Растительная клетка - Plant cell
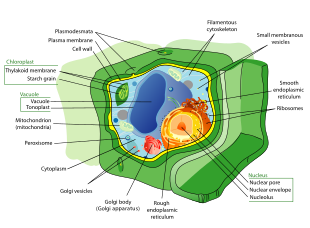
Клетки растений находятся эукариотический клетки присутствуют в зеленые растения, фотосинтезирующие эукариоты королевства Plantae. Их отличительными особенностями являются первичные клеточные стенки, содержащие целлюлозу, гемицеллюлозы и пектин, наличие пластид, способных осуществлять фотосинтез и накапливать крахмал, большое количество вакуоль регулирующего тургорное давление, отсутствие жгутики или же центриоли, за исключением гамет, и уникальный метод деления клеток, включающий образование клеточной пластинки или фрагмопласт который отделяет новые дочерние клетки.
Характеристики растительных клеток
- Растительные клетки имеют клеточные стенки, построенный вне клеточная мембрана и состоит из целлюлоза, гемицеллюлозы, и пектин. Их состав контрастирует с клеточными стенками грибы, которые сделаны из хитин, из бактерии, которые сделаны из пептидогликан и из археи, которые сделаны из псевдопептидогликан. Во многих случаях лигнин или же Суберин секретируются протопласт как вторичные слои стенки внутри первичной клеточной стенки. Кутин секретируется за пределы первичной клеточной стенки и во внешние слои вторичной клеточной стенки эпидермальных клеток листьев, стеблей и других надземных органов с образованием кутикула растений. Клеточные стенки выполняют множество важных функций. Они придают форму тканям и органам растения и играют важную роль в межклеточной коммуникации и взаимодействиях между растениями и микробами.[1]
- Многие типы растительных клеток содержат большой центральный вакуоль, заполненный водой объем, окруженный мембраной, известной как тонопласт[2] который поддерживает клетки тургор, контролирует движение молекулы между цитозоль и сок, хранит полезные материалы, такие как фосфор и азот [3] и переваривает отходы белки и органеллы.
- Специализированные пути межклеточной коммуникации, известные как плазмодесматы,[4] возникают в виде пор в первичной клеточной стенке, через которые плазмалемма и эндоплазматический ретикулум[5] смежных ячеек непрерывны.
- Клетки растений содержат пластиды, наиболее заметное существо хлоропласты, содержащие пигмент зеленого цвета хлорофилл который преобразует энергию солнечного света в химическую энергию, которую растение использует для производства собственной пищи из воды и углекислого газа в процессе, известном как фотосинтез.[6] Другие типы пластид - это амилопласты, специализированный для крахмал место хранения, элайопласты специализированный для толстый хранение и хромопласты специализируется на синтезе и хранении пигменты. Как в митохондрии, геном которых кодирует 37 генов,[7] пластиды имеют свои геномы около 100–120 уникальных гены[8] и интерпретируются как возникшие как прокариотический эндосимбионты живущих в камерах раннего эукариотический предок наземные растения и водоросли.[9]
- Деление клеток у наземных растений и некоторых групп водорослей, особенно Харофиты[10] и Хлорофит Заказ Trentepohliales,[11]происходит путем строительства фрагмопласт как шаблон для построения клеточная пластина в конце цитокинез.
- Подвижное, свободное плавание сперма из мохообразные и птеридофиты, саговники и Гинкго являются единственными клетками наземных растений, которые имеют жгутики[12] аналогично тем, что в клетки животных,[13][14] но хвойные породы и цветущие растения не имеют подвижных сперматозоидов и лишены жгутиков и центриоли.[15]
Типы растительных клеток и тканей
Клетки растений дифференцируются от недифференцированных меристематический клетки (аналогичные стволовым клеткам животных), чтобы сформировать основные классы клеток и тканей корни, стебли, листья, цветы и репродуктивные структуры, каждая из которых может состоять из нескольких типов клеток.
