Микронуклеус - Micronucleus
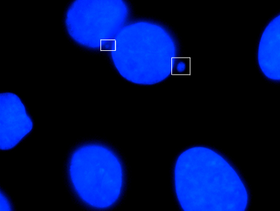
Микронуклеус это название, данное маленькому ядру, которое образуется всякий раз, когда хромосома или фрагмент хромосомы не включается в одно из дочерних ядер во время деления клетки. Обычно это признак генотоксических событий и хромосомной нестабильности. Микроядра обычно наблюдаются в раковых клетках и могут указывать на события повреждения генома, которые могут увеличить риск развития или дегенеративных заболеваний.[1] Микроядра образуются во время анафазы из-за отставания ацентрической хромосомы или фрагментов хроматид, вызванных неправильно или нереставрированными разрывами ДНК или нерасхождением хромосом. Эта неправильная сегрегация хромосом может быть результатом гипометилирования повторяющихся последовательностей, присутствующих в перицентромерной ДНК, нарушений в белках кинетохор или их сборки, дисфункционального аппарата веретена или дефектных генов контрольных точек анафазы.[2] Многие микроядерные тесты были разработаны для проверки наличия этих структур и определения их частоты в клетках, подвергшихся воздействию определенных химических веществ или стрессовых условиях.
Период, термин микронуклеус может также относиться к меньшему ядро в инфузория простейшие, такой как Парамеций. В деление он делится на митоз, И в спряжение это обеспечивает сочетание гамета ядра, взаимное слияние которых зигота образуется ядро, которое дает начало макронуклеусы и микроядра особей следующего цикла деления.[3]
Открытие
Микроядра во вновь образованных красных кровяных тельцах человека известны как Тела Хауэлл-Джолли потому что эти структуры были впервые идентифицированы и описаны в эритроцитах гематологами Уильямом Хауэллом и Джастином Джолли. Позже было обнаружено, что эти структуры связаны с дефицитом витаминов, таких как фолиевая кислота и B12. Взаимосвязь между образованием микроядер и воздействием факторов окружающей среды была впервые обнаружена в клетках кончика корня, подвергшихся воздействию ионизирующего излучения. Об индукции микроядер химическим веществом впервые сообщили в клетках асцитной опухоли Эрлиха, обработанных колхицином.[2]
Формирование
Микроядра в основном возникают в результате ацентрических фрагментов хромосом или отставания целых хромосом, которые не включены в дочерние ядра, образованные в результате митоза, потому что они не могут правильно прикрепиться к веретену во время сегрегации хромосом в анафазе. Эти полные хромосомы или фрагменты хроматид в конечном итоге заключены в ядерные мембраны и структурно похожи на обычные ядра, хотя и меньше по размеру. Это маленькое ядро называется микроядром. Формирование микроядер можно наблюдать только в клетках, подвергающихся делению ядра, и его можно четко увидеть с помощью цитохалазин B блокировать цитокинез и производить двухъядерные клетки.[2]
Ацентрические фрагменты хромосом могут возникать по-разному. Один из способов состоит в том, что нарушение целостности двухцепочечных разрывов ДНК может приводить к симметричным или асимметричным обменам хроматид и хромосом, а также к хроматидным и хромосомным фрагментам. Если повреждение ДНК превышает репарационную способность клетки, нерепарированные двухцепочечные разрывы ДНК также могут привести к ацентрическим фрагментам хромосом. Еще один способ возникновения эксцентрических фрагментов хромосом - это когда дефекты в генах, связанные с гомологичной рекомбинационной репарацией (например, ATM, BRCA1, BRCA2 и RAD51), приводят к дисфункциональному безошибочному пути репарации гомологичной рекомбинационной ДНК и заставляют клетку прибегать к ошибке. -prone non-homologous end-joining (NHEJ) путь репарации, повышающий вероятность неправильной репарации разрывов ДНК, образования дицентрических хромосом и ацентрических фрагментов хромосом. Если ферменты в пути репарации NHEJ также являются дефектными, разрывы ДНК могут вообще не исправляться. Кроме того, одновременная эксцизионная репарация поврежденных или несоответствующих оснований, включенных в ДНК, которые находятся в непосредственной близости и на противоположных комплементарных цепях ДНК, может приводить к двухцепочечным разрывам ДНК и образованию микроядер, особенно если этап заполнения пробелов в пути репарации не завершен.[2]
Микроядра могут также образовываться из фрагментированных хромосом, когда нуклеоплазматические мостики (NPB) образуются, растягиваются и разрушаются во время телофазы.[2]
