Красный импортный огненный муравей - Red imported fire ant - Wikipedia
| Красный импортный огненный муравей | |
|---|---|
 | |
| Группа рабочих-пожарных муравьев | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Членистоногие |
| Учебный класс: | Насекомое |
| Заказ: | Перепончатокрылые |
| Семья: | Formicidae |
| Подсемейство: | Myrmicinae |
| Род: | Соленопсис |
| Разновидность: | S. invicta |
| Биномиальное имя | |
| Solenopsis invicta Бурен, 1972 г. | |
| Синонимы[1] | |
| |
В красный импортный огненный муравей (Solenopsis invicta), также известный как огненный муравей или же РИФА, это вид муравьев, обитающий в Южной Америке. Член рода Соленопсис в подсемействе Myrmicinae, это было описанный швейцарского энтомолога Феликс Сански как вариант в 1916 году. специфический имя invicta был передан муравью в 1972 г. как отдельный вид. Однако вариант и вид были одним и тем же муравьем, и название сохранилось благодаря его широкому использованию. Красный импортированный огненный муравей, хотя и имеет южноамериканское происхождение, был случайно завезен в Австралию, Новую Зеландию, несколько стран Азии и Карибского бассейна, а также в США. Красный импортный огненный муравей полиморфный, поскольку рабочие появляются разных форм и размеров. Цвет муравья красный и несколько желтоватый с коричневым или черным. желудок, но самцы полностью черные. Красные завезенные огненные муравьи доминируют на измененных территориях и живут в самых разных средах обитания. Их можно найти в тропических лесах, нарушенных территориях, пустынях, лугах, вдоль дорог и зданий, а также в электрическом оборудовании. Колонии образуют большие насыпи, построенные из почвы без видимых входов, потому что построены кормовые туннели, и рабочие выходят далеко от гнезда.
Эти муравьи демонстрируют самые разные виды поведения, например строят плоты, когда чувствуют, что уровень воды повышается. Они также показывают некрофорное поведение, где сокамерники выбрасывают обрезки или мертвых муравьев на кучи мусора за пределами гнезда. Собирательство происходит в теплые или жаркие дни, хотя ночью они могут оставаться на улице. Рабочие общаются серией полухимикаты и феромоны, которые используются для вербовки, сбора пищи и защиты. Они есть всеядные и есть мертвых млекопитающих, членистоногих, насекомых, семена и сладкие вещества, такие как нектар из двукрылое животное насекомые, с которыми они развились отношения. Хищники включают паукообразных, птиц и многих насекомых, включая других муравьев, стрекоз, уховерток и жуков. Муравей является хозяином паразитов и ряда патогенов, нематод и вирусов, которые рассматриваются как потенциальные агенты биологической борьбы. Брачный полет происходит в теплое время года, а крылатые может спариваться до 30 минут. Основание колонии может быть осуществлено одной маткой или группой королев, которые позже будут бороться за господство, как только появятся первые рабочие. Рабочие могут жить несколько месяцев, а королевы - годами; количество колоний может варьироваться от 100 000 до 250 000 особей. У красного импортированного огненного муравья существуют две формы общества: полигинные колонии (гнезда с несколькими матками) и моногинные колонии (гнезда с одной маткой).
Яд играет важную роль в жизни муравья, так как он используется для поимки добычи или для защиты.[2] Около 95% яда состоит из нерастворимых в воде пиперидин алкалоиды известный как соленопсины, остальная часть включает смесь токсичных белков, которые могут быть особенно эффективными для чувствительных людей. Ежегодно в Соединенных Штатах от них ужаливают более 14 миллионов человек, и ожидается, что у многих из них разовьется аллергия на этот яд. Большинство пострадавших испытывают сильное жжение и припухлость с последующим образованием бесплодия. пустулы, который может сохраняться несколько дней. Однако от 0,6% до 6,0% людей могут страдать от анафилаксия, который может быть фатальным, если его не лечить. Общие симптомы включают головокружение, боль в груди, тошноту, сильное потоотделение, низкое кровяное давление, одышку и невнятную речь. От нападений красных завезенных огненных муравьев зарегистрировано более 80 смертей. Лечение зависит от симптомов; тем, кто испытывает только боль и образование пустул, не требуется медицинской помощи, но тем, кто страдает анафилаксией, дается эпинефрины. Иммунотерапия экстрактом всего тела используется для лечения пострадавших и считается высокоэффективной.[нужна цитата ]
Муравей считается печально известным вредителем, ежегодно наносящим ущерб на миллиарды долларов и наносящим ущерб дикой природе. Муравьи процветают в городских районах, поэтому их присутствие может отпугнуть их от активного отдыха. Гнезда можно строить под такими конструкциями, как тротуары и фундаменты, что может вызвать структурные проблемы или привести к их разрушению. Они могут не только повредить или разрушить конструкции, но и завезенные красным огненные муравьи могут повредить оборудование и инфраструктуру и повлиять на бизнес, землю и стоимость имущества. В сельском хозяйстве они могут повредить урожай и технику, а также угрожать пастбищам. Известно, что они вторгаются в самые разные культуры, а насыпи, построенные на сельскохозяйственных угодьях, могут помешать уборке урожая. Они также представляют угрозу для животных и домашнего скота, способные нанести серьезные травмы или убить их, особенно молодых, слабых или больных животных. Несмотря на это, они могут быть полезными, поскольку поедают обычных насекомых-вредителей сельскохозяйственных культур. Обычные методы борьбы с этими муравьями включают травлю и окуривание; другие методы могут быть неэффективными или опасными. Из-за своей дурной славы и важности муравей стал одним из самых изученных насекомых на планете, даже соперничая с ним. западная медоносная пчела (Apis mellifera).[3][4]
Этимология и общие имена
В специфический эпитет красного импортного огненного муравья, invicta, происходит от латинский, и означает «непобедимый» или «непобедимый».[5][6][7] Эпитет происходит от фразы Рома Инвикта («Непокоренный Рим»), использовавшаяся как вдохновляющая цитата до падение Западной Римской Империи в 476 ОБЪЯВЛЕНИЕ. В общий имя, Соленопсис, переводится как "внешний вид" или "лицо" с Древнегреческий. Это сложный двух древнегреческих слов -солен, что означает «труба» или «канал», и опсис, что означает «внешний вид» или «взгляд».[8][9] Муравей широко известен как «красный импортированный огненный муравей» (сокращенно RIFA) из-за жжения, вызываемого его укусом.[10][11] Альтернативные названия включают: «огненный муравей», «красный муравей» или «бродячий муравей».[12][13] В Бразилии местные жители называют муравья Toicinhera, которое происходит от португальского слова Toicinho (свиной жир).[14]
Таксономия

Красный завезенный огненный муравей был впервые описан швейцарским энтомологом. Феликс Сански в журнальной статье 1916 г., опубликованной Physis.[15] Первоначально названный Solenopsis saevissima wagneri из синтип рабочий собран из Сантьяго-дель-Эстеро, Аргентина, Сански полагал, что муравей был разновидностью S. saevissima; специфический эпитет, Вагнери, происходит от фамилии Э.Р.Вагнера, собравшего первые образцы.[16] В тип материал в настоящее время находится в Натуристорический музей Базеля, Швейцария, но дополнительные рабочие, возможно, размещаются в Национальный музей естественной истории, Париж.[17] В 1930 году американская мирмеколог Уильям Крейтон рассмотрел род Соленопсис и реклассифицировал таксон как Solenopsis saevissima electra wagneri в инфраподвидовый звание, отметив, что он не смог собрать рабочих, которые ссылались бы на первоначальное описание Сански.[18] В 1952 г. S. saevissima комплекс видов был изучен и вместе с девятью другими названиями видовой группы, S. saevissima electra wagneri был синонимом S. saevissima saevissima.[19] Эта реклассификация была принята австралийским энтомологом Джорджем Эттершанком в его пересмотре рода и в каталоге неотропических муравьев Уолтера Кемпфа 1972 года.[20][21]
В 1972 году американский энтомолог Уильям Бурен описал то, что он считал новым видом, назвав его Solenopsis invicta.[22] Бурен собрал голотип рабочий из Куяба в Mato Grosso, Бразилия, и предоставил первое официальное описание муравья в журнальной статье, опубликованной Энтомологическим обществом Джорджии. Он случайно ошибся в написании invicta в качестве invica над страницами описания вида, хотя было ясно, что invicta было предполагаемое написание из-за постоянного использования имени в статье.[23] Типовой материал в настоящее время размещен в Национальный музей естественной истории, Вашингтон, округ Колумбия.[22]

В обзоре комплекса видов в 1991 году американский энтомолог Джеймс Трагер синонимизированный S. saevissima electra wagneri и С. вагнери вместе.[23] Трагер неправильно цитирует Соленопсис saevissima electra wagneri как исходное имя, ошибочно полагая, что имя С. вагнери был недоступен и использовал имя Бурена S. invicta. Ранее Трагер считал, что S. invicta был схож с S. saevissima до сравнения материала с С. вагнери. Трагер отмечает, что С. вагнери имеет приоритет над S. invicta, имя никогда не использовалось выше инфраподвидового ранга. Использование названия с момента Сански не связано с собранными образцами, и в результате nomen nudum.[23] В 1995 г. английский мирмеколог Барри Болтон исправил ошибку Трагера, распознав С. вагнери как действительное имя и синоним S. invicta.[24] Он заявляет, что Трагер ошибочно классифицировал С. вагнери как недоступное имя и цитирует S. saevissima electra wagneri как исходный таксон. Он заключает, что С. вагнери фактически является исходным именем и имеет приоритет над S. invicta.[24][25]
В 1999 году Стив Шаттак и его коллеги предложили сохранить название S. invicta.[17] Поскольку первое описание S. invictaбыло опубликовано более 1800 научных работ с использованием этого имени, в которых обсуждается широкий круг тем, касающихся его экологического поведения, генетики, химической коммуникации, экономических последствий, методов контроля, населения и физиологии. Они заявляют, что использование С. вагнери является «угрозой» номенклатурной стабильности для ученых и не ученых; систематики, возможно, смогли адаптироваться к такому изменению названия, но в таком случае может возникнуть путаница в названии. В связи с этим Шаттак и его коллеги предложили продолжить использование S. invicta и нет С. вагнери, поскольку это имя использовалось редко; с 1995 по 1998 год было опубликовано более 100 статей с использованием S. invicta и только трое используют С. вагнери. Они просили, чтобы Международная комиссия по зоологической номенклатуре (ICZN) использовать полномочия подавить С. вагнери в целях Принципа приоритета, а не для Принцип омонимии. Кроме того, они просили, чтобы имя S. invicta быть добавленным в Официальный список конкретных названий в зоологии и что С. вагнери быть добавленным в Официальный указатель отклоненных недействительных конкретных названий в зоологии.[17] После рассмотрения предложение было проголосовано энтомологическим сообществом и было поддержано всеми, кроме одного избирателя. Они отмечают, что нет оправдания подавлению С. вагнери; вместо этого было бы лучше отдать предпочтение S. invicta над С. вагнери всякий раз, когда автор рассматривал их как сородичи. МКЗН сохранит S. invicta и подавить С. вагнери в обзоре 2001 года.[26] В соответствии с настоящей классификацией красный завезенный огненный муравей является представителем рода Соленопсис в племени Соленопсидини, подсемейство Myrmicinae. Это член семьи Formicidae, принадлежащих к порядку Перепончатокрылые, отряд насекомых, содержащих муравьев, пчелы, и осы.[27]


Филогения
Красный импортированный огненный муравей является членом S. saevissima видовая группа. Членов можно отличить по их двучленикам на конце жгутика у рабочих и маток, а второй и третий сегменты жгутика вдвое длиннее и шире у более крупных рабочих. Полиморфизм встречается у всех видов и челюсти иметь четыре зуба.[22] Следующее кладограмма показывает положение завезенного красного огненного муравья среди других членов S. saevissima видовая группа:[а][28]
| Соленопсис |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Фенотипические и генетические данные позволяют предположить, что красный завезенный огненный муравей и черный импортный огненный муравей (Соленопсис Richteri) отличаются друг от друга, но имеют близкое генетическое родство.[29][30][31][32] Гибридизация Между двумя муравьями встречается в областях, где они контактируют, с гибридной зоной, расположенной в Миссисипи. Такая гибридизация возникла в результате вторичного контакта между этими двумя муравьями несколько десятилетий назад, когда они впервые столкнулись друг с другом на юге. Алабама.[29][33] На основе митохондриальная ДНК, осмотрел гаплотипы не образуют монофилетический клады. Некоторые из исследованных гаплотипов образуют более близкое родство с С. мегергейтс, С. quinquecuspis и С. Рихтери чем они делают с другими S. invicta гаплотипы. Возникновение возможного парафилетический группировка предполагает, что красный импортный огненный муравей и С. quinquecuspis Возможные группы скрытых видов состоят из нескольких видов, которые не могут быть различимы морфологически.[32][34]
Генетика
Исследования показывают, что митохондриальная ДНК вариации происходят существенно в полигиния общества (гнезда с несколькими матками),[35] но никаких изменений не обнаружено в моногиния общества (гнезда с одиночной маткой).[36] Триплоидия (хромосомная аномалия) часто встречается у красных завезенных огненных муравьев (до 12% у непродуктивных самок), что связано с высокой частотой диплоидных самцов.[37] Красный импортированный огненный муравей - первый вид, обладающий ген зеленой бороды, благодаря которому естественный отбор может способствовать альтруистическое поведение. Рабочие, у которых есть этот ген, могут различать маток, содержащих его, и маток, у которых нет, по-видимому, по запахам. Рабочие убивают маток, не содержащих гена.[38][39] В 2011 году ученые заявили, что полностью последовательный красный импортный огненный муравей геном от мужчины.[40]
Описание

