Дж .; Smith, J.M .; ван дер Мерве, штат Нью-Джерси; Рид, К .; Grant, C.C .; Ayliffe, L.K .; Робинсон, Т.Ф. (2003). «Рацион южноафриканских быков: данные о стабильных изотопах». Журнал маммологии. 84 (2): 471–9. Дои:10.1644 / 1545-1542 (2003) 084 <0471: DOSABS> 2.0.CO; 2.
- ^ а б c d е Feldhamer, George A .; Дрикамер, Ли С .; Весси, Стивен Х .; Мерритт, Джозеф Ф .; Краевский, Кэри (2007). Маммология: адаптация, разнообразие, экология. Издательство Университета Джона Хопкинса. С. 519–522. ISBN 978-0-8018-8695-9.
- ^ а б Krebs, J.R .; Дэвис, Н. (1997). Поведенческая экология: эволюционный подход (4-е изд.). Вили-Блэквелл. ISBN 9780865427310.
- ^ а б Эстес, Р. Д. (2004). Руководство по поведению африканских млекопитающих: включая копытных, плотоядных и приматов (4-е изд.). Беркли: Калифорнийский университет Press. стр.158–66. ISBN 978-0-520-08085-0.
- ^ Грей, Гэри Дж .; Симпсон, К. Дэвид (1980). "Ammotragus lervia". Виды млекопитающих. 1980 (144): 1–7. Дои:10.2307/3504009. JSTOR 3504009.
- ^ Rosa, H.J.D .; Брайант, М.Дж. (2003). «Сезонность воспроизводства овец». Исследования мелких жвачных животных. 48 (3): 155–71. Дои:10.1016 / S0921-4488 (03) 00038-5.
- ^ Chemineau, P .; Pelletier, J .; Guérin, Y .; Colas, G .; Ravault, J.P .; Touré, G .; Almeida, G .; Thimonier, J .; Ортавант Р. (1988). «Фотопериодические препараты и лечение мелатонином для контроля сезонного воспроизводства овец и коз». Размножение, питание, развитие. 28 (2B): 409–22. Дои:10.1051 / номер: 19880307. PMID 3413339.
- ^ Prendergast, B.J .; Мозингер, Б .; Колаттукуды, П. Э .; Нельсон, Р. Дж. (2002). «Экспрессия гена гипоталамуса у репродуктивно фотореактивных и фоторефрактерных сибирских хомяков». Труды Национальной академии наук. 99 (25): 16291–6. Bibcode:2002PNAS ... 9916291P. Дои:10.1073 / pnas.232490799. ЧВК 138604. PMID 12456888.
- ^ Оуэн-Смит, Н. (1993). «Сравнительные показатели смертности мужских и женских куду: цена полового диморфизма размера». Журнал экологии животных. 62 (3): 428–40. Дои:10.2307/5192. JSTOR 5192.
- ^ Toigo, C .; Гайяр, Дж. М. (2003). «Причины выживания взрослых копытных животных с ориентацией на пол: диморфизм размеров пола, тактика спаривания или суровость окружающей среды?». Ойкос. 101 (2): 376–84. Дои:10.1034 / j.1600-0706.2003.12073.x.
- ^ Эстес, Р. Д. (1991). «Значение рогов и других мужских вторичных половых признаков у самок крупного рогатого скота». Прикладная наука о поведении животных. 29 (1–4): 403–51. Дои:10.1016 / 0168-1591 (91) 90264-Х.
- ^ Цедер, М.А. (2006). Документирование приручения: новые генетические и археологические парадигмы. Беркли, Калифорния: Калифорнийский университет Press. п. 317. ISBN 978-0520246386.
- ^ Фелан, Бенджамин; Фелан, Бенджамин (24 июля 2013 г.). «Чужое молоко». Slate.com. Получено 10 октября 2014.
- ^ Хьюз, Том; Хьюз, Мередит Сэйлс (2005). Gastronomie !: Кулинарные музеи и объекты культурного наследия Франции. Банкер Хилл Паблишинг. п. 19. ISBN 978-1-59373-029-1.
- ^ «Говядина нежирная органическая». WHFoods. 18 октября 2004 г.. Получено 1 апреля 2015.
- ^ "| Национальная ассоциация зубров". Bisoncentral.com. В архиве из оригинала от 20 января 2011 г.. Получено 1 апреля 2015.
- ^ Veldmeijer, A.J .; Харрис, С. (2014). Почему кожа?: Материальные и культурные аспекты кожи. Sidestone Press. С. 31–6. ISBN 9789088902611.
- ^ "Овцы-мериносы в Австралии". Архивировано из оригинал на 2006-11-05. Получено 1 апреля 2015.
- ^ Колай, А. К. (2007). Навоз и удобрения. Нью-Дели: Атлантические публикации. п. 98. ISBN 978-8126908103.
- ^ Ви, Э. (1975). «Германские стеклянные рожки для питья». Журнал исследований стекла. 17: 74–87.
- ^ "Басни Эзопа". Басни Эзопа. Получено 10 октября 2014.
- ^ Клевать. «Запись: Химера». Получено 31 марта 2015.
- ^ Эберхард, В. Словарь китайских символов: скрытые символы в китайской жизни и мысли. Лондон: Рутледж. ISBN 978-0-415-00228-8.
внешняя ссылка
- . Британская энциклопедия (11-е изд.). 1911 г.
- . Новая энциклопедия Кольера. 1921.