Образование микроядер также может быть результатом неправильной сегрегации хромосом во время анафазы. Гипометилирование цитозина в центромерных и перицентромерных областях и повторы более высокого порядка сателлитной ДНК в центромерной ДНК могут приводить к таким событиям хромосомной потери. Классическая сателлитная ДНК обычно сильно метилирована по остаткам цитозина, но может стать почти полностью неметилированной из-за Синдром МКФ (Иммунодефицит, нестабильность центромеры и синдром лицевых аномалий) или после лечения ингибиторами ДНК-метилтрансферазы. С момента сборки кинетохора На белки центромер влияет метилирование цитозиновых и гистоновых белков, снижение целостности гетерохроматина в результате гипометилирования может мешать прикреплению микротрубочек к хромосомам и ощущению натяжения правильных соединений микротрубочки-кинетохоры. Другими возможными причинами потери хромосом, которые могут привести к образованию микроядер, являются дефекты взаимодействия кинетохор и микротрубочек, дефекты сборки митотического веретена, дефекты контрольных точек митоза, аномальная амплификация центросом и слияния концов теломеров, которые приводят к тому, что дицентрические хромосомы отделяются от веретена во время анафаза. Микроядра, происходящие из событий потери хромосом и ацентрических фрагментов хромосом, можно различить с помощью панцентромерных ДНК-зондов.[2]
Идентификация
Количество микроядер на клетку можно предсказать, используя следующую формулу:
AF - количество ацентрических фрагментов и F = 0,5 - 0,5P, где P равно вероятности того, что фрагменты включены в традиционное ядро и не образуют микроядра.[4]

Одно исследование, в котором для окрашивания ядерного материала использовалось окрашивание по Гимзе, установило следующие критерии для идентификации микроядер:
1) диаметр менее 1/3 первичного ядра,
2) отсутствие ретракции (исключает мелкие частицы пятен),
3) цвет такой же или светлее, чем основное ядро (за исключением крупных пятен),
4) расположение в пределах 3 или 4 ядерных диаметров от основного ядра, не касаясь его, и
5) не более двух, связанных с одним первичным ядром (3 или более микроядра, вероятно, являются полиморфами или прорубицитами с ядерными фрагментами).[5]
Анализы
Микроядерные тесты предоставляют важную информацию о способности химического вещества влиять на структуру и функцию хромосом. Например, многие известные канцерогены человека дают положительный результат в тестах на микроядра млекопитающих. В этих тестах организмы обрабатываются химическим веществом и измеряется результирующая частота появления микроядер. Если наблюдается заметное увеличение количества клеток с микроядрами, можно сделать вывод, что химическое вещество вызывает структурные и / или числовые хромосомные повреждения. Поскольку тесты на микроядра должны проводиться на активно делящихся клетках, стволовые клетки костного мозга и эритроциты, которые они производят при делении клеток, являются идеальными кандидатами. Эти клетки испытывают постоянный, быстрый оборот, а отсутствие настоящего ядра в эритроцитах делает микроядра легко видимыми под микроскопом.[1]
Системы анализа микроядер очень экономичны, требуют гораздо меньших навыков при подсчете баллов, чем обычные метафазные тесты, и намного быстрее, чем эти обычные тесты. Поскольку микронуклеусные анализы надежно и быстро отражают хромосомные аберрации, они чрезвычайно полезны для быстрой оценки хромосомных повреждений. В частности, анализ CBMNcyt (цитокинез-блокирующий микроядерный цитом) чрезвычайно универсален и является одним из предпочтительных методов измерения уровня хромосомного повреждения и хромосомной нестабильности в клетках. Анализ микроядер с блокировкой цитокинеза (CBMN) был впервые разработан для оценки микроядер в клетках, завершивших деление ядра, путем их блокирования на стадии биядерных ядер перед цитокинезом. Позже он превратился в «цитомный» анализ CBMN для дальнейшего изучения гибели клеток, цитостаза и биомаркеров повреждения ДНК. Основным недостатком использования микроядерных тестов является то, что они не могут определять различные типы хромосомных аберраций и могут зависеть от скорости митоза и доли гибели клеток, искажая результаты.[2]
Паттерны в формировании