Красные импортные рабочие-пожарные муравьи различаются по размеру от маленького до среднего, что делает их полиморфный. Рабочие измеряют от 2,4 до 6,0 мм (от 0,094 до 0,236 дюйма).[41] Головка имеет размеры от 0,66 до 1,41 мм (от 0,026 до 0,056 дюйма) и от 0,65 до 1,43 мм (от 0,026 до 0,056 дюйма) в ширину. У более крупных рабочих (как и у крупных рабочих) их головы имеют ширину от 1,35 до 1,40 мм (от 0,053 до 0,055 дюйма) и от 1,39 до 1,42 мм (от 0,055 до 0,056 дюйма). Антенна Scapes от 0,96 до 1,02 мм (от 0,038 до 0,040 дюйма), а грудной длина от 1,70 до 1,73 мм (от 0,067 до 0,068 дюйма).[22] Голова за глазами становится шире с округлыми затылочные доли присутствует, и в отличие от похожих на вид S. richteriпик долей находится дальше средней линии, но затылочное иссечение не такое складчатое. Стебли у крупных рабочих не выходят за затылочную вершину на один или два диаметра скапуса; эта особенность более заметна в С. Рихтери. У средних рабочих черешки достигают затылочных вершин и превышают заднюю границу у самых мелких рабочих. У мелких и средних рабочих голова имеет более эллиптические стороны. Голова мелких рабочих спереди шире, чем сзади.[22] У основных рабочих переднеспинка у него нет угловатых плеч и нет утопленной заднемедиальной области. Промезонотум выпуклый, а проподеум основание округлое, а также выпуклое. Основание и склон равны по длине. В шов промезонотума у более крупных рабочих либо сильный, либо слабый. В черешок имеет толстую и тупую чешуйку; если смотреть сзади, он не такой округлый сверху, как S. richteri, а иногда он может быть частично усеченным. Рыхлиток большой и широкий, у более крупных рабочих шире своей длины. Постпетиоле может быть менее широким спереди и более широким сзади. На тыльной стороне дорсальной поверхности имеется поперечное вдавление. В С. Рихтери, эта особенность тоже присутствует, но намного слабее.[22]
Скульптура очень похожа на С. Рихтери.[22] В пунктах возникает волосистость, и они часто имеют удлиненную форму на дорсальной и вентральной частях головы. На грудной клетке растяжки присутствуют, но они менее выгравированы с меньшим количеством проколов, чем на груди. С. Рихтери. На черешке точки расположены по бокам. Послевкусие при осмотре сверху имеет сильную шагрень с отчетливыми поперечными пунктировками. Бока покрыты глубокими точками, которые кажутся меньше, но глубже. В С. Рихтери, проколы крупнее и неглубокие. Это придает поверхности более непрозрачный вид. В некоторых случаях вокруг задней части могут присутствовать точки.[22] Волосность похожа на С. Рихтери. Эти волосы прямостоячие и различаются по длине, кажутся длинными с каждой стороны переднеспинка и мезонотум; на голове длинные волосы видны продольными рядами. Многочисленные прижатые опушенные волоски на черешковой чешуе; это наоборот в С. Рихтери, поскольку эти волоски редкие.[22] Рабочие выглядят красными и желтоватыми с коричневым или полностью черным. желудок.[15] Желудочные пятна иногда наблюдаются у более крупных рабочих, где они не так ярко окрашены, как у рабочих. S. richteri. Пятно желудка обычно покрывает небольшую часть первого желудочного пятна. тергит. Грудь однотонная, от светло-красновато-коричневого до темно-коричневого. Ноги и тазики обычно слегка заштрихованы. У крупных рабочих голова имеет однородный цветной рисунок, с затылок и вершина кажется коричневым. Другие части головы, включая переднюю часть, щеки и центральную часть наличник, желтоватые или желтовато-коричневые. Передние границы щек и мандибул темно-коричневые; они оба имеют одинаковый оттенок с затылком. Пейзажи и фуникулеры могут быть одного цвета с головой или одного оттенка с затылком. Светлые участки головы у рабочих малого и среднего размера ограничиваются только лобной областью с присутствием темного пятна, напоминающего стрелу или ракету. Иногда гнезда могут быть разных цветов. Например, рабочие могут быть намного темнее, а пятно в желудке может полностью отсутствовать или иметь темно-коричневый цвет.[22]
Королевы имеют длину головы от 1,27 до 1,29 мм (от 0,050 до 0,051 дюйма) и ширину от 1,32 до 1,33 мм (от 0,052 до 0,052 дюйма).[22] Плечи имеют размер от 0,95 до 0,98 мм (от 0,037 до 0,039 дюйма), а грудная клетка - от 2,60 до 2,63 мм (от 0,102 до 0,104 дюйма). Голову почти не отличить от С. Рихтери, но затылочное иссечение менее складчатое, а гребни значительно короче. Стеблевидная чешуя выпуклая, напоминает чешую. С. Рихтери. У раструба прямые стороны, которые никогда не вогнуты, в отличие от С. Рихтери где они вогнуты. Грудная клетка практически идентична, но между метаплевральной полосатой зоной и проподеальной зоной остается свободное пространство. дыхальца либо узкая складка, либо ее нет. Боковые части черешка пунктированы. Стороны раструба непрозрачный с наличием проколов, но не наблюдается нерегулярной шероховатости. Передняя часть спинки шагрень, а в средней и задней областях - поперечные точечные бороздки. Во всех этих регионах есть прямостоячие волоски. Передние части черешка и раструба имеют прижатое опушение, которое также заметно на проподеуме. По цвету царица такая же, как у рабочего: брюшко темно-коричневое, а ноги, черепа и грудная клетка светло-коричневые с темными полосами на спине. мезоскутум. Голова желтоватая или желтовато-коричневая вокруг центральных областей, затылок и нижние челюсти имеют цвет грудной клетки, а жилки крыльев варьируются от бесцветных до бледно-коричневых.[22] Мужчины выглядят похожими на S. richteri, но верхняя граница черешковых чешуек более вогнутая. У обоих видов дыхальца раструба и черешка сильно выступают. Все тело самца однотонно-черного цвета, но усики беловатые. Как и у королевы, жилки крыльев бесцветные или бледно-коричневые.[22]

Красный импортированный муравей может быть ошибочно идентифицирован как похожий на С. Рихтери.[22] Эти два вида можно отличить друг от друга с помощью морфологического исследования головы, грудной клетки и раструба. В S. richteriстороны головы широко эллиптические, а сердцевидная форма, наблюдаемая у красного импортного огненного муравья, отсутствует. Область затылочных долей, которая расположена рядом со средней линией и иссечением затылочной части, кажется более похожей на складку. С. Рихтери чем у красного импортного огненного муравья. Пейзажи С. Рихтери длиннее, чем у красного импортного огненного муравья, а переднеспинка имеет сильные угловатые плечи. Такой характер почти отсутствует у красного импортного огненного муравья. Мелководный, но затонувший участок известен только крупным рабочим С. Рихтери, который расположен в задней области спина переднеспинки. Эта особенность полностью отсутствует у более крупных красных привозных пожарных муравьев. Промезонотум красного завезенного муравья сильно выпуклый, тогда как у S. richteri. При осмотре основание проподеума удлиненное и прямое в С. Рихтери, а у красного импортного огненного муравья выпуклый и короче. У него также есть широкий раструб с прямыми или расходящимися сторонами. Постпетиооле в С. Рихтери уже со сходящимися сторонами. В С. Рихтерипоперечное вдавление на постеродорсальной части раструба сильное, но слабое или отсутствует у красного завезенного огненного муравья.[22] Кроме того, S. richteri рабочие на 15% крупнее, чем привозные красные рабочие-пожарные муравьи, имеют черно-коричневый цвет и имеют желтую полосу на спинной стороне брюшка.[42][43]
Выводок

Яйца крошечные и имеют овальную форму, примерно неделю остаются неизменными. Через неделю яйцо принимает форму зародыша и превращается в личинку при удалении скорлупы.[44] Личинки имеют размер 3 мм (0,12 дюйма).[45] Они похожи на S. geminata личинки, но их можно отличить по покровам с шипики поверх дорсальной части задних сомитов. Волосы на теле имеют размер от 0,063 до 0,113 мм (от 0,0025 до 0,0044 дюйма) с зубчатым кончиком. Усики у обеих по две или три сенсилла. В верхняя губа меньше с двумя волосками на передней поверхности 0,013 мм (0,00051 дюйма). В верхняя челюсть между кардо и ножками имеется склеротизированная полоса. В губа также имеет небольшую склеротизованную повязку.[45] Трубки губные железы Известно, что они производят или секретируют белковое вещество, которое имеет богатый уровень пищеварительных ферментов, в том числе протеазы и амилазы, которые действуют как внекишечное переваривание твердой пищи. Средняя кишка также содержит амилазы, ротейсы и упасы. Узкие клетки в его резервуаре практически не имеют функции секреции.[46] Куколки напоминают взрослых особей любой касты, за исключением того, что их ноги и усики плотно прижаты к телу. Они кажутся белыми, но со временем куколки темнеют, когда они почти готовы к созреванию.[47]
Четыре личинки возрастов были описаны на основе отличительных морфологических признаков.[44][48][49] Личинок малых и крупных рабочих невозможно отличить до последнего возраста, когда различия в размерах становятся очевидными.[49] При окукливании более заметная разница в ширине головы между кастами. Репродуктивные личинки крупнее личинок рабочих и имеют дискретные морфологические различия в ротовом аппарате.[49] Личинок четвертого возраста самцов и маток можно отличить по относительной форме и окраске тела.[49] а также внутренние имагинальные гоноподальные диски могут различаться.
Полиморфизм
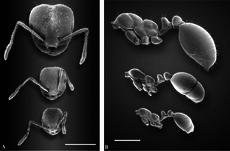
Красный импортированный огненный муравей полиморфен с двумя разными кастами рабочих: мелкими рабочими и крупными рабочими (солдатами). Подобно многим муравьям, проявляющим полиморфизм, молодые, более мелкие муравьи не кормятся и вместо этого стремятся к расплоду, в то время как более крупные рабочие уходят за кормом.[50][51][52][53] В зарождающихся колониях полиморфизма не существует, но вместо этого они заняты мономорфными рабочими, называемыми «минимами» или «нанитиками». Средняя ширина головы в тестируемых колониях увеличивается в течение первых шести месяцев развития.[54][55] В пятилетних колониях ширина головы мелких рабочих уменьшается, но у крупных рабочих ширина головы остается прежней. Общий вес крупного рабочего вдвое превышает вес второстепенного рабочего, когда они впервые приезжают, и к шести месяцам основные рабочие в четыре раза тяжелее второстепенных рабочих. Когда основные рабочие развиваются, они могут составлять значительную часть рабочей силы, причем до 35% составляют основные рабочие в одной колонии.[54] Это не влияет на производительность колонии, поскольку полиморфные колонии и гнезда с небольшими рабочими производят выводки примерно с одинаковой скоростью, а полиморфизм не является преимуществом или недостатком, когда источники пищи не ограничены. Однако полиморфные колонии более энергетически эффективны, и в условиях, когда пища ограничена, полиморфизм может дать небольшое преимущество в производстве расплода, но это зависит от уровней пищевого стресса.[56]
По мере того как рабочие муравьи вырастают до больших размеров, форма головы изменяется из-за того, что длина головы увеличивается одновременно с общей длиной тела, а ширина головы может увеличиваться на 20%. Длина усиков растет медленно; усики могут вырасти только на 60% к тому времени, когда длина тела удвоится, таким образом, относительная длина усиков уменьшается на 20% по мере того, как длина тела удваивается.[55] Все отдельные ноги тела изометричны с длиной тела, что означает, что даже когда длина тела удваивается, ноги также удваиваются. Однако не все ноги одинаковой длины; переднегрудная часть составляет 29% длины ноги, среднегрудная - 31%, а заднегрудная - 41%. Первые две пары ног одинаковой длины, а последняя пара длиннее.[55] В целом морфологический облик рабочего кардинально меняется, когда он вырастает. Голова показывает наибольшее изменение формы и высоту алинотум растет быстрее, чем его длина, где соотношение рост / длина составляет 0,27 у мелких рабочих и 0,32 у крупных рабочих.[55] Из-за этого более крупные рабочие, как правило, имеют горбатую форму и крепкий алинотум в отличие от более мелких рабочих.Ни один из сегментов черешка не изменяет форму при изменении размеров тела. Ширина брюшной полости растет быстрее, чем ее длина, где ширина может составлять 96% от ее длины, но увеличивается до 106%.[55]
Физиология
Как и другие насекомые, красный импортный огненный муравей дышит через систему наполненных газом трубок, называемых трахеи связаны с внешней средой через дыхальца. Терминальные ветви трахеи (трахеолы ) непосредственно контактируют с внутренними органами и тканями. Транспортировка кислорода к клеткам (и углекислого газа из клеток) происходит через распространение газов между трахеолами и окружающей тканью, чему способствует прерывистый газообмен.[57] Как и в случае с другими насекомыми, прямое сообщение между системой трахеи и тканями устраняет необходимость в сети циркулирующей жидкости для транспортировки O2.[58] Таким образом, красные импортированные огненные муравьи и другие членистоногие могут иметь скромную систему кровообращения, хотя у них очень дорогие метаболические потребности.[59]
В выделительная система состоит из трех регионов. Базальная область состоит из трех клеток, находящихся в задней части средней кишки. Передняя и верхняя полости образованы основаниями четырех Мальпигиевы канальцы.[60] Верхняя полость открывается в просвет тонкой кишки. Прямая кишка представляет собой большой, но тонкостенный мешок, занимающий заднюю пятую часть личинок. Выброс отходов контролируется ректальные клапаны которые ведут к анусу.[60] Иногда личинки выделяют жидкость, состоящую из мочевая кислота, вода и соли.[61] Это содержимое часто выносится рабочими на улицу и выбрасывается, но колонии в условиях водного стресса могут поглотить это содержимое.[60] в репродуктивная система, королевы выпускают феромон что предотвращает сделки и оогенез у девственных самок; те, что тестируются в колониях без матки, начинают ооцит развиваются после деформации и начинают откладывать яйца.[62] Дегенерация летных мышц инициируется брачными и ювенильными гормонами и предотвращается с помощью аллатэктомия тела.[63][64] Гистолиз начинается с роспуска миофибрилла и медленный распад миофиламенты. Такое растворение продолжается до тех пор, пока не будут достигнуты единственные свободные материалы Z-линии, которые также исчезнут; только ядра и пластинчатые тела оставаться.[60] В одном исследовании аминокислоты увеличение гемолимфа после осеменения.[65]
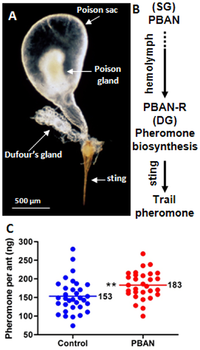
В железистая система содержит четыре железы: нижнечелюстную, верхнечелюстную, губную и постглоточный железы.[60] Заглоточная часть хорошо развита у матки, тогда как остальные железы больше у рабочих. Постглоточная железа функционирует как вакуум для поглощения жирные кислоты и триглицериды, а также желудочный слепая кишка.[66] Функции других желез остаются плохо изученными. В одном исследовании, посвященном ферментам пищеварительной системы взрослых муравьев, липаза активность была обнаружена в нижнечелюстных и губных железах, а также инвертаза Мероприятия. В Железа Дюфура найденный в муравье действует как источник след феромонов, хотя ученые полагали, что ядовитая железа была источником феромона королевы.[60][67][68] Нейрогормон нейропептид, активирующий биосинтез феромонов содержится в муравье, которое активирует биосинтез феромонов железы Дюфура.[69] В сперматека у кошек есть железа, которая выполняет функцию поддержания спермы. У самцов эти железы отсутствуют, но те, которые связаны с его головой, морфологически схожи с таковыми у рабочих, но эти железы могут действовать по-другому.[66]