Паренхима
Клетки паренхимы живые клетки, у которых есть функции от хранения и поддержки до фотосинтез (мезофилл клетки) и загрузка флоэмы (передаточные клетки ). Помимо ксилемы и флоэмы в их сосудистых пучках, листья состоят в основном из клеток паренхимы. Некоторые клетки паренхимы, например эпидермис, специализируются на проникновении света и фокусировании или регулировании газообмен, но другие относятся к наименее специализированным клеткам в растительной ткани и могут оставаться тотипотент, способные делиться с образованием новых популяций недифференцированных клеток на протяжении всей своей жизни.[16] Клетки паренхимы имеют тонкие, проницаемые первичные стенки, позволяющие переносить небольшие молекулы между ними, а их цитоплазма отвечает за широкий спектр биохимических функций, таких как нектар секреция, или изготовление вторичные продукты это обескураживает травоядный. Клетки паренхимы, которые содержат много хлоропластов и связаны в первую очередь с фотосинтезом, называются хлоренхима клетки. Другие, такие как большинство клеток паренхимы в картофель клубни и семя семядоли из бобовые, имеют функцию хранения.
Колленхима
Клетки колленхимы - клетки колленхимы живы в зрелом возрасте и имеют утолщенные клеточные стенки целлюлозы.[17] Эти клетки созревают из производных меристемы, которые изначально напоминают паренхиму, но различия быстро становятся очевидными. Пластиды не развиваются, а секреторный аппарат (ER и Golgi) разрастается, секретируя дополнительную первичную стенку. Стенка обычно наиболее толстая в углах, где соприкасаются три или более ячеек, и самая тонкая, когда соприкасаются только две ячейки, хотя возможны и другие способы утолщения стенки.[17] Пектин и гемицеллюлоза являются доминирующими составляющими клеточных стенок колленхимы двудольные покрытосеменные, который может содержать всего 20% целлюлозы в Петаситы.[18] Клетки колленхимы, как правило, довольно удлиненные и могут делиться поперечно, создавая вид перегородок. Роль этого типа клеток состоит в том, чтобы поддерживать растение в осях, все еще растущих в длину, и придавать тканям гибкость и прочность на разрыв. В первичной стенке отсутствует лигнин, который сделал бы ее прочной и жесткой, поэтому этот тип клеток обеспечивает то, что можно было бы назвать пластиковой опорой - опорой, которая может удерживать молодой стебель или черешок в воздухе, но в клетках, которые могут растягиваться по мере того, как клетки вокруг них удлиненный. Эластичная опора (без эластичной защелки) - хороший способ описать, что делает колленхима. Части нитей сельдерея - колленхима.

Склеренхима
Склеренхима представляет собой ткань, состоящую из двух типов клеток, склереиды и волокна которые загустели, одревесневший второстепенные стены[17]:78 заложен внутри первичная клеточная стенка. Вторичные стенки укрепляют клетки и делают их непроницаемыми для воды. Следовательно, скереиды и волокна обычно мертвы при функциональной зрелости, а цитоплазма отсутствует, оставляя пустую центральную полость. Склероиды или каменные кельи (от греч. склерос, жесткий) представляют собой твердые твердые клетки, которые придают листьям или фруктам песчаную текстуру. Они могут препятствовать развитию травоядных, повреждая пищеварительные тракты на личиночных стадиях мелких насекомых. Склериды образуют твердую стенку косточки персиков и многих других фруктов, обеспечивая физическую защиту развивающегося ядра. Волокна представляют собой удлиненные клетки с одревесневшими вторичными стенками, которые обеспечивают несущую поддержку и прочность на разрыв листья и стебли травянистых растений. Волокна склеренхимы не участвуют в проведении ни воды, ни питательных веществ (как в ксилема ) или углеродных соединений (как в флоэма ), но вполне вероятно, что они возникли как модификации инициалов ксилемы и флоэмы у ранних наземных растений.