Многочисленные исследования показали, что частота микроядер у женщин выше, чем у мужчин, и что количество микроядер увеличивается примерно до 70 лет. Уровни микроядер колеблются от 0,5 до 1,4% у мужчин и от 0,9 до 1,8% у женщин. Гендерные различия в основном наблюдались в более молодых возрастных группах (<= 50 лет) с почти двукратной разницей между мужчинами и женщинами. Структура количества микроядер после 70 лет противоречива. Некоторые исследования показали, что у лиц старше 70 лет частота микроядер увеличивается у обоих полов. С другой стороны, другие исследования показали, что в самых старших возрастных группах частота микроядер выравнивается. Дефицит микроядер в некоторых наиболее старших возрастных группах можно объяснить тем фактом, что микроядерные клетки преимущественно уничтожаются апоптозом. Однако более высокая частота микроядер соответствует снижению эффективности репарации ДНК и повышенной нестабильности генома, что типично для пожилых людей. Возрастное увеличение частоты микроядер также хорошо согласуется с возрастным увеличением гипоплоидии и возрастным увеличением потери половых хромосом. В качестве альтернативы, выравнивание частоты появления микроядер у пожилых людей могло бы предложить порог геномной нестабильности, который нельзя преодолеть, если человек хочет выжить. Если бы это было так, женщины, кажется, достигли этого порога быстрее, чем мужчины.[6]
Половые хромосомы вносят вклад в большинство случаев потери хромосом с возрастом. У женщин Х-хромосома может составлять до 72% наблюдаемых микроядер, из которых 37%, по-видимому, лишены функциональной сборки кинетохор, возможно, из-за инактивации Х-хромосомы. Многочисленные исследования показали, что частота аутосомно-положительных микроядер у обоих полов и положительных по половым хромосомам МЯ у мужчин была сходной и оставалась неизменной в более старших группах, в то время как частота Х-положительных МН у женщин была выше, чем средняя частота аутосомных -положительный МН и продолжал увеличиваться до самого старшего возраста.[2]
Частота хромосомных аберраций, поврежденных клеток и микроядер значительно выше у курильщиков, чем у некурящих.[7]
У нормальных людей и многих других млекопитающих, у которых нет ядер в эритроцитах, микроядра быстро удаляются селезенка. Следовательно, высокая частота микроядер в периферической крови человека указывает на разрыв или отсутствие селезенки. У мышей они не удаляются, что является основанием для in vivo микронуклеусный тест.
Смотрите также
Рекомендации
- ^ а б «Микронуклеус». ntp.niehs.nih.gov. Архивировано из оригинал на 2016-10-18. Получено 2016-10-14.
- ^ а б c d е ж грамм час Fenech, M .; Кирш-Волдерс, М .; Натараджан, А. Т .; Surralles, J .; Crott, J. W .; Parry, J .; Norppa, H .; Истмонд, Д. А .; Такер, Дж. Д. (01.01.2011). «Молекулярные механизмы образования микроядер, нуклеоплазматических мостиков и ядерных зачатков в клетках млекопитающих и человека». Мутагенез. 26 (1): 125–132. Дои:10.1093 / mutage / geq052. ISSN 0267-8357. PMID 21164193.
- ^
 Одно или несколько предыдущих предложений включают текст из публикации, которая сейчас находится в всеобщее достояние: Чисхолм, Хью, изд. (1911). "Микронуклеус ". Британская энциклопедия. 18 (11-е изд.). Издательство Кембриджского университета. п. 391.
Одно или несколько предыдущих предложений включают текст из публикации, которая сейчас находится в всеобщее достояние: Чисхолм, Хью, изд. (1911). "Микронуклеус ". Британская энциклопедия. 18 (11-е изд.). Издательство Кембриджского университета. п. 391. - ^ Сэвидж, Джон Р. К. (1988-01-01). «Комментарий к количественной связи между микроядрами и хромосомными аберрациями». Письма об исследованиях мутаций. 207 (1): 33–36. Дои:10.1016/0165-7992(88)90008-5. PMID 3336377.
- ^ Земляк, Павел I .; Хеддл, Джон А. (1976-12-01). «Производство микроядер из хромосомных аберраций в облученных культурах лимфоцитов человека». Мутационные исследования / Фундаментальные и молекулярные механизмы мутагенеза. 41 (2–3): 321–331. Дои:10.1016/0027-5107(76)90105-6.
- ^ Войда, Алина; Зенткевич, Ева; Витт, Михал (01.05.2007). «Влияние возраста и пола на частоту нерасхождения микроядер и хромосом у людей долгожительства и более молодого возраста». Мутагенез. 22 (3): 195–200. Дои:10.1093 / mutage / gem002. ISSN 0267-8357. PMID 17284771.
- ^ Бандана Гангули, Бани (1993-08-01). «Деление клеток, хромосомные повреждения и образование микроядер в периферических лимфоцитах здоровых доноров: в зависимости от возраста донора». Исследование мутаций / ДНК-анализ. 295 (3): 135–148. Дои:10.1016 / 0921-8734 (93) 90015-У. PMID 7689700.