Муравей сталкивается со многими респираторными проблемами из-за очень изменчивой окружающей среды, которая может вызвать увеличение высыхание, гипоксия, и гиперкапния. Жаркий влажный климат вызывает учащение пульса и дыхания, что увеличивает потерю энергии и воды.[58][70] Гипоксия и гиперкапния могут возникнуть в результате того, что колонии завезенных красных огненных муравьев живут в плохо вентилируемых помещениях. терморегулирующий курганы и подземные гнезда. Прерывистый газообмен (ДГЭ) может позволить муравьям выжить в гиперкапнических и гипоксических условиях, часто встречающихся в их норах;[57] он идеально подходит для адаптации к этим условиям, поскольку позволяет муравьям увеличивать период O2 потребление и CO2 изгнание самостоятельно через манипуляции с дыхательными путями. Успех вторжения красного завезенного огненного муравья может быть связан с его физиологической устойчивостью к абиотический стресс, будучи более теплостойкими и более адаптируемыми к стрессу высыхания, чем S. richteri. Это означает, что муравей менее уязвим к жаре и высыханию. Несмотря на то что S. richteri имеет более высокое содержание воды, чем красный импортный огненный муравей, S. richteri был более уязвим к стрессу высыхания. Меньшая чувствительность к высыханию обусловлена меньшей потерей воды.[71] Колонии, живущие в незатененных и более теплых местах, как правило, более устойчивы к жаре, чем те, которые живут в затененных и более прохладных местах.[72]
Скорость обмена веществ, косвенно влияющая на дыхание, также зависит от температуры окружающей среды. Вершина горы метаболизм происходит примерно при 32 ° C.[73] Метаболизм и, следовательно, частота дыхания постоянно увеличивается с повышением температуры. DGE останавливается при температуре выше 25 ° C, хотя причина этого в настоящее время неизвестна.[74]
На частоту дыхания также оказывает значительное влияние каста. У самцов частота дыхания значительно выше, чем у самок и рабочих, отчасти из-за их способности к полету и большей мышечной массы. В целом у мужчин больше мускулов и меньше жира, что приводит к более высокому уровню метаболизма O2 требовать.[74]Хотя скорость метаболизма наиболее высока при 32 ° C, колонии часто процветают при немного более низких температурах (около 25 ° C). Высокая скорость метаболизма, связанная с более высокими температурами, является ограничивающим фактором для роста колоний, потому что потребность в потреблении пищи также увеличивается. В результате более крупные колонии, как правило, обнаруживаются в более прохладных условиях, поскольку метаболические потребности, необходимые для поддержания колонии, снижаются.[73]
Распространение и среда обитания
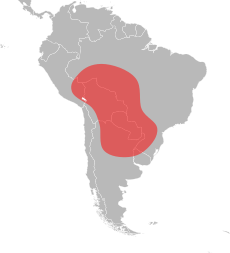
Импортированные красные огненные муравьи являются родными для тропических районов Центральной и Южной Америки, где их географический ареал простирается от юго-востока Перу до центральной Аргентины и на юге Бразилии.[75][76][77][78] В отличие от его географического ареала в Северной Америке, его ареал в Южной Америке значительно отличается. У него очень длинный диапазон с севера на юг, но очень узкий с востока на запад. Самая северная находка красного завезенного огненного муравья - Порту-Велью в Бразилии, а его самый южный рекорд - Resistencia в Аргентине; это расстояние около 3000 км (1900 миль). Для сравнения, ширина его узкого хребта составляет около 350 км (220 миль), и это, скорее всего, уже в южной части Аргентины и Парагвая и в северных районах бассейна реки Амазонки.[79] Наиболее известные записи о завезенных красных огненных муравьях находятся в Пантанальский район Бразилии. Однако внутренняя часть этой области не была тщательно исследована, но очевидно, что этот вид обитает в благоприятных местах вокруг нее. Считается, что регион Пантанал является родиной завезенного красного огненного муравья; рассеяние гидрохора через плавучие муравьиные плоты, можно легко объяснить популяцию далекого юга вокруг Парагвай и Реки Гуапоре. Западная протяженность его ареала точно не известна, но его численность там может быть ограничена. Он может быть обширным в восточной части Боливии из-за присутствия региона Пантанал.[79]
Эти муравьи происходят из Аргентины, и красный импортированный огненный муравей, скорее всего, пришел именно отсюда, когда они впервые вторглись в Соединенные Штаты; в частности, популяции этих муравьев были обнаружены в провинциях Чако, Корриентес, Formosa, Сантьяго-дель-Эстеро, Санта-Фе, и Тукуман.[23][79][80] Северо-восточные районы Аргентины являются наиболее вероятным предположением, откуда пришли муравьи-вторжения.[77] В Бразилии они встречаются в северной части Мату-Гросу и в Рондония И в Штат Сан-Паулу. Красный импортный огненный муравей и S. saevissima находятся парапатрический в Бразилии, с контактными зонами, известными в Мату-Гросу-ду-Сул, Штат Парана и Сан-Паулу.[76][81] В Парагвае они встречаются по всей стране и были зарегистрированы в Бокерон, Каагуасу, Canindeyú, Центральная, Гуайра, Eembucú, Парагуари, и Presidente Hayes отделы; Трагер утверждает, что муравей распространен во всех регионах страны.[82][83][84] Они также встречаются на большей части северо-востока Боливии и, в меньшей степени, на северо-западе Уругвая.[79][85]

Красный завезенный огненный муравей может доминировать в измененных областях и жить в различных средах обитания. Он может выжить в экстремальных погодных условиях тропических лесов Южной Америки, а в неспокойных районах часто можно увидеть гнезда рядом с дорогами и зданиями.[86][87] Муравья часто наблюдали вокруг поймы реки Парагвай.[88] В местах, где присутствует вода, они обычно встречаются вокруг: оросительных каналов, озер, прудов, водохранилищ, рек, ручьев, берегов рек и мангровых болот.[82][86] Гнезда встречаются в сельскохозяйственных районах, на прибрежных территориях, в заболоченных местах, на остатках прибрежных дюн, в пустынях, лесах, лугах, естественных лесах, дубовых лесах, мезический лес, опавшие листья, окраины пляжа, кустарники, вдоль железных и автомобильных дорог, а также в городских районах.[89] В частности, они встречаются на обрабатываемых землях, управляемых лесах и плантациях, нарушенных территориях, в системах интенсивного животноводства и теплицах.[86][90] Было обнаружено, что завезенные красные огненные муравьи вторгаются в здания, в том числе в медицинские учреждения.[91] В городских условиях колонии обитают на открытых участках, особенно если местность солнечная.[91] Сюда входят: городские сады, площадки для пикников, газоны, детские площадки, школьные дворы, парки и поля для гольфа.[82][91] В некоторых районах в среднем 200 насыпей на акр.[92] Зимой колонии перемещаются под тротуары или в здания, а недавно повязанные матки переходят на пастбища.[86][91] Красные привозные огненные муравьи в основном встречаются на высоте от 5 до 145 м (от 16 до 476 футов) над уровнем моря.[82]
Насыпи варьируются от маленьких до больших, от 10 до 60 см (от 3,9 до 23,6 дюйма) в высоту и 46 см (18 дюймов) в диаметре, без видимых входов.[86][93] Рабочие могут получить доступ к своим гнездам только через серию туннелей, которые выходят из центральной области. Такие выступы могут простираться на расстояние до 25 футов от центрального холма либо прямо вниз к земле, либо, чаще, сбоку от исходного холма.[94] Построенные из почвы, насыпи ориентированы так, что длинные части насыпи обращены к солнцу ранним утром и перед закатом.[86][93] Курганы обычно имеют овальную форму с длинной осью гнезда, ориентированной с севера на юг.[95] Эти муравьи также тратят большое количество энергии на строительство гнезда и транспортировку расплода, что связано с терморегуляция. Расплод вывозят в районы с высокими температурами; рабочие отслеживают температурные режимы насыпи и не полагаются на поведенческие привычки.[96] Внутри гнезд насыпи содержат серию узких горизонтальных туннелей с подземными валами и узлами, доходящими до корней травы на 10–20 см (от 3,9 до 7,9 дюйма) ниже поверхности; эти шахты и узлы соединяют туннели насыпей с подземными камерами. Эти камеры около 5 см.2 (0,77 дюйма2) и достигают глубины от 10 до 80 см (от 3,9 до 31,5 дюйма). Среднее количество муравьев в одной подземной камере составляет около 200.[97][98][99]
Введение

Красные привозные огненные муравьи относятся к числу наихудший инвазивные виды в мире.[100][101] Некоторые ученые считают завезенного красного огненного муравья «специалистом по беспокойствам»; нарушение человеком окружающей среды может быть основным фактором воздействия муравьев (огненные муравьи, как правило, предпочитают нарушенные участки). Это показано в одном эксперименте, демонстрирующем, что покос и вспашка на изучаемых территориях уменьшили разнообразие и численность местных видов муравьев, в то время как красные завезенные огненные муравьи, обнаруженные на нетронутых лесных участках, уменьшили только пару видов.[102][103]
В США красный завезенный огненный муравей впервые прибыл в морской порт Мобил, Алабама на грузовом судне с 1933 по 1945 гг.[b][c][79][106][107][108] Прибытие примерно с 9-20 неродственными матками,[109][110] красный завезенный огненный муравей в то время был лишь редкостью, поскольку энтомологи не могли собрать никаких экземпляров (первые наблюдения были сделаны в 1942 г., а в 1937 г. им предшествовал рост популяции); популяция этих муравьев резко увеличилась к 1950-м годам.[111][112][113] С момента своего завоза в США красный завезенный огненный муравей распространился по южным штатам и северо-востоку Мексики, оказывая негативное воздействие на дикую природу и нанося экономический ущерб.[75][114][115][116] Распространение красных импортных огненных муравьев может быть ограничено, так как они почти полностью уничтожены во время Теннесси зимы, таким образом, они могут достичь своего самого северного ареала.[117][118][119] Однако глобальное потепление может позволить завезенным красным огненным муравьям расширить свой географический ареал.[120] По данным на 2004 год, муравей встречается в 13 штатах и занимает более 128 миллионов гектаров земли, а на одном акре земли можно найти до 400 курганов.[121][122] В Министерство сельского хозяйства США по оценкам, они расширяются на 193 км (120 миль) в год.[91]
Красные привозные огненные муравьи были впервые обнаружены в Квинсленд, Австралия, 2001 г.[123][124] Предполагалось, что муравьи присутствовали в морских контейнерах, прибывающих в Порт Брисбена, скорее всего, из Северной Америки.[125] Неофициальные данные свидетельствуют о том, что огненные муравьи могли присутствовать в Австралии за шесть-восемь лет до официальной идентификации. Возможный ущерб от завезенного красного огненного муравья побудил правительство Австралии быстро отреагировать. Совместное государственное и федеральное финансирование Австралийский доллар 175 миллионов было выделено на шестилетнюю программу ликвидации.[126][127][128] Сообщалось, что после нескольких лет искоренения уровень уничтожения превысил 99% на ранее зараженных участках. Программа получила расширенный Содружество финансирование в размере около 10 миллионов австралийских долларов в течение как минимум двух лет для лечения остаточных инвазий, обнаруженных совсем недавно.[129] В декабре 2014 г. гнездо было обнаружено на Port Botany, Сидней, в Новый Южный Уэльс. Порт был помещен на карантин, проведена операция по удалению.[130] В сентябре 2015 года жители США были обнаружены в аэропорту Брисбена.[131]

Красные привозные огненные муравьи распространились за пределы Северной Америки. В Группа специалистов по инвазивным видам (ISSG) сообщает о муравьях, населяющих три из Каймановы острова. Однако источники, на которые ссылается ISSG, не сообщают о них на острове, но недавние коллекции указывают на их присутствие.[75] В 2001 году в Новой Зеландии были обнаружены завезенные красные огненные муравьи, но через несколько лет они были успешно уничтожены.[75][132] В Индии зарегистрированы завезенные красные огненные муравьи.[133] Малайзия,[134] Филиппины[135] и Сингапур.[75] Однако эти отчеты оказались неверными, поскольку собранные там муравьи были ошибочно идентифицированы как красный импортированный огненный муравей. В Сингапуре муравьи, скорее всего, тоже были идентифицированы неправильно. В Индии обследованные муравьи в Саттур Талук, Индия, перечислили завезенных красных огненных муравьев там в больших популяциях; Между тем, за пределами обследованной территории сообщений о муравье не поступало. В 2016 году ученые заявили, что, несмотря на отсутствие муравья в Индии, завезенный красный огненный муравей, скорее всего, найдет подходящие места обитания в экосистеме Индии, если ему будет предоставлена возможность.[136] В сообщениях с Филиппин, скорее всего, неправильно идентифицировали собранный материал как красного импортированного огненного муравья, так как популяции там не обнаружены.[75] Это было обнаружено в Гонконг и материковый Китай в 2004 г., где они распространились на несколько провинций, а также Макао и Тайвань.[137][138][139][140] Никакие географические или климатические барьеры не препятствуют дальнейшему распространению этих муравьев, поэтому они могут распространяться по тропическим и субтропическим регионам Азии.[121][141] В Европе единственное гнездо было найдено в Нидерландах в 2002 году.[78]
Примерно в 1980 г. красные завезенные огненные муравьи начали распространяться по Вест-Индии, где о них впервые сообщили в Пуэрто-Рико и Виргинские острова США.[142][143] В период с 1991 по 2001 год муравей был зарегистрирован в Тринидаде и Тобаго, нескольких районах Багамских островов, Британских Виргинских островах, Антигуа и островах Теркс и Кайкос.[75][144][145] С тех пор красные завезенные огненные муравьи были зарегистрированы на большем количестве островов и регионов, а новые популяции были обнаружены в: Ангилья, Сен-Мартен, Барбуда, Монсеррат, Сент-Китс, Невис, Аруба, и Ямайка.[75] Муравьи, зарегистрированные на Арубе и Ямайке, были обнаружены только на полях для гольфа; эти курсы импортируют дерьмо из Флорида, поэтому такой ввоз может быть важным способом распространения муравья по Вест-Индии.[75]
Популяции, обитающие за пределами Северной Америки, происходят из Соединенных Штатов. В 2011 году ДНК образцов из Австралии, Китая и Тайваня была проанализирована, и результаты показали, что они родственны образцам из Соединенных Штатов.[146] Несмотря на свое распространение, S. geminata имеет больший географический ареал, чем красный завезенный огненный муравей, но может быть легко вытеснен им. Из-за этого почти весь его экзотический ареал в Северной Америке был утерян, и муравей там почти исчез. На обочинах дорог во Флориде 83% этих сайтов S. geminata присутствует, когда красный завезенный огненный муравей отсутствовал, но только 7%, когда он присутствует.[75] Это означает, что муравей, вероятно, может вторгнуться во многие тропические и субтропические регионы, где S. geminata популяции присутствуют.[75]
Поведение и экология

Импортированные красные огненные муравьи чрезвычайно устойчивы и адаптировались к условиям наводнения и засухи. Если муравьи ощущают повышенный уровень воды в своих гнездах, они соединяются вместе и образуют плавучий шар или плот с рабочими снаружи и царицей внутри.[147][148][149] Выводок переносится на самую высокую поверхность.[150] Они также используются как основание плота, за исключением яиц и более мелких личинок. Перед тем, как погрузиться в воду, муравьи нырнут в воду и разорвут связь с сушей. В некоторых случаях рабочие могут намеренно снять с плота всех самцов, в результате чего самцы утонут. Продолжительность жизни плота может достигать 12 дней. Муравьи, пойманные в ловушку под водой, спасаются, поднимаясь на поверхность с помощью пузырьков, которые собираются с погруженного в воду субстрата.[150] Из-за большей уязвимости для хищников, завезенные красные огненные муравьи значительно агрессивнее во время сплава. Рабочие обычно доставляют более высокие дозы яда, что снижает опасность нападения других животных. Из-за этого, а также из-за того, что доступно большее количество муравьев, плоты потенциально опасны для тех, кто с ними сталкивается.[151]
Некрофорное поведение встречается у красного импортного огненного муравья. Рабочие выбрасывают несъеденные продукты и другие подобные отходы подальше от гнезда. Активный компонент не был идентифицирован, но были задействованы жирные кислоты, накапливающиеся в результате разложения, и кусочки бумаги, покрытые синтетическим олеиновая кислота обычно вызывало некрофорную реакцию. Процесс, лежащий в основе такого поведения у импортированных красных огненных муравьев, был подтвержден Блюмом (1970): ненасыщенные жиры, такие как олеиновая кислота, вызывают поведение при удалении трупов.[152] Рабочие также по-разному реагируют на мертвых рабочих и куколок. Мертвых рабочих обычно выносят из гнезда, тогда как у куколок может пройти день, прежде чем произойдет некрофорная реакция. Куколки инфицированы Metarhizium anisopliae обычно выбрасываются рабочими по более высокой ставке; 47,5% непораженных трупов выбрасываются в течение суток, а для пораженных трупов этот показатель составляет 73,8%.[153]
Красные привозные огненные муравьи негативно влияют на семена прорастание. Однако степень повреждения зависит от того, как долго семена остаются уязвимыми (засыхают и прорастают), а также от численности муравьев.[154] Одно исследование показало, что в то время как эти муравьи привлекаются и удаляют семена, которые адаптировались для распространения муравьев, завезенные красные огненные муравьи повреждают эти семена или перемещают их в неблагоприятные для прорастания места. В семенах, переданных в семьи, 80% Сангвинария канадская семена были скарифицированы и 86% Виола круглолистная семена были уничтожены.[155] Небольшие проценты длиннолистная сосна (Сосна болотная) семена, депонированные рабочими, успешно прорастают, что свидетельствует о том, что красные завезенные огненные муравьи помогают перемещению семян в экосистеме длиннолистной сосны. Семена, несущие элайосомы собираются с большей скоростью, чем семена, не содержащие элайосомы, и хранят их не в гнездах, а в поверхностных кучах мусора в окрестностях насыпи.[156]
Собирательство и общение