Ксилема
Ксилема представляет собой сложную сосудистую ткань, состоящую из проводящих воду трахеиды или же элементы судна вместе с волокнами и клетками паренхимы. Трахеиды [19] представляют собой удлиненные клетки с одревесневшим вторичным утолщением клеточных стенок, специализированные для проведения воды и впервые появившиеся у растений во время их перехода на сушу в Силурийский период более 425 миллионов лет назад (см. Cooksonia ). Наличие ксилемы трахеид определяет сосудистые растения или трахеофиты. Трахеиды представляют собой заостренные, удлиненные клетки ксилемы, самые простые из которых имеют непрерывные первичные клеточные стенки и одревесневшие вторичные утолщения стенок в виде колец, обручей или сетчатых сетей. Более сложные трахеиды с клапанными перфорациями называются окаймленные ямы характеризуют голосеменные. В папоротники и другие птеридофиты и голосеменные есть только ксилема трахеиды, в то время как цветущие растения Также есть ксилемные сосуды. Элементы сосуда представляют собой полые клетки ксилемы без торцевых стенок, которые выровнены встык так, чтобы образовывать длинные непрерывные трубки. У мохообразных отсутствует настоящая ткань ксилемы, но их спорофиты имеют водопроводящую ткань, известную как гидром, которая состоит из удлиненных клеток более простой конструкции.
Флоэма
Флоэма это специализированная ткань для транспортировки пищи у высших растений, в основном транспортирующая сахароза вдоль градиентов давления, создаваемых осмосом, процесс, называемый перемещение. Флоэма представляет собой сложную ткань, состоящую из двух основных типов клеток: ситовые трубки и тесно связанный компаньоны вместе с клетками паренхимы, волокнами флоэмы и склероидами.[17]:171 Ситчатые трубы соединены встык с перфорированными концевыми пластинами между известными как сита, которые позволяют переносить фотосинтез между элементами сита. В элементах ситовой трубки отсутствуют ядра и рибосомы, а их метаболизм и функции регулируются соседними ядросодержащими клетками-компаньонами. Сопутствующие ячейки, подключенные к ситчатым трубкам через плазмодесматы, отвечают за загрузку флоэмы сахара. В мохообразные не хватает флоэмы, но мох спорофиты имеют более простую ткань с аналогичной функцией, известную как лептом.

Эпидермис
В эпидермис растений представляет собой специализированную ткань, состоящую из клеток паренхимы, которая покрывает внешние поверхности листьев, стеблей и корней. В эпидермисе могут присутствовать несколько типов клеток. Среди них следует отметить замыкающие клетки устьиц, которые контролируют скорость газообмен между растением и атмосферой, железистые волосы и волосы на одежде или трихомы, а корневые волоски первичных корней. В эпидермисе побегов большинства растений только замыкающие клетки есть хлоропласты. Хлоропласты содержат зеленый пигмент хлорофилл, необходимый для фотосинтеза. Эпидермальные клетки надземных органов возникают из поверхностного слоя клеток, известного как туника (Слои L1 и L2), покрывающий растение стрелять апексом,[17] тогда как кора и сосудистые ткани возникают из самого внутреннего слоя верхушки побега, известного как корпус (Слой L3). Эпидермис корней образуется из слоя клеток, находящегося непосредственно под корневым покровом. Эпидермис всех надземных органов, кроме корней, покрыт кутикула сделано из полиэстер Cutin или полимер Cutan (или оба), с поверхностным слоем эпикутикулярные воски. Считается, что эпидермальные клетки первичного побега являются единственными растительными клетками, обладающими биохимической способностью синтезировать кутин.[20]
Смотрите также
Рекомендации
- ^ Кигстра, К. (2010). «Стенки растительных клеток». Физиология растений. 154 (2): 483–486. Дои:10.1104 / стр.110.161240. ЧВК 2949028. PMID 20921169.
- ^ Ворон, JA (1997). «Вакуоль: анализ затрат и выгод». Достижения в ботанических исследованиях. 25: 59–86. Дои:10.1016 / S0065-2296 (08) 60148-2. ISBN 9780120059256.
- ^ Рэйвен, Дж. (1987). «Роль вакуолей». Новый Фитолог. 106 (3): 357–422. Дои:10.1111 / j.1469-8137.1987.tb00149.x.
- ^ Опарка, KJ (1993). «Передача сигналов через плазмодесматы - запущенный путь». Семинары по клеточной биологии. 4 (2): 131–138. Дои:10.1006 / scel.1993.1016. PMID 8318697.
- ^ Хеплер, ПК (1982). «Эндоплазматический ретикулум в формировании клеточной пластинки и плазмодесм». Протоплазма. 111 (2): 121–133. Дои:10.1007 / BF01282070. S2CID 8650433.