Колонии завезенных красных огненных муравьев имеют поверхности туннелей, которые выступают из поверхностей, на которых рабочие кормятся.[157][158] Эти области выступа обычно находятся на их собственной территории, но большая колонизация муравьев может повлиять на это.[159] Туннели предназначены для эффективного взаимодействия тела, конечностей и антенн со стенами, и рабочий также может перемещаться внутри них исключительно быстро (более девяти длин тела в секунду).[160] Норы выходят из любой точки на территории колонии, и работникам, собирающим пищу, может потребоваться полметра, чтобы добраться до поверхности. Если предположить, что средний собиратель проходит 5 м, более 90% времени на поиск корма приходится на туннели днем и редко - ночью. Рабочие собирают корм при температуре почвы до 27 ° C (80 ° F) и температуре поверхности 12–51 ° C (53–123 ° F).[158] Рабочие, подвергающиеся воздействию температуры 42 ° C (107 ° F), рискуют умереть от жары.[158] К осени количество рабочих, добывающих пищу, быстро снижается, а зимой они появляются редко. Это может быть связано с влиянием температуры почвы и уменьшением предпочтения источников пищи. Эти предпочтения уменьшаются только при низкой продуктивности расплода. В северных регионах США районы слишком холодные, чтобы муравьи могли кормиться, но в других регионах, таких как Флорида и Техас, кормление может происходить круглый год. Когда идет дождь, рабочие не собирают корм на улице, так как выходные отверстия временно заблокированы, следы феромонов размываются, а сборщики могут быть физически поражены дождем. Влажность почвы также может влиять на поведение рабочих при поиске пищи.[158]
Когда рабочие собирают пищу, для него характерно три этапа: поиск, вербовка и транспортировка.[161] Рабочие обычно ищут мед чаще, чем другие источники пищи, и вес пищи не влияет на время поиска. Рабочие могут нанять других сокамерников, если пища, которую они нашли, слишком тяжелая, и для прибытия максимального числа нанятых рабочих требуется до 30 минут. Более легкие источники пищи занимают меньше времени и обычно быстро транспортируются.[161] Собиратели становятся разведчиками и ищут пищу исключительно за пределами поверхности, а через две недели могут умереть от старости.[162]
Рабочие общаются серией полухимикаты и феромоны. Эти методы общения используются в различных видах деятельности, таких как вербовка сородичей, поиск пищи, привлечение и защита; например, рабочий может секретировать след феромонов если обнаруженный источник пищи слишком велик, чтобы его нести.[163] Эти феромоны синтезируются Железа Дюфура и может идти от обнаруженного источника пищи обратно в гнездо.[164][165] Компоненты в этих следовых феромонах также видоспецифичны только для этого муравья, в отличие от других муравьев с обычными феромонами на хвосте.[166] Ядовитый мешок у этого вида был идентифицирован как новое место хранения феромона королевы; Известно, что этот феромон вызывает ориентацию у рабочих особей, что приводит к отложению расплода.[68][167] Это также аттрактант, когда рабочие собираются в области, где высвобождается феромон.[163] Возможно, присутствует феромон выводка, так как рабочие могут разделять выводок по возрасту и касте, после чего следует вылизывание, уход и антенна.[163] Если колония подвергается атаке, рабочие выделяют феромоны тревоги.[168] Однако у рабочих эти феромоны развиты слабо. Рабочие могут обнаружить пиразины которые производятся крылатками; эти пиразины могут участвовать в брачный полет, а также срабатывание сигнализации.[169]
Красные импортированные огненные муравьи могут различать своих товарищей по гнезду и других посредством химической связи и специфических запахов колонии.[163][170] Рабочие предпочитают копать в гнездах материалы из своей собственной колонии, а не из почвы в незагнанных районах или из других импортированных красных колоний огненных муравьев. Одно исследование предполагает, что, поскольку диета колонии похожа, единственной разницей между гнездовой и негнездовой почвой было гнездование самих муравьев. Следовательно, рабочие могут переносить запах колоний в почву.[170] Запах колонии может зависеть от окружающей среды, поскольку рабочие в колониях, выращиваемых в лаборатории, менее агрессивны, чем в дикой природе.[171] Реплики, производные от королевы, могут регулировать распознавание сородичей у рабочих и амин уровни. Однако эти сигналы не играют важной роли в распознавании на уровне колонии, но они могут служить формой распознавания касты в гнездах.[172][173] Рабочие, живущие в моногиния общества, как правило, очень агрессивны и нападают на злоумышленников из соседних гнезд. В колониях без маток добавление чужеродных маток или рабочих не увеличивает агрессию среди населения.[174]
Рацион питания

Красные привозные огненные муравьи всеядные, а фуражиры считаются мусорщики а не хищников.[175] Рацион муравьев состоит из мертвых млекопитающих, членистоногие,[176] насекомые, дождевые черви, позвоночные и твердые пищевые продукты, такие как семена. Однако этот вид предпочитает жидкую пищу твердой. Жидкая пища, которую собирают муравьи, - это сладкие вещества из растений или нектар -производство двукрылые.[93][175][177][178] Добыча членистоногих может включать: двукрылое животное взрослые особи, личинки и куколки, и термиты. Известно, что потребление сахарной аминокислоты влияет на набор рабочих для выращивания нектаров. У мимических растений с сахаром редко есть рабочие, которые могут ими питаться, тогда как у растений с сахаром и аминокислотами их много.[179][180] Среда обитания, в которой они живут, может определять, какой корм они собирают больше всего; например, показатели успешности получения твердой пищи наиболее высоки на прибрежных участках озер, в то время как высокие уровни жидких источников были собраны на пастбищах.[181] Определенные диеты также могут повлиять на рост колонии, при этом лабораторные колонии демонстрируют высокий рост, если их кормить медовой водой.Колонии, которые питаются насекомыми и сахарной водой, могут вырасти исключительно большими за короткий период времени, тогда как те, которые не питаются сахарной водой, растут значительно медленнее. Колонии, которые не питаются насекомыми, полностью прекращают производство расплода.[178] В целом, в колониях регулируется объем перевариваемой соками пищи.[182] Личинки способны проявлять независимые аппетиты к источникам, таким как твердые белки, растворы аминокислот и растворы сахарозы, и они также предпочитают эти источники разбавленным растворам. Такое поведение связано с их способностью сообщать рабочим о голоде. Скорость потребления зависит от типа, концентрации и состояния пищи, которой они питаются. Рабочие, как правило, привлекают больше сокамерников к источникам пищи, богатым сахарозой, чем к источникам белка.[182]
Распределение еды играет важную роль в колонии. Это поведение различается в разных колониях: мелкие рабочие получают больше еды, чем более крупные, если небольшая колония серьезно лишена пищи. Однако в более крупных колониях более крупные рабочие получают больше еды. Рабочие могут эффективно жертвовать сахарную воду другим сокамерникам, причем некоторые из них действуют как доноры. Эти «доноры» раздают свои источники пищи получателям, которые также могут выступать в качестве доноров. Рабочие также могут делиться большей частью своей еды с другими товарищами по гнезду.[183] В колониях, не переживающих голод, еда по-прежнему распределяется между рабочими и личинками. Одно исследование показывает, что мед и соевое масло скармливались личинкам через 12–24 часа после того, как они оставались рабочими. Соотношение распределения этих источников пищи составляло 40% по отношению к личинкам и 60% по отношению к рабочему для меда, а для соевого масла этот показатель составлял около 30 и 70% соответственно.[184] Импортированные красные огненные муравьи также запасаются определенными источниками пищи, такими как кусочки насекомых, вместо того, чтобы сразу же потреблять их. Эти куски обычно переносятся ниже поверхности насыпи в самые сухие и теплые места.[185]
Этот вид занимается трофалаксис с личинками.[186] Независимо от характеристик и состояния каждой личинки, их кормят примерно одинаковым количеством жидкой пищи. Скорость трофалаксиса может увеличиваться с личиночным лишением пищи, но такое увеличение зависит от размера каждой личинки. Личинки, которых кормят регулярно, обычно получают небольшое количество. Чтобы достичь насыщения, всем личинкам, независимо от их размера, обычно требуется восемь часов кормления.[187]
Хищники

Ряд насекомых, паукообразные, и птицы охотятся на этих муравьев, особенно когда королевы пытаются основать новую колонию.[188] В то время как в отсутствие защищающихся рабочих королевы огненных муравьев должны полагаться на свой яд, чтобы отпугнуть виды-конкуренты. [2] Многие виды стрекоза, включая Анакс юниус, Pachydiplax longipennis, Соматохлора провокаторы, и Tramea carolina, поймайте маток, пока они в полете; 16 видов пауков, в том числе паук-волк Lycosa timuga и южный паук черная вдова (Latrodectus mactans), активно убивают красных привозных огненных муравьев. L. mactans захватывает все касты вида (рабочих, королев и самцов) в своей сети. Эти муравьи составляют 75% добычи, пойманной пауком. Несовершеннолетний L. mactans также были замечены пауки, ловящие муравьев.[188][189] Другими беспозвоночными, которые охотятся на красных импортированных огненных муравьев, являются уховертки (Лабидура рипария ) и тигровых жуков (Цицинделла точечная ).[188] Птицы, которые едят этих муравьев, включают дымоход стриж (Chaetura pelagica), восточная королевская птица (Tyrannus tyrannus), а восточный бобуайт (Colinus virginianus virginianus). Восточный бобуайт атакует этих муравьев, выкапывая холмы в поисках молодых королев.[188] Красные привозные огненные муравьи были обнаружены в содержимом желудка внутри броненосцы.[190]
Было замечено, что многие виды муравьев нападают на королев и убивают их. По-видимому, яд королев огненных муравьев химически адаптирован для быстрого подавления обидных муравьев-конкурентов.[2] К хищным муравьям относятся: Ectatomma edentatum, Эфебомирмекс виды, Ласиус неонигер, Фейдол виды, Погономирмекс бадиус, и Conomyrma insana, который является одним из самых значимых.[188][191][192] C. insana Известно, что муравьи являются эффективными хищниками против королев-основателей в изученных районах Северной Флориды. Напор атак, инициированных C. insana со временем увеличиваются, заставляя королев проявлять различные реакции, включая побег, укрытие или защиту. Большинство маток, на которых нападают эти муравьи, в конечном итоге погибают.[191] Королевы, которые находятся в группах, имеют более высокие шансы на выживание, чем одиночные королевы, если на них нападают S. geminata.[193] Муравьи могут атаковать цариц на земле и вторгаться в гнезда, жаля и расчленяя их. Другие муравьи, такие как P. porcula попробуйте взять голову и желудок, и К. Клара вторгаются группами. Кроме того, некоторые муравьи пытаются вытащить маток из их гнезд, дергая за усики или ноги.[192] Маленькие мономорфные муравьи полагаются на рекрутов, чтобы убить королев, и не нападают на них, пока не прибудет подкрепление. Помимо убийства королевы, некоторые муравьи могут украсть яйца для потребления или испустить репеллент, который эффективен против импортированных красных огненных муравьев.[192] Некоторые виды муравьев могут совершать набеги на колонии и уничтожать их.[194]
Паразиты, патогены и вирусы

Мухи в роду Псевдактеон (форидные мухи) паразитируют на муравьях. Некоторые виды этого рода, например Псевдактеон трехстворчатый, были введены в окружающую среду с целью борьбы с завезенным огненным муравьем. Эти мухи паразитоиды красного завезенного огненного муравья в его естественном ареале в Южной Америке, и его можно привлечь через алкалоиды яда муравьев.[195] Один вид, Pseudacteon obtusus, нападает на муравья, приземляясь на заднюю часть головы и откладывая яйцо. Расположение яйца не позволяет муравью успешно удалить его.[196] Личинки мигрируют к голове, а затем развиваются, питаясь гемолимфа, мышечная ткань и нервная ткань. Примерно через две недели они заставляют муравья упасть, выпустив фермент который растворяет мембрану, прикрепляющую голову к телу. Муха окукливается в отделившейся головной капсуле, появляющейся через две недели.[196][197] П. трикуспис Другой форидный муха, паразитирующая для этого вида. Хотя паразитизм со стороны этих мух не влияет на плотность и активность популяции муравьев, он оказывает небольшое влияние на популяцию колонии.[198] В Strepsipteran насекомое Caenocholax fenyesi известно, что они заражают самцов муравьев этого вида и атакуют яйца,[199][200] и клещ Pyemotes tritici считается потенциальным биологическим агентом против завезенных красных огненных муравьев, способных паразитировать на каждой касте в колонии.[201] Бактерии, такие как Вольбахия, был обнаружен у красного импортного огненного муравья; Известно, что красный импортированный огненный муравей заражает три различных варианта бактерий. Однако его влияние на муравья неизвестно.[202][203] Solenopsis daguerrei является репродуктивным паразитом для колоний красных импортированных огненных муравьев.[204]
Большое количество болезнетворных микроорганизмов и нематод также заражают завезенных красных огненных муравьев. Возбудители включают Myrmecomyces annellisae, Mattesia виды, Steinernema виды,[205] а мермитид нематода,[206] Vairimorpha invictae, который может передаваться через живых личинок и куколок, а также через мертвых взрослых особей.[207][208] и Тетрадонема соленопсис, который может быть фатальным для большой части колонии.[209] Лица, инфицированные Metarhizium anisopliae имеют тенденцию выполнять трофалаксис чаще и имеют повышенное предпочтение хинин, алкалоидное вещество.[210] Форид летает с Kneallhazia solenopsae может служить векторов в передаче болезни муравьям.[211] Ослабляя колонию, инфекции, вызванные этим заболеванием, локализуются в жировой ткани тела, а споры возникают только у взрослых людей.[212][213] Смертность инфицированной колонии обычно выше, чем в здоровой.[206] Эти муравьи являются хозяином Конидиобол,[214] Myrmicinosporidium durum, и Боверия бассиана, каждый из которых является паразитическим грибком. У инфицированных людей споры покрывают все тело, и они выглядят темнее, чем обычно.[215][216][217] Токсичность из-за антимикробных свойств летучие вещества произведенные муравьями, могут значительно снизить скорость прорастания Б. bassiana внутри колонии.[218]
Вирус, S. invicta 1 (SINV-1 ), был обнаружен примерно в 20% полей огненных муравьев, где, по-видимому, вызывает медленную гибель инфицированных колоний. Он оказался самоподдерживающимся и передаваемым. После внедрения он может уничтожить колонию в течение трех месяцев. Исследователи считают, что вирус потенциально может стать жизнеспособным биопестицид управлять огненными муравьями.[219][220][221] Также были обнаружены еще два вируса: S. invicta 2 (SINV-2) и S. invicta 3 (SINV-3). Полигинный колонии, как правило, сталкиваются с более серьезными инфекциями по сравнению с моногинный колонии. Также могут возникать множественные вирусные инфекции.[222][223]
Жизненный цикл и воспроизводство

Брачный полет у красных завозных огненных муравьев начинается в более теплое время года (весна и лето), обычно через два дня после дождя. Время крылатые выходить и мат - с полудня до 3:00 вечера.[224][225] В свадебных полетах, зарегистрированных в Северной Флориде, в среднем участвуют 690 женщин и мужчин в одном полете.[226] Самцы первыми покидают гнездо, и оба пола охотно отправляются в полет, практически не предпринимая никаких предполетных действий. Однако рабочие роются в холмике, возбужденно стимулируемые нижнечелюстными железами в голове крылатых крыльев.[227][228] Поскольку в насыпях нет отверстий, рабочие формируют отверстия во время брачного полета, чтобы выплеснуть крылья. Такое поведение у рабочих, вызванное феромонами, включает быстрый бег, возвратно-поступательные движения и повышенную агрессию. Рабочие также группируются вокруг крыльев, когда они взбираются на растительность, и в некоторых случаях пытаются опустить их обратно, прежде чем они взлетят. Химические сигналы самцов и самок во время брачного полета привлекают рабочих, но химические сигналы, посылаемые рабочими, не привлекают других сокамерников. Это также вызывает тревожное поведение рабочих, что приводит к более высокому уровню поиска крылатых стволов.[227][229]
Самцы летают на большей высоте, чем самки: пойманные самцы обычно находятся на высоте от 100 до 300 м (от 330 до 980 футов) над поверхностью, а самки - только от 60 до 120 м (от 200 до 390 футов) над поверхностью. Брачный полет длится примерно полчаса, а самки обычно пролетают менее 1,6 км (0,99 мили) до приземления. Около 95% маток успешно спариваются и спариваются только один раз; некоторые мужчины могут быть бесплодны из-за того, что доли яичек не развиваются.[224][230][231][232][233][234] В полигинных колониях самцы не играют существенной роли, и поэтому большинство из них бесплодны; одна из причин этого - избегать спаривания с другими видами муравьев. Это также делает селективную смертность самцов, что может повлиять на систему размножения, успех спаривания и, поток генов.[235][236] Идеальные условия для начала брачного полета - это когда уровень влажности выше 80% и температура почвы выше 18 ° C (64 ° F). Брачные полеты происходят только при температуре окружающей среды 24–32 ° C (75–89 ° F).[224]
Королевы часто встречаются в 1–2,3 милях от гнезда, из которого они прилетели. Основание колонии может осуществляться индивидуально или группами, известными как плеометроз.[237] Эти совместные усилия соучредительниц способствуют росту и выживанию зарождающейся колонии; Гнезда, основанные несколькими матками, начинают период роста с в три раза большим количеством рабочих по сравнению с колониями, основанными одной маткой. Несмотря на это, такие ассоциации не всегда устойчивы.[238][239][240][241] Появление первых рабочих разжигает борьбу королева-королева и королева-рабочая. В плеометротических условиях только одна королева выходит победителем, тогда как проигравшие королевы впоследствии убиваются рабочими.[242] Двумя факторами, которые могут повлиять на выживание отдельных маток, являются их относительные боевые способности и их относительный вклад в производство рабочих. Размер, показатель боеспособности, положительно коррелирует с выживаемостью. Однако манипулирование относительным вкладом королевы в производство рабочих не коррелировало с выживаемостью.[243]
Одна матка откладывает от 10 до 15 яиц через 24 часа после спаривания.[93] В установленных гнездах королева наносит яд на каждое яйцо, которое, возможно, содержит сигнал, призывающий рабочих переместить его.[244] Эти яйца остаются неизменными в размере в течение одной недели, пока из них не вылупятся личинки. К этому времени матка отложит еще от 75 до 125 яиц. Личинки, которые вылупляются из яиц, обычно на несколько дней покрыты оболочкой скорлупы. Личинки могут освободить ротовой аппарат от панциря с помощью движений тела, но при вылуплении все еще нуждаются в помощи рабочих. Личиночная стадия делится на четыре возрастов, что наблюдается на стадии линьки. В конце каждой линьки виден кусок неизвестного материала, соединенный с exuviae если они изолированы от рабочих. Личиночная стадия длится от шести до 12 дней, прежде чем их тела значительно расширятся и станут куколками; стадия куколки длится от 9 до 16 дней.[44][93]
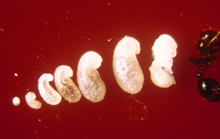
Как только первые особи достигают стадии куколки, матка прекращает яйценоскость до тех пор, пока не созреют первые рабочие. Этот процесс занимает от двух недель до одного месяца. Молодых личинок кормят маслами, которые срыгивают из ее урожая, а также трофические яйца или выделения. Она также кормит детенышей своими мышцами крыльев, обеспечивая их необходимыми питательными веществами. Первое поколение рабочих всегда мало из-за ограниченного количества питательных веществ, необходимых для развития. Эти рабочие, известные как минимумы или нанитики, вырываются из покоев королевы и начинают добывать пищу, необходимую для колонии. Строительство кургана также происходит в это время. В течение месяца после рождения первого поколения начинают развиваться более крупные рабочие (основные рабочие), и в течение шести месяцев курган будет заметен, если его осмотреть, и в нем проживает несколько тысяч жителей. Зрелая матка способна откладывать 1500 яиц в день; все рабочие бесплодны, поэтому не могут воспроизводить потомство.[93][245][246][247]
Колония может расти исключительно быстро. Колонии, в которых в мае работало 15–20 человек, к сентябрю выросли до более 7000 человек. Эти колонии начали производить репродуктивных муравьев, когда им был год, и к тому времени, когда им исполнилось два года, в них было более 25 000 рабочих. Население увеличилось вдвое до 50 000, когда этим колониям исполнилось три года.[248] В зрелом возрасте колония может содержать от 100 000 до 250 000 особей, но другие отчеты предполагают, что колонии могут содержать более 400 000 особей.[d][249][250][251][252] Колонии полигинов могут расти намного больше, чем колонии моногинов.[250]
Несколько факторов способствуют росту колонии. Температура играет важную роль в росте и развитии колонии; рост колонии прекращается ниже 24 ° C, а время развития сокращается с 55 дней при температуре 24 ° C до 23 дней при 35 ° C. Рост установленных колоний происходит только при температуре от 24 до 36 ° C. Нанитовый выводок также развивается намного быстрее, чем мелкий рабочий выводок (примерно на 35% быстрее), что полезно для основания колоний.[253] Известно, что колонии, имеющие доступ к неограниченному количеству кормовых насекомых, значительно увеличиваются, но этот рост еще больше ускоряется, если они могут получить доступ к растительным ресурсам, колонизированным двукрылое животное насекомые.[254] В зарождающихся моногинных колониях, где производятся диплоидные самцы, уровень смертности колоний значительно высок, а рост колоний медленный. В некоторых случаях моногинные колонии подвержены 100% смертности на ранних стадиях развития.[255]

Продолжительность жизни рабочего муравья зависит от его размера, хотя в среднем составляет около 62 дней.[256] Ожидается, что второстепенные рабочие проживут от 30 до 60 дней, тогда как более крупные рабочие живут намного дольше. Более крупные рабочие, продолжительность жизни которых составляет от 60 до 180 дней, живут на 50–140% дольше, чем их коллеги меньшего размера, но это зависит от температуры.[93][257] Однако известно, что рабочие, содержащиеся в лабораторных условиях, живут от 10 до 70 недель (от 70 до 490 дней); максимальная зарегистрированная продолжительность жизни рабочего составляет 97 недель (или 679 дней).[258] Королевы живут намного дольше рабочих - от двух до почти семи лет.[93][258]
В колониях королевы - единственные муравьи, которые могут изменять соотношение полов, которое можно предсказать. Например, королевы, происходящие из колоний, производящих самцов, имеют тенденцию производить преимущественно самцов, в то время как королевы, происходящие из колоний с благоприятным соотношением полов, производят самок.[259][260] Королевы также осуществляют контроль над производством полов через феромоны которые влияют на поведение рабочих как по отношению к личинкам мужского, так и женского пола.[261]
Моногиния и многоженство
В красном импортированном огненном муравье есть две формы общества: полигинные колонии и моногинные колонии.[262][263][262] Полигинные колонии существенно отличаются от моногинных колоний по социальные насекомые. Предыдущее сокращение опыта в королеве плодовитость, расселение, долголетие и родство с соседями.[264][265][266][267] Полигинные королевы также меньше физиогастральный чем моногинные королевы и рабочие мельче.[268][269] Понимание механизмов вербовки маток является неотъемлемой частью понимания того, как эти различия в фитнес сформированы. Необычно то, что количество взрослых маток в колонии не влияет на набор новых маток. Уровни феромона королевы, который, по-видимому, связан с числом королевы, играют важную роль в регуляции воспроизводства. Из этого следовало, что рабочие будут отказываться от новых маток при воздействии большого количества этого феромона королевы. Более того, данные подтверждают утверждение о том, что королевы обеих популяций заходят в гнезда случайным образом, без учета количества присутствующих более старых маток.[270] Нет никакой корреляции между количеством старых маток и количеством вновь набранных маток. Были выдвинуты три гипотезы, объясняющие принятие нескольких маток в основанные колонии: мутуализм, родственный отбор, и паразитизм.[271] Гипотеза мутуализма гласит, что сотрудничество приводит к повышению личной пригодности пожилых королев. Однако эта гипотеза не согласуется с тем фактом, что увеличение числа маток снижает как продуктивность матки, так и продолжительность ее жизни.[272] Родственный отбор также кажется маловероятным, учитывая, что королевы наблюдали за сотрудничеством в обстоятельствах, когда они статистически не связаны.[273] Следовательно, королевы не улучшают личную физическую форму, допуская новых маток в колонию. Паразитизм существовавших ранее гнезд, по-видимому, является лучшим объяснением многоженства. Одна из теорий состоит в том, что так много маток пытаются проникнуть в колонию, что рабочие сбиваются с толку и непреднамеренно позволяют нескольким маткам присоединиться к ней.[273]
Рабочие Monogyne убивают иностранных королев и агрессивно защищают свою территорию. Однако не все модели поведения универсальны, в первую очередь потому, что поведение рабочих зависит от экологического контекста, в котором они развиваются, а также от манипуляций со стороны рабочего. генотипы может вызвать изменение в поведении. Следовательно, поведение коренного населения может отличаться от поведения интродуцированного населения.[274] В исследовании по оценке агрессивного поведения рабочих моногинных и полигинных красных огненных муравьев путем изучения взаимодействия на нейтральных аренах и разработки надежных этограмма для быстрого различения моногиния и полигиния колонии красных завезенных огненных муравьев в поле,[275] Рабочие-моногиняне и полигинники различали сокамерников и иностранцев, о чем свидетельствует различное поведение - от терпимости до агрессии. Муравьи-моногины всегда нападали на чужеродных муравьев независимо, были ли они из моногинных или полигинных колоний, в то время как муравьи-полигины распознавали, но не нападали на чужеродных муравьев-полигиний, в основном проявляя позы, аналогичные поведению, принимаемому после нападения муравьев. Псевдактеон фориды. Враждебное поведение по сравнению с предупреждением во многом зависело от социальной структуры рабочих. Таким образом, поведение по отношению к иностранным рабочим было методом характеристики моногинных и полигинных колоний.[276] Большинство колоний на юго-востоке и юге центральной части США имеют тенденцию быть моногинными.[241]
Моногинная территориальная площадь импортированной красной колонии огненных муравьев и размер насыпи положительно коррелируют, что, в свою очередь, регулируется размером колонии (количеством и биомассой рабочих), расстоянием от соседних колоний, плотностью добычи и коллективной конкурентоспособностью колонии. способность. В отличие от этого, дискриминация соплеменников среди полигинных колоний менее выражена, поскольку рабочие терпят сородич муравьи чужды колонии, принимают других гетерозигота королевы и не защищают свою территорию агрессивно от сородичей-полигинников.[277] Эти колонии могут увеличить свою репродуктивную продуктивность в результате наличия большого количества маток и возможности эксплуатации больших территорий с помощью кооперативного набора и связанных между собой курганов.[278]
У красного импортированного огненного муравья присутствует социальная хромосома. Эта хромосома может различать социальную организацию колонии, несущей один из двух вариантов суперген (B и b) который содержит более 600 гены. Социальную хромосому часто сравнивают с половыми хромосомами, потому что они имеют схожие генетические особенности.[279] и они определяют фенотип колонии аналогичным образом. Например, колонии, несущие исключительно вариант B этой хромосомы, принимают одиночных маток BB, но колонии с вариантами B и b будут принимать только нескольких маток Bb.[280] Различия в другом гене также могут определять, будет ли в колонии одна или несколько маток.[281]
Отношения с другими организмами

Когда полигинные формы вторгаются в районы, где колонии еще не созданы, разнообразие местных членистоногие и позвоночные сильно снижается.[282] Это очевидно, поскольку популяции изоподы, клещи и клопы скарабеи значительно снизиться. Они также могут значительно изменить популяции многих семейств мух и жуков, в том числе: Каллифориды, Histeridae, Muscidae, Саркофагиды, Silphidae, и Staphylinidae. Несмотря на это, в одном обзоре было обнаружено, что некоторые насекомые могут не поражаться импортированными красными огненными муравьями; например, плотность равноногих уменьшается в районах, зараженных красными завезенными огненными муравьями, но сверчки рода Gryllus не затронуты. Бывают случаи, когда разнообразие определенных видов насекомых и членистоногих увеличивается в районах, где присутствуют завезенные красные огненные муравьи.[283][284][285] Красные завезенные огненные муравьи - важные хищники пещерных беспозвоночных, некоторые из которых находятся под угрозой исчезновения. Это включает в себя уборщики урожая, псевдоскорпионы, пауки, жужелицы и жуки-пселафиды. Наибольшее беспокойство вызывает не сам муравей, а наживка, используемая для его лечения, потому что это может оказаться фатальным. Деревянные улитки Stock Island (Orthalicus reses) вымерли в дикой природе; хищничество красных импортированных огненных муравьев считается основным фактором исчезновения улиток.[283] В целом, красные импортированные огненные муравьи предпочитают определенных членистоногих другим, хотя они нападают и убивают любого беспозвоночного, которое не может защитить себя или убежать.[286] Биоразнообразие членистоногих увеличивается после того, как популяции красных завезенных огненных муравьев будут сокращены или полностью уничтожены.[114][287]
Взаимодействие между завезенными красными огненными муравьями и млекопитающими редко документировалось. Тем не менее, случаи смерти пойманных в ловушку животных от завезенных красных огненных муравьев наблюдались.[283][288][289] Показатели смертности в восточный хлопок (Сильвилаг флориданский) молодняк составляет от 33 до 75% из-за завезенных красных огненных муравьев.[290] Считается, что красные привозные огненные муравьи оказывают сильное влияние на многих герпетофауна разновидность; ученые отметили сокращение населения в Кингснейк Флориды (Лампропелтис getula floridana), а также яйца и взрослые особи восточная ящерица (Sceloporus undulatus) и шестицилиндровый гонщик (Aspidoscelis sexlineata) являются источником пищи.[291][292][293] Из-за этого восточные ящерицы из забора приспособились к более длинным ногам и новому поведению, чтобы избежать красного завезенного огненного муравья.[294][295] Взрослый трехпалые коробчатые черепахи (Террапен каролина триунгуис), Хьюстонская жаба (Anaxyrus houstonensis) молодняк, и Американский аллигатор (Аллигатор миссисипиенсис) детеныши также подвергаются нападению и гибели этими муравьями.[283][296][297][298] Несмотря на эту в основном отрицательную связь, одно исследование показывает, что завезенные красные огненные муравьи способны влиять на передачу трансмиссивных болезней, регулируя популяцию клещей и изменяя динамику переносчиков и хозяев, тем самым снижая скорость передачи не только животным, но и людям. .[299]
Уровень смертности птиц хорошо изучен; были случаи, когда ни один молодняк не дожил до взрослой жизни в районах с высокой плотностью огненных муравьев. Многие птицы, в том числе Ласточки гнездятся на скале, утки, цапли, перепел, и крачки пострадали от завезенных красных огненных муравьев.[300] Наземные гнездящиеся птицы, особенно наименьшая крачка (Sterna antillarum), уязвимы для атак огненных муравьев.[301][298] Воздействие завезенных красных огненных муравьев на птиц, гнездящихся в колониях, особенно серьезно; водоплавающие птицы уровень смертности может составлять 100%, хотя этот фактор был ниже у птиц, откладывающихся раньше срока. Выживаемость выводка снижается в Американские скальные ласточки (Petrochelidon pyrrhonota), если они находятся в контакте с рабочими-добытчиками.[302] Певчая птица выживаемость гнезд снижается в районах, где присутствуют красные завезенные огненные муравьи, но выживаемость в белоглазый виреон (Vireo griseus) и черношапочный виреон (Вирео атрикапилла) количество гнезд увеличивается с 10% до 31% и с 7% до 13%, когда огненные муравьи отсутствуют или когда они не могут напасть на них.[303] Импортированные красные огненные муравьи могут косвенно способствовать низкой выживаемости расплода в Прерийный цыпленок Аттуотера.[304] Первоначально считалось, что муравьи связаны с сокращением количества зимующих птиц, таких как сорокопут (Ланиус Людовиканус), но более позднее исследование показало, что усилия по уничтожению муравьев с помощью пестицида Мирекс, который, как известно, имел токсические побочные эффекты, был во многом виноват.[305][306]
Красные привозные огненные муравьи - сильные конкуренты многих видов муравьев. Им удалось вытеснить множество местных муравьев, что привело к ряду экологических последствий.[307] Однако исследования показывают, что эти муравьи не всегда являются лучшими конкурентами, подавляющими местных муравьев. Нарушение среды обитания до их прибытия и ограничения пополнения - более вероятные причины подавления местных муравьев.[308] Между Tapinoma melanocephalum и Pheidole fervida красный завезенный огненный муравей сильнее обоих видов, но проявляет разный уровень агрессии. Например, они менее враждебны по отношению к T. melanocephalum в отличие от P. fervida. Показатели смертности в T. melanocephalum и П.Fervida при борьбе с красными привозными огненными муравьями - 31,8% и 49,9% соответственно. Однако уровень смертности среди завезенных красных рабочих-муравьев составляет всего 0,2–12%.[309] В импортный сумасшедший муравей (Nylanderia fulva) демонстрирует большее доминирование, чем красный импортированный огненный муравей, и, как известно, вытесняет их в местах обитания, где они встречаются друг с другом.[310] Более крупные колонии мостовых муравьев (Tetramorium caespitum ) могут уничтожить импортированные красные колонии огненных муравьев, что привело энтомологов к выводу, что этот конфликт между двумя видами может помочь препятствовать распространению завезенных красных огненных муравьев.[311][312] Люди, инфицированные SINV-1, могут быть убиты быстрее, чем здоровые Monomorium chinense. Это означает, что муравьи, инфицированные вирусом SINV-1, более слабые, чем их здоровые собратья, и, скорее всего, будут уничтожены М. chinense. Однако крупных рабочих, независимо от того, инфицированы они или нет, убивают редко.[313]
В районах, где он является родным, красный завезенный огненный муравей по-прежнему является доминирующим видом и сосуществует с 28 видами муравьев в галерея лес пробелы и десять видов в ксерофитный лесные луга, выигрывая в наиболее агрессивных взаимодействиях с другими муравьями. Однако некоторые муравьи могут быть совместно доминирующими в областях, где они сосуществуют, например, Аргентинский муравей, где они конкурируют симметрично.[314][315] Рабочие регулярно вступают в пищевую конкуренцию с другими муравьями и могут подавить эксплуатацию пищевых ресурсов полупереводными насекомыми, производящими падь (особенно из Фенакокк соленопсис ) от аборигенных муравьев; однако красные импортные огненные муравьи не могут уничтожить T. melanocephalum полностью, хотя они потребляют больше пищи. Вместо этого два муравья могут мирно сосуществовать и делить падь.[316][317] При встрече с соседними огненными муравьями рабочие могут взять на себя изображающий смерть поведения, чтобы избежать их с успехом. Однако такое поведение наблюдается только у молодых рабочих, поскольку пожилые работники либо бегут, либо сопротивляются, когда им угрожают.[318]
Как уже упоминалось, красные импортные огненные муравьи и аргентинские муравьи конкурируют друг с другом. Уровни смертности различаются в зависимости от сценария (т.е. коэффициенты смертности в условиях противостояния колоний ниже, чем в условиях противостояния в полевых условиях). Основные рабочие также могут выдержать большее количество травм, что увеличивает смертность аргентинских муравьев. Для того, чтобы колония аргентинских муравьев успешно уничтожила моногинную колонию из 160 000 рабочих, колонии потребуется 396 800 рабочих. Колония, уменьшившаяся в размерах из-за успешной обработки наживкой, подвержена нападению аргентинских муравьев. Муравьи могут играть жизненно важную роль в уничтожении ослабленных колоний огненных муравьев, а также могут быть важны в замедлении распространения этих огненных муравьев, особенно в сильно зараженных аргентинских муравьёвых районах.[319] Несмотря на это, популяции аргентинских муравьев на юго-востоке США сократились после завоза красного огненного муравья.[320]

Красные импортные огненные муравьи установили отношения с инвазивным мучнистый червец, Фенакокк соленопсис. Известно, что рост колонии увеличивается, если муравьи имеют доступ к ресурсам, производимым П. соленопсис, а плотность популяции этих мучнистых червецов значительно выше в районах, где есть завезенные красные огненные муравьи (распространение П. соленопсис в районах, где это не родной, объясняется наличием муравьев). Кроме того, ожидаемая продолжительность жизни и репродуктивная способность П. соленопсис оба увеличиваются. Рабочие могут даже перенести этих мучнистых червецов обратно в свое гнездо.[321] Хищничество П. соленопсис уменьшается из-за того, что работники по уходу препятствуют хищничеству и паразитизму естественных врагов. На мучнистых червецов охотятся божья коровка (Menochilus sexmaculatus ) и являются хозяевами двух паразитов (Энасиус бамбавалей и Ацерофагус кокосовый ), если муравьев нет, но это другое дело, если муравьи есть. На растениях с красными привозными огненными муравьями реже встречаются божьи коровки, а количество нимф-мумий значительно ниже.[321] Это ясно видно, поскольку ученые наблюдали, что хлопковая тля (Тля gossypii ) популяции, а также увеличилось количество яиц сторожевой совки в районах с присутствием красных завезенных огненных муравьев.[322][323][324] Красные привозные огненные муравьи установили взаимоотношения с другим мучнистым червецом (Dysmicoccus morrisoni ). Муравьи способствуют росту колоний Д. Моррисони с помощью защиты, прикрывая колонии мусором и собирая выделяемую ими росу.[325]
Токсикология
В токсикология красного импортного яда огненных муравьев относительно хорошо изучен. В яд важен для муравья, позволяя ему поймать добычу и защитить себя. Около 14 миллионов человек ежегодно ужаливают в Соединенных Штатах,[326] которые могут страдать от реакций от легкой экземы до анафилактического шока.[327] Наиболее частые реакции на укусы огненных муравьев - это ощущение жжения в месте укуса, за которым следует крапивница и образование пустул.[328] У некоторых жертв может развиться более серьезная аллергическая реакция, которая может быть опасной для жизни.[329][330]Яд в основном состоит из нерастворимых алкалоидов с небольшой водянистой фазой.[331] содержит около 46 белков[332] из которых четыре указаны как аллергены.[333][334]
Отношения с людьми
Как вредители

Из-за своей дурной славы и агрессивного поведения красные импортированные огненные муравьи считаются вредители. В Соединенных Штатах Управление по контролю за продуктами и лекарствами по оценкам, более 5 миллиардов долларов ежегодно тратится на лечение, повреждение и контроль зараженных территорий.[335][336] Только в Техасе красные импортированные огненные муравьи нанесли ущерб домашнему скоту, дикой природе и здоровью населения на сумму 300 миллионов долларов.[337] Приблизительно 36 долларов на одно американское домохозяйство, и более 250 миллионов долларов было потрачено на борьбу с завезенными красными огненными муравьями и их искоренение. Частные агентства ежегодно тратят от 25 до 40 миллионов долларов на пестициды.[93] Неудачное искоренение посевов в Австралии может ежегодно обходиться экономике в миллиарды долларов, и, согласно исследованию правительства Квинсленда, оценочная стоимость может достигнуть 43 миллиардов долларов в течение 30 лет.[338]
Красные привозные огненные муравьи процветают в городских районах, особенно на задних дворах, на полях для гольфа, в парках, зонах отдыха, на школьных площадках и на обочинах улиц, где их присутствие может сдерживать занятия на свежем воздухе. Если они входят в дом или собственность, они могут причинить вред домашним животным, если они находятся в клетке, загоне, привязаны или не могут сбежать.[336][339] Гнезда можно строить под тротуарами или даже под дорогами, а также под проездами, фундаментами, газонами, краями тротуаров, под плитами патио, в электрических коробках или возле линий электропередач. Колония может выкапывать огромное количество почвы, что приводит к структурным проблемам на проезжей части, тротуарах и стенах, а также может вызывать образование выбоин на дорогах.[93][336][340][341][342] Дополнительный урон насыпями может быть нанесен деревьям, дворовым растениям и трубам; некоторые конструкции также могут обрушиться. Колонии могут мигрировать в дома людей после сильного дождя, чтобы укрыться от насыщенной почвы.[47][93]
Мало того, что они процветают в городских районах, красные завезенные огненные муравьи могут повредить оборудование и инфраструктуру, а также повлиять на бизнес, землю и стоимость имущества.[339] Их также привлекает электричество; Электростимулированные рабочие выделяют ядовитые алкалоиды, феромоны тревоги и феромоны найма, что, в свою очередь, привлекает на место больше рабочих.[343] В результате красные привозные огненные муравьи могут уничтожить электрическое оборудование.[344] Это известно как магнетизм, где ученые определили внутренние магнитные материалы, которые могут играть роль в ориентационном поведении.[345][346] Известно, что они прорывают электрическую изоляцию, что приводит к повреждению электродвигателей, ирригационных линий, насосов, сигнальных ящиков, трансформаторов, телефонных станций и другого оборудования.[336][347] Колонии собираются около электрических полей и могут вызывать короткие замыкания или мешать работе переключателей и оборудования, такого как кондиционеры, компьютеры и водяные насосы. Также известно, что они заражают посадочные площадки аэропортов и светофоры.[347]
В сельском хозяйстве

Красный завезенный огненный муравей является серьезным сельскохозяйственным вредителем в тех районах, где он не является местным. Они способны повредить посевы, угрожают пастбищам и сады.[339] Сами насыпи могут разрушить сельскохозяйственное оборудование, такое как ирригационные системы, и повредить технику во время сбора урожая.[348] Известно, что муравьи вторгаются в посевы соевых бобов, вызывая снижение урожайности, и могут причинить 156 миллионов долларов убытков посевам сои на юго-востоке США.[93][349][350] Репортажи из Грузии и Северная Каролина утверждают, что урожай сои от 16,8 до 49,1 кг / га невозможно было собрать из-за помех от насыпей огненных муравьев; комбайны перескакивали через насыпи, препятствующие уборке урожая, а фермеры поднимали косилки на своих комбайнах, чтобы не задеть насыпи.[351][352][353] Кормление красных импортированных огненных муравьев может привести к значительному ущербу для многих других культур, включая бобы, капусту, цитрусовые, кукурузу, огурцы, баклажаны, окра, арахис, картофель, сорго, подсолнечник и сладкий картофель. Муравьи также мешают работе корневой системы растений и питаются молодняком.[93][354] Иногда колонии строят холмы вокруг или возле основания цитрусовых деревьев, откусывают новые побеги и питаются цветами или развивающимися фруктами. Цитрусовые деревья часто опоясанный или убили.[355]
Несмотря на статус вредителя и известность, красный импортированный огненный муравей может принести пользу. Муравей - эффективный хищник насекомых, поэтому он может служить биологическим агентом против других видов вредителей, особенно на полях сахарного тростника.[356] К насекомым-вредителям, которых убивает муравей, относятся: долгоносик (Anthonomus grandis) в посевах хлопка,[357][358][359] бурильные молотки сахарного тростника (Diatraea saccharalis) на полях сахарного тростника,[360][361][362] рог летит (Гематобия раздражает) в навозе,[363] бархатные гусеницы (Anticarsia gemmatalis) в соевых бобах и белокрылки которые встречаются в теплицах.[364][365] Ряд исследований показывает, что завезенные красные огненные муравьи не мешают и не нападают на насекомых, имеющих экономическое значение на хлопковых полях, в результате чего многие фермеры в юго-восточных регионах США считают импортированных красных огненных муравьев полезными.[366] Однако некоторые ученые предположили, что полезный статус завезенного красного огненного муравья трудно предсказать, если не учитывать географию, размер растения, время года, влажность почвы и использование инсектицидов. Эти факторы могут снизить эффективность завезенных красных огненных муравьев в качестве средств борьбы с вредителями. Еще одним фактором является то, что рабочие действуют без разбора и убивают полезных насекомых, таких как хищники, которые поедают роговых мух и других вредителей на пастбищах, а также хищников тлей и щитовок.[366][367][368] Они также снижают эффективность паразитических ос против видов вредителей, поедая личинок и куколок.[366][369]
Контроль
По сравнению с другими видами муравьев, такими как Anoplolepis gracilipes, которые быстро захватывают области, в которых они были заключены, красные импортированные огненные муравьи довольно легко контролировать.[370] Первые предложения по борьбе с муравьем появились в 1957 году, когда Конгресс США санкционировал программу искоренения с использованием федерального и государственного финансирования. Исследования муравья и его биологии продолжались после создания программы искоренения, и для их уничтожения использовались многие химические вещества. Однако ученые обнаружили, что эти инсектициды убивали местную фауну, а Агентство по охране окружающей среды впоследствии объявил их вне закона. Некоторые ученые даже сомневались, были ли муравьи вредителями или нет.[371][372] Сегодня маловероятно, что красный завезенный огненный муравей будет уничтожен в таких регионах, как США. Управлять популяциями можно должным образом, если использовать комплексный подход.[373] Некоторые ученые рассматривали возможность использования против него естественных врагов муравьев; Это включает в себя Kneallhazia solenopsae и Б. bassiana. Форидные мухи также рассматриваются как потенциальные биологические агенты, поскольку они могут снизить активность кормления завезенных красных огненных муравьев и повлиять на уровни популяции. Однако они не могут повлиять на скорость роста колонии.[374] Кроме того, паразитические муравьи, паразитические осы, клещи, другие патогены, нематоды и грибы считаются потенциальными биологическими агентами.[375] Другие предполагают, что популяцию можно поддерживать или сокращать, управляя несколькими экологическими факторами.[376]