- ^ Бассхэм, Джеймс Алан; Lambers, Hans, eds. (2018). «Фотосинтез: важность, процесс и реакции». Энциклопедия Британника. Получено 2018-04-15.
- ^ Андерсон, S; Банкир, AT; Barrell, BG; de Bruijn, MH; Колсон, АР; Друин, Дж; Eperon, IC; Nierlich, DP; Роу, BA; Sanger, F; Шрайер, PH; Смит, AJ; Staden, R; Янг, И.Г. (1981). «Последовательность и организация митохондриального генома человека». Природа. 290 (5806): 4–65. Bibcode:1981Натура.290..457A. Дои:10.1038 / 290457a0. PMID 7219534. S2CID 4355527.
- ^ Cui, L; Veeraraghavan, N; Рихтер, А; Стена, К; Янсен, РК; Либенс-Мак, Дж; Макаловская, I; dePamphilis, CW (2006). «ChloroplastDB: база данных генома хлоропластов». Исследования нуклеиновых кислот. 34 (90001): D692-696. Дои:10.1093 / nar / gkj055. ЧВК 1347418. PMID 16381961.
- ^ Маргулис, L (1970). Происхождение эукариотических клеток. Нью-Хейвен: издательство Йельского университета. ISBN 978-0300013535.
- ^ Льюис, штат Луизиана; Маккорт, РМ (2004). «Зеленые водоросли и происхождение наземных растений» (PDF). Американский журнал ботаники. 91 (10): 1535–1556. Дои:10.3732 / ajb.91.10.1535. PMID 21652308.
- ^ Лопес-Баутиста, JM; Waters, DA; Чепмен, Р.Л. (2003). «Фрагмопластин, зеленые водоросли и эволюция цитокинеза». Международный журнал систематической и эволюционной микробиологии. 53 (6): 1715–1718. Дои:10.1099 / ijs.0.02561-0. PMID 14657098.
- ^ Silflow, CD; Лефевр, Пенсильвания (2001). «Сборка и подвижность ресничек и жгутиков эукариот. Уроки Хламидомонада Reinhardtii". Физиология растений. 127 (4): 1500–1507. Дои:10.1104 / стр.010807. ЧВК 1540183. PMID 11743094.
- ^ Мантон, I; Кларк, Б. (1952). "Электронно-микроскопическое исследование сперматозоида Сфагнум". Журнал экспериментальной ботаники. 3 (3): 265–275. Дои:10.1093 / jxb / 3.3.265.
- ^ Паолилло младший, диджей (1967). «О строении аксонемы жгутиков Polytrichum juniperinum". Труды Американского микроскопического общества. 86 (4): 428–433. Дои:10.2307/3224266. JSTOR 3224266.
- ^ Ворон, PH; Эверт, РФ; Eichhorm, SE (1999). Биология растений (6-е изд.). Нью-Йорк: W.H. Фримен. ISBN 9780716762843.
- ^ Г., Хаберланд (1902). "Kulturversuche mit изолированное Pflanzenzellen". Mathematisch-naturwissenschaftliche. Akademie der Wissenschaften в Wien Sitzungsberichte. 111 (1): 69–92.
- ^ а б c d е Каттер, EG (1977). Анатомия растений. Часть 1. Клетки и ткани.. Лондон: Эдвард Арнольд. ISBN 0713126388.
- ^ Рулофсен, PA (1959). Стенка растительной клетки. Берлин: Gebrüder Borntraeger. КАК В B0007J57W0.
- ^ MT Тайри; MH Zimmermann (2003) Структура ксилемы и подъем сока, 2-е издание, Springer-Verlag, Нью-Йорк, США
- ^ Колаттукуди, PE (1996) Биосинтетические пути кутина и воска и их чувствительность к экологические стрессы. В: Кутикулы растений. Эд. Дж. Керстиенс, BIOS Scientific publishers Ltd., Оксфорд, стр. 83–108
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Растение группы | |||||||||||
| |||||||||||
| |||||||||||
| Рост и привычка растений | |||||||||||
| Размножение | |||||||||||
| Таксономия растений | |||||||||||
| Упражняться | |||||||||||
| |||||||||||
| |||||||||||