Для борьбы с популяциями использовалось несколько приманок. Насыпи уничтожаются за считанные недели, если на них использовать наживку. Приманки считаются эффективными и простыми в использовании против импортированных красных огненных муравьев, по сравнению с промыванием, опудриванием или окуриванием. Их посыпают на холм, затем муравьи берут и съедают.[377][378] Определенные приманки, такие как приманки, регулирующие рост, и водные приманки, содержащие борную кислоту и сахарозу, приносят пользу местной фауне, и для уничтожения колонии обычно требуются низкие концентрации.[379][380] Другие приманки, используемые против красных импортных огненных муравьев, включают: Amdro, Подъем, гидраметилнон и Maxforce.[381]
Твердые и жидкие инсектициды приманки, если они неправильно применены в месте, могут быть перемещены через поток воды в непредусмотренные источники воды. Часто это происходит из-за неточечных источников, включая множество диффузных источников. По мере таяния снега и дождя, проходящего через землю, вода собирает неправильно нанесенные инсектициды и откладывает их в более крупных водоемах, реках, заболоченных территориях или просачивается в водоразделы.
Исследователи также экспериментировали с другими методами, такими как окуривание, инъекция насыпи, промывание насыпи и поверхностное напыление. Обливание насыпи - это заливка в насыпь больших объемов токсичной жидкости. Хотя это может повлиять на большую часть насыпи, возможно, она не достигнет королевы, что предотвратит разрушение колонии. Обработка поверхности пылью похожа на пропитку насыпи, за исключением того, что инсектициды наносятся поверх насыпи и впитываются в почву при намокании. Для инъекций в насыпь используют давление и вводят инсектициды в насыпь, но это может не повлиять на королеву. Насыпи можно эффективно удалить с помощью фумигации. Иногда применяется широковещательная обработка путем опрыскивания зараженных участков сельскохозяйственной техникой. Наконец, можно использовать точечную обработку инсектицидами путем просверливания и введения в насыпь остаточного инсектицида.[93][377]
Помимо химического контроля, другие методы, которые можно использовать против этих муравьев, включают механические и электрические устройства. Однако неизвестно, эффективны ли эти устройства. Защита от муравьев может быть эффективной против колоний, гнездящихся внутри зданий, за счет заделки и заделки трещин, что успешно подавляет популяцию за пределами стен. Домовладельцы использовали свои собственные методы для удаления насыпей, обливая их кипятком или поджигая их легковоспламеняющимися жидкостями. Хотя эти методы могут быть эффективными, они не рекомендуются, поскольку могут нанести вред человеку и окружающей среде.[93][377]
Примечания
- ^ Несмотря на то что S. geminata появляется на кладограмме, это не член видовой группы. Он включен только для того, чтобы служить «точкой крепления»; если быть точным, он показывает, где исследуемая кладограмма соединяется с Дерево жизни.[28]
- ^ Некоторые источники предполагают, что они впервые прибыли в 1929 году.[104]
- ^ Красные привозные огненные муравьи, скорее всего, достигли Жители Нового Орлеана в Луизиана, также, но наличие Аргентинские муравьи не давали муравьям селиться и распространяться.[105]
- ^ Население отдельной колонии может изменяться в зависимости от сезона.[249]
Рекомендации
Сноски
- ^ Джонсон, Н.Ф. (19 декабря 2007 г.). "Solenopsis invicta Бурен ". Сервер имен перепончатокрылых, версия 1.5. Колумбус, Огайо, США: Государственный университет Огайо. Архивировано из оригинал 20 сентября 2016 г.. Получено 1 апреля 2015.
- ^ а б c Фокс, Эдуардо Г.П .; У, Сяоцин; Ван, Лэй; Чен, Ли; Лу Юн-Юэ; Сюй, Ицзюань (февраль 2019 г.). «Изозоленопсин А из яда королевы быстро выводит из строя конкурентов огненных муравьев». Токсикон. 158: 77–83. Дои:10.1016 / j.toxicon.2018.11.428. PMID 30529381. S2CID 54481057.
- ^ Чинкель 2006, п. vii.
- ^ Табер 2000, п. 12.
- ^ Bowersock, G.W .; Браун, П. (1999). Поздняя античность: путеводитель по постклассическому миру (2-е изд.). Кембридж: Belknap Press издательства Гарвардского университета. п.284. ISBN 978-0-674-51173-6.
- ^ Starr, C .; Evers, C .; Старр, Л. (2010). Биология: концепции и приложения без физиологии (8-е изд.). Белмонт, Калифорния: обучение Cengage. п.431. ISBN 978-0-538-73925-2.
- ^ Льюис, П. (24 июля 1990 г.). "Могучие огненные муравьи маршируют с юга". Нью-Йорк Таймс. Получено 2 ноября 2016.
- ^ Департамент окружающей среды, воды, наследия и искусства. «Банк видов: Solenopsis invicta (Семейство Formicidae) ". Исследование биологических ресурсов Австралии: Справочник фауны Австралии. Канберра: Правительство Австралии. Архивировано из оригинал 21 сентября 2016 г.. Получено 8 апреля 2014.CS1 maint: несколько имен: список авторов (связь)
- ^ Чинкель 2006 С. 13–14.
- ^ Дрис, Б. (2002). «Медицинские проблемы и рекомендации по лечению красного завезенного огненного муравья» (PDF). Техасский импортный проект по исследованию и управлению огненных муравьев. Техасский университет A&M. Получено 23 августа 2016.
- ^ Бут, К .; Дхами, М. (2008). Красный импортный огненный муравей (Solenopsis invicta): Обзор литературы по определению возраста колонии. (PDF). Центр биоразнообразия и биобезопасности (Отчет). Министерство сырьевых отраслей: Биобезопасность Новой Зеландии. п. 2. Архивировано из оригинал (PDF) 9 октября 2016 г.. Получено 29 ноябрь 2018.
- ^ Мастерсон, Дж. (2007). "Название вида: Solenopsis invicta". Смитсоновский морской вокзал. Архивировано из оригинал 20 сентября 2016 г.. Получено 2 апреля 2016.
- ^ Кармайкл, А. (2005). "Красный импортный огненный муравей" Solenopsis invicta Бурен (Hymenoptera: Formicidae: Myrmicinae) ". PaDIL (Австралийская биозащита). Получено 2 апреля 2016.
- ^ Бухс 2005, п. 13.
- ^ а б Санчи, Ф. (1916). "Формициды sudaméricains nouveaux ou peu connus". Physis. 2: 365–399. Дои:10.5281 / ZENODO.14374.
- ^ Табер 2000, п. 25.
- ^ а б c Shattuck, S.O .; Porter, S.D .; W., D.P. (1999). "Дело 3069. Solenopsis invicta Бурен, 1972 (Insecta, Hymenoptera): предлагается сохранить видовое название ». Вестник зоологической номенклатуры. 56 (1): 27–30. Дои:10.5962 / bhl.part.23022. ISSN 0007-5167.
- ^ Крейтон, W.S. (1930). "Новый мировой вид рода Соленопсис (Hymenop. Formicidae) " (PDF). Труды Американской академии искусств и наук. 66 (2): 39–152. Дои:10.2307/20026320. JSTOR 20026320. Архивировано из оригинал (PDF) 4 марта 2016 г.
- ^ Уилсон, Э. (1952). "The Solenopsis saevissima комплекс в Южной Америке (Hymenoptera: Formicidae) ». Memórias do Instituto Oswaldo Cruz. 50: 60–68. Дои:10.1590 / S0074-02761952000100003. ISSN 0074-0276. PMID 13012835.
- ^ Эттершанк, Г. (1966). "Общая версия мира Myrmicinae, относящаяся к Соленопсис и Фидологетон (Hymenoptera: Formicidae) ". Австралийский зоологический журнал. 14 (1): 73–171. Дои:10.1071 / ZO9660073.
- ^ Кемпф, W.W. (1972). "Catalogo abreviado das formigas da Regiao Neotropical". Studia Entomologica (N.S.). 15: 3–344.
- ^ а б c d е ж грамм час я j k л м п о Бурен, В.Ф. (1972). «Ревизионные исследования по систематике завезенных огненных муравьев». Журнал энтомологического общества Джорджии. 7: 1–26. Дои:10.5281 / zenodo.27055.
- ^ а б c d Трэджер, Дж. К. (1991). "Ревизия огненных муравьев, Solenopsis geminata группа (Hymenoptera: Formicidae: Myrmicinae) ". Журнал Нью-Йоркского энтомологического общества. 99 (2): 141–198. Дои:10.5281 / zenodo.24912. JSTOR 25009890.
- ^ а б Болтон, Б. (1995). Новый общий каталог муравьев мира. Кембридж: Издательство Гарвардского университета. С. 388–391. ISBN 978-0-674-61514-4.
- ^ Табер 2000, п. 26.
- ^ Международная комиссия по зоологической номенклатуре (2001 г.). "Мнение 1976 г. Solenopsis invicta Buren, 1972 (Insecta, Hymenoptera): сохранено видовое название ». Вестник зоологической номенклатуры. 58 (2): 156–157. ISSN 0007-5167.
- ^ Болтон, Б. (2016). "Solenopsis invicta". AntCat. Получено 19 августа 2016.
- ^ а б Чинкель 2006, п. 14.
- ^ а б Росс, К.Г .; Meer, R.K.V .; Fletcher, D.J.C .; Варго, Э. (1987). «Биохимические фенотипические и генетические исследования двух интродуцированных огненных муравьев и их гибридов (Hymenoptera: Formicidae)». Эволюция. 41 (2): 280–293. Дои:10.2307/2409138. JSTOR 2409138. PMID 28568766.
- ^ Росс, К.Г .; Vargo, E.L .; Флетчер, Дэвид Дж. К. (1987). «Сравнительная биохимическая генетика трех видов огненных муравьев в Северной Америке, с особым упором на две социальные формы Solenopsis invicta (Hymenoptera: Formicidae) ". Эволюция. 41 (5): 979–990. Дои:10.2307/2409186. JSTOR 2409186. PMID 28563420.
- ^ Чинкель 2006, п. 15–16.
- ^ а б Pitts, J.P .; McHugh, J.V .; Росс, К. (2005). «Кладистический анализ огненных муравьев Solenopsis saevissima видовая группа (Hymenoptera: Formicidae) ". Zoologica Scripta. 34 (5): 493–505. Дои:10.1111 / j.1463-6409.2005.00203.x.
- ^ Чинкель 2006, п. 503.
- ^ Питтс, Дж. П. (2002). Кладистический анализ видовой группы Solenopsis saevissima (Hymenoptera: Formicidae) (PDF) (Кандидат наук.). Университет Джорджии, Афины. С. 42–43.
- ^ Табер 2000, п. 43.
- ^ Росс, К.Г .; Шумейкер, Д.Д. (1997). «Ядерная и митохондриальная генетическая структура двух социальных форм огненного муравья. Solenopsis invicta: понимание переходов к альтернативной социальной организации ". Наследственность. 78 (6): 590–602. Дои:10.1038 / хди.1997.98. S2CID 19225329.
- ^ Krieger, M.J.B .; Росс, К.Г .; Чанг, C..W.Y .; Келлер, Л. (1999). «Частота и происхождение триплоидии у огненного муравья. Solenopsis invicta". Наследственность. 82 (2): 142–150. Дои:10.1038 / sj.hdy.6884600. S2CID 13144037.
- ^ Keller, L .; Росс, К. (1998). «Эгоистичные гены: зеленая борода у красного огненного муравья» (PDF). Природа. 394 (6693): 573–575. Bibcode:1998Натура.394..573K. Дои:10.1038/29064. S2CID 4310467.
- ^ Графен, А. (1998). «Эволюционная биология: зеленая борода как смертный приговор» (PDF). Природа. 394 (6693): 521–522. Bibcode:1998Натура.394..521Г. Дои:10.1038/28948. S2CID 28124873.
- ^ Wurm, Y .; Wang, J .; Риба-Грогнуз, О .; Корона, М .; Nygaard, S .; Хант, Б.Г .; Инграм, К.К .; Falquet, L .; Nipitwattanaphon, M .; Gotzek, D .; Dijkstra, M.B .; Oettler, J .; Графиня, Ф .; Shih, C.-J .; Wu, W.-J .; Yang, C.-C .; Thomas, J .; Beaudoing, E .; Pradervand, S .; Флегель, В .; Кук, E.D .; Fabbretti, R .; Stockinger, H .; Long, L .; Farmerie, W.G .; Oakey, J .; Boomsma, J.J .; Pamilo, P .; Yi, S.V .; Heinze, J .; Goodisman, M.A.D .; Farinelli, L .; Харшман, К .; Hulo, N .; Cerutti, L .; Xenarios, I .; Shoemaker, D .; Келлер, Л. (2011). «Геном огненного муравья Solenopsis invicta" (PDF). Труды Национальной академии наук. 108 (14): 5679–5684. Bibcode:2011PNAS..108.5679W. Дои:10.1073 / pnas.1009690108. ЧВК 3078418. PMID 21282665.
- ^ Хеджес, С.А. (1997). Морленд, Д. (ред.). Справочник по борьбе с вредителями (8-е изд.). Справочник Маллис и Компания технического обучения. С. 531–535.
- ^ Чинкель 2006, п. 24.
- ^ Бухс 2005, п. 11.
- ^ а б c Petralia, R. S .; Винсон, С. Б. (1979). «Морфология развития личинок и яиц завезенного огненного муравья». Solenopsis invicta". Анналы Энтомологического общества Америки. 72 (4): 472–484. Дои:10.1093 / aesa / 72.4.472.
- ^ а б Wheeler, G.C .; Уилер, Дж. (1977). «Дополнительные исследования личинок муравьев: Myrmicinae». Труды Американского энтомологического общества. 103: 581–602. Дои:10.5281 / zenodo.25100.
- ^ Petralia, R.S .; Соренсен, A.A .; Винсон, С. (1980). «Система губных желез личинок завезенного огненного муравья, Solenopsis invicta Бурень ". Исследования клеток и тканей. 206 (1): 145–156. Дои:10.1007 / BF00233615. PMID 6153574. S2CID 28786982.
- ^ а б Greenberg, L .; Кабашима, Дж. (2014). "Примечания к вредителям: Импортированный красный огненный муравей". Общегосударственная программа комплексной борьбы с вредителями. Сельское хозяйство и природные ресурсы, Калифорнийский университет. Получено 4 апреля 2016.
- ^ О'Нил, Дж .; Маркин, Г. (1975). «Личинки завезенного огненного муравья, Solenopsis invicta Бурен (Hymenoptera: Formicidae) ». Журнал Канзасского энтомологического общества. 48 (2): 141–151. JSTOR 25082731.
- ^ а б c d Фокс, Эдуардо Г. П.; Солис, Дэниел Расс; Росси, Моника Ланцони; Делаби, Жак Юбер Шарль; де Соуза, Родриго Фернандо; Буэно, Одаир Корреа (2012). «Сравнительная морфология неполовозрелых бразильских огненных муравьев (Hymenoptera: Formicidae: Solenopsis)». Психея: журнал энтомологии. 2012: 1–10. Дои:10.1155/2012/183284.
- ^ Mirenda, J.T .; Винсон, С. (1981). «Разделение труда и определение касты в красном импортном огненном муравье». Solenopsis invicta Бурень ". Поведение животных. 29 (2): 410–420. Дои:10.1016 / S0003-3472 (81) 80100-5. S2CID 53199246.
- ^ Шмид-Хемпель 1998, п. 144.
- ^ Hölldobler & Wilson, 1990 г., п. 311.
- ^ Hölldobler & Wilson, 1990 г., п. 318.
- ^ а б Чинкель, W.R. (1988). «Рост колонии и онтогенез полиморфизма рабочих у огненного муравья». Solenopsis invicta". Поведенческая экология и социобиология. 22 (2): 103–115. Дои:10.1007 / BF00303545. S2CID 8455126.
- ^ а б c d е Tschinkel, W.R .; Михеев, А.С .; Сторц, С. (2003). «Аллометрия рабочих муравья-огня, Solenopsis invicta". Журнал науки о насекомых. 3 (2): 2. Дои:10.1673/031.003.0201. ЧВК 524642. PMID 15841219.
- ^ Porter, S.D .; Чинкель, W.R. (1985). «Полиморфизм огненных муравьев: эргономика производства выводков». Поведенческая экология и социобиология. 16 (4): 323–336. Дои:10.1007 / BF00295545. JSTOR 4599785. S2CID 5967997.
- ^ а б Vogt, J.T .; Аппель, А.Г. (2000). «Прерывистый газообмен в муравейнике, Solenopsis invicta Бурен: кастовые различия и температурные эффекты ». Журнал физиологии насекомых. 46 (4): 403–416. Дои:10.1016 / S0022-1910 (99) 00123-7. PMID 12770204.
- ^ а б Клоуден, М.Дж. (2007). Физиологические системы у насекомых. Амстердам; Бостон: Elsevier / Academic Press. С. 357–383, 433–449. ISBN 978-0-12-369493-5.
- ^ Hill, R.W .; Wyse, G.A .; Андерсон, М. (2012). Физиология животных (3-е изд.). Сандерленд, Массачусетс: Sinauer Associates. С. 612–614. ISBN 978-0-87893-662-5.
- ^ а б c d е ж Винсон, С. (1983). «Возвращение к физиологии завезенного огненного муравья». Флоридский энтомолог. 66 (1): 126–139. Дои:10.2307/3494559. JSTOR 3494559.
- ^ Petralia, R.S .; Уильямс, HJ; Винсон, С. (1982). «Ультраструктура задней кишки и продукты выделения личинок завезенного огненного муравья, Solenopsis invicta Бурень ". Насекомые Sociaux. 29 (2): 332–345. Дои:10.1007 / BF02228760. S2CID 7324976.
- ^ Fletcher, D.J.C .; Блюм, М. (1981). "Феромональный контроль деформации и оогенеза у девственных королев огненных муравьев". Наука. 212 (4490): 73–75. Bibcode:1981 Наука ... 212 ... 73F. Дои:10.1126 / science.212.4490.73. PMID 17747633.
- ^ Баркер, Дж. Ф. (1978). «Нейроэндокринная регуляция созревания ооцитов у завезенного огненного муравья». Solenopsis invicta". Общая и сравнительная эндокринология. 35 (3): 234–237. Дои:10.1016/0016-6480(78)90067-9. PMID 689357.
- ^ Jones, R.G .; Davis, W.L .; Hung, A.C.F .; Винсон, С. (1978). «Вызванный осеменением гистолиз летной мускулатуры огненных муравьев (Соленопсис, spp.): ультраструктурное исследование ». Американский журнал анатомии. 151 (4): 603–610. Дои:10.1002 / aja.1001510411. PMID 645619.
- ^ Тоом, П.М.; Johnson, C.P .; Купп, E.W. (1976). «Использование резервов организма во время предпозиционной деятельности Solenopsis invicta". Анналы Энтомологического общества Америки. 69 (1): 145–148. Дои:10.1093 / aesa / 69.1.145.
- ^ а б Винсон, С.Б .; Phillips, S.A .; Уильямс, HJ (1980). «Функция заглоточных желез красного завезенного огненного муравья, Solenopsis invicta бурень ". Журнал физиологии насекомых. 26 (9): 645–650. Дои:10.1016/0022-1910(80)90035-9.
- ^ Vander Meer, R.K .; Лофгрен, К.С. (1989). "Биохимические и поведенческие свидетельства гибридизации огненных муравьев, Solenopsis invicta и Соленопсис Richteri (Hymenoptera: Formicidae) ". Журнал химической экологии. 15 (6): 1757–1765. Дои:10.1007 / BF01012263. PMID 24272179. S2CID 23144401.
- ^ а б Vander Meer, R.K .; Glancey, B.M .; Lofgren, C.S .; Гловер, А .; Tumlinson, J.H .; Рокка, Дж. (1980). «Ядовитый мешок красных импортированных королев огненных муравьев: источник феромонного аттрактанта». Анналы Энтомологического общества Америки. 73 (5): 609–612. Дои:10.1093 / aesa / 73.5.609.
- ^ Choi, M.Y .; Vander Meer, R.K .; Рену, М. (2012). «Биосинтез феромона муравьиных следов запускается нейропептидным гормоном». PLOS ONE. 7 (11): e50400. Bibcode:2012PLoSO ... 750400C. Дои:10.1371 / journal.pone.0050400. ЧВК 3511524. PMID 23226278.
- ^ Эльзен, Г.В. (1986). «Потребление кислорода и потеря воды у импортного огненного муравья. Solenopsis invicta Бурень ". Сравнительная биохимия и физиология A. 84 (1): 13–17. Дои:10.1016/0300-9629(86)90035-6.
- ^ Chen, J .; Рашид, Т .; Feng, G .; Хьюз, В. (2014). "Сравнительное исследование между Solenopsis invicta и Соленопсис Richteri на устойчивость к тепловым и высыхающим нагрузкам ». PLOS ONE. 9 (6): e96842. Bibcode:2014PLoSO ... 996842C. Дои:10.1371 / journal.pone.0096842. ЧВК 4051589. PMID 24915009.
- ^ Boyles, J.G .; Обри, Д.П .; Hickman, C.R .; Мюррей, К.Л .; Timpone, J.C .; Опс, C.H. (2009). "Вариации физиологической реакции завезенных красных огненных муравьев (Solenopsis invicta) к мелкомасштабной тепловой неоднородности ». Журнал термобиологии. 34 (2): 81–84. Дои:10.1016 / j.jtherbio.2008.10.005.
- ^ а б Porter, S.D .; Чинкель, W.R. (1993). «Тепловые предпочтения огненных муравьев: поведенческий контроль роста и обмена веществ». Поведенческая экология и социобиология. 32 (5). Дои:10.1007 / BF00183787. S2CID 9840395.
- ^ а б Vogt, J.T .; Аппель, А.Г. (1999). «Стандартная скорость метаболизма огненного муравья, Solenopsis invicta Бурен: влияние температуры, массы и касты ». Журнал физиологии насекомых. 45 (7): 655–666. Дои:10.1016 / S0022-1910 (99) 00036-0. PMID 12770351.
- ^ а б c d е ж грамм час я j k Веттерер, Дж. (2013). "Экзотическое распространение Solenopsis invicta (Hymenoptera: Formicidae) за пределами Северной Америки ». Социобиология. 60: 50–55. Дои:10.13102 / socialbiology.v60i1.50-55.
- ^ а б Allen, G.E .; Buren, W. F .; Williams, R. N .; Menezes, M.D .; Whitcomb, W.H. (1974). "Красный импортный огненный муравей, Solenopsis invicta; распространение и среда обитания в Мату-Гросу, Бразилия ". Анналы Энтомологического общества Америки. 67 (1): 43–46. Дои:10.1093 / aesa / 67.1.43.
- ^ а б Mescher, M.C .; Росс, К.Г .; Shoemaker, D.D .; Keller, L .; Кригер, М.Дж. Б. (2003). "Распространение двух социальных форм огненного муравья. Solenopsis invicta (Hymenoptera: Formicidae) в ареале коренных жителей Южной Америки " (PDF). Анналы Энтомологического общества Америки. 96 (6): 810–817. Дои:10.1603 / 0013-8746 (2003) 096 [0810: DOTTSF] 2.0.CO; 2.[постоянная мертвая ссылка ]
- ^ а б Ноордейк Дж. (2010). «Анализ риска для огненных муравьев в Нидерландах». Лейден: Европейское исследование беспозвоночных Stichting. п. 15. Получено 5 апреля 2016.
- ^ а б c d е Buren, W.F .; Allen, G.E .; Whitcomb, W.H .; Lennartz, F.E .; Уильямс, Р. (1974). «Зоогеография завезенных огненных муравьев». Журнал Нью-Йоркского энтомологического общества. 82 (2): 113–124. JSTOR 25008914.
- ^ Caldera, E.J .; Росс, К.Г .; DeHeer, Ch.J .; Шумейкер, Д. Д. (2008). "Предполагаемый местный источник инвазивного огненного муравья Solenopsis invicta в США". Биологические вторжения. 10 (8): 1457–1479. Дои:10.1007 / s10530-008-9219-0. S2CID 4471306.
- ^ Pesquero, M.A .; Диас, A.M.P.M (2011). «Географическая переходная зона Соленопсис огненные муравьи (Hymenoptera: Formicidae) и Псевдактеон паразитоиды мух (Diptera: Phoridae) в штате Сан-Паулу, Бразилия ". Неотропическая энтомология. 40 (6): 647–652. Дои:10.1590 / S1519-566X2011000600003. PMID 23939270.
- ^ а б c d "Разновидность: Solenopsis invicta Бурен, 1972 г. ". AntWeb. Калифорнийская академия наук. Получено 5 апреля 2016.
- ^ Уайлд, A.L. (2007). «Каталог муравьев Парагвая (Hymenoptera: Formicidae)» (PDF). Zootaxa. 1622: 1–55. Дои:10.11646 / zootaxa.1622.1.1. ISSN 1175-5334.
- ^ Санчи, Ф. (1923). "Соленопсис et autres fourmis néotropicales ". Revue Suisse de Zoologie. 30 (8): 245–273. Дои:10.5281 / ZENODO.14217.
- ^ Аренс, M.E .; Росс, К.Г .; Шумейкер, Д.Д. (2005). «Филогеографическое строение огненного муравья. Solenopsis invicta в своем родном южноамериканском ареале: роль естественных барьеров и взаимосвязь среды обитания " (PDF). Эволюция. 59 (8): 1733–1743. Дои:10.1111 / j.0014-3820.2005.tb01822.x. PMID 16329243.[постоянная мертвая ссылка ]
- ^ а б c d е ж "Solenopsis invicta (красный импортный огненный муравей) ". Справочник по инвазивным видам. КАБИ. 2014 г.. Получено 12 апреля 2016.
- ^ Табер 2000, п. 28.
- ^ Cokendolpher, J.C .; Филлипс, С.А. младший (1989). "Скорость распространения красного завезенного огненного муравья, Solenopsis invicta (Hymenoptera: Formicidae) в Техасе ". Юго-западный натуралист. 34 (3): 443–449. Дои:10.2307/3672182. JSTOR 3672182.
- ^ "Solenopsis invicta (насекомое)". Глобальная база данных инвазивных видов. Группа специалистов по инвазивным видам. 2010 г.. Получено 2 апреля 2016.
- ^ Бухс 2005, п. 12.
- ^ а б c d е Kemp, S.F .; deShazo, R.D .; Moffitt, J.E .; Williams, D.F .; Бюнер, В.А. (2000). «Расширение среды обитания завезенного огненного муравья (Solenopsis invicta): проблема общественного здравоохранения ". Журнал аллергии и клинической иммунологии. 105 (4): 683–691. Дои:10.1067 / mai.2000.105707. PMID 10756216. S2CID 8280947.
- ^ Desforges, J.F .; deShazo, R.D .; Мясник, Б.Т .; Бэнкс, W.A. (1990). «Реакции на укусы завезенного огненного муравья». Медицинский журнал Новой Англии. 323 (7): 462–466. Дои:10.1056 / NEJM199008163230707. PMID 2197555.
- ^ а б c d е ж грамм час я j k л м п о Collins, L .; Шеффран, Р.Х. (2001). «Красный импортный огненный муравей, Solenopsis invicta Бурен ". Избранные существа UF / IFAS. Университет Флориды. Архивировано из оригинал 21 сентября 2016 г.. Получено 13 апреля 2016.
- ^ "Колонии муравьев и социальная структура | Термикс". www.terminix.com. Получено 19 ноября 2019.
- ^ Хаббард, доктор медицины; Каннингем, W.G. (1977). «Ориентация холмика в муравейнике. Solenopsis invicta (Hymenoptera, Formicidae, Myrmicinae) ». Насекомые Sociaux. 24 (1): 3–7. Дои:10.1007 / BF02223276. S2CID 5851856.
- ^ Penick, C.A .; Чинкель, В. Р. (2008). «Терморегулирующий перенос расплода в муравейнике», Solenopsis invicta". Насекомые Sociaux. 55 (2): 176–182. Дои:10.1007 / s00040-008-0987-4. S2CID 15284230.
- ^ Cassill, D .; Tschinkel, W.R .; Винсон, С. (2002). "Сложность гнезда, размер группы и выращивание расплода у огненного муравья, Solenopsis invicta". Насекомые Sociaux. 49 (2): 158–163. Дои:10.1007 / s00040-002-8296-9. HDL:10806/3290. S2CID 13938249.
- ^ Табер 2000, п. 29.
- ^ Табер 2000, п. 31.
- ^ «100 наихудших инвазивных чужеродных видов в мире». Глобальная база данных инвазивных видов. Группа специалистов по инвазивным видам. Получено 26 января 2014.
- ^ Моаллем Дж. (5 декабря 2013 г.). "Есть причина, по которой их называют сумасшедшими муравьями'". Нью-Йорк Таймс. Получено 23 августа 2016.
- ^ Фонтан, Х. (8 декабря 2008 г.). «Огненные муравьи побеждают благодаря изменениям земли, а не лучшему построению». Нью-Йорк Таймс. Получено 31 октября 2016.
- ^ King, J.R .; Чинкель, W.R. (2008). «Экспериментальные доказательства того, что антропогенные воздействия вызывают вторжения огненных муравьев и экологические изменения». Труды Национальной академии наук. 105 (51): 20339–20343. Bibcode:2008PNAS..10520339K. Дои:10.1073 / pnas.0809423105. ЧВК 2629336. PMID 19064909.
- ^ Capinera 2008, п. 3116.
- ^ Бухс 2005, п. 20.
- ^ Wilcove, D.S .; Rothstein, D .; Dubow, J .; Филлипс, А .; Лосос, Э. (1998). «Количественная оценка угроз для находящихся под угрозой исчезновения видов в Соединенных Штатах» (PDF). Бионаука. 48 (8): 607–615. Дои:10.2307/1313420. JSTOR 1313420. Архивировано из оригинал (PDF) 12 августа 2011 г.
- ^ Чинкель 2006, п. 28.
- ^ Бухс 2005, п. 9.
- ^ Фонтан, Х. (8 июля 2008 г.). «Отслеживание нашествия муравьев до горстки королев». Нью-Йорк Таймс. Получено 2 ноября 2016.
- ^ Росс, К.Г .; Шумейкер, Д.Д. (2008). «Оценка числа основателей популяции инвазивных насекомых-вредителей: огненных муравьев. Solenopsis invicta в США". Труды Королевского общества B: биологические науки. 275 (1648): 2231–2240. Дои:10.1098 / rspb.2008.0412. ЧВК 2603238. PMID 18577505.
- ^ Чинкель 2006, п. 25.
- ^ Бухс 2005, п. 23.
- ^ Адкинс, Х.Г. (1970). «Завезенный огненный муравей на юге США». Летопись Ассоциации американских географов. 60 (3): 578–592. Дои:10.1111 / j.1467-8306.1970.tb00742.x.
- ^ а б Epperson, D.M .; Аллен, C.R. (2010). «Красный завезенный огненный муравей воздействует на горных членистоногих на юге Миссисипи». Американский натуралист из Мидленда. 163 (1): 54–63. Дои:10.1674/0003-0031-163.1.54. S2CID 12992133.
- ^ Callcott, A.-M .; Коллинз, Х.Л. (1996). «Нашествие и расширение ареала завезенных огненных муравьев (Hymenoptera: Formicidae) в Северной Америке в 1918–1995 гг.». Флоридский энтомолог. 79 (2): 240–251. Дои:10.2307/3495821. JSTOR 3495821.
- ^ Корзухин, д.м.н .; Porter, S.D .; Thompson, L.C .; Уайли, С. (2001). "Моделирование пределов диапазона для пожарного муравья, зависящего от температуры. Solenopsis invicta (Hymenoptera: Formicidae) в Соединенных Штатах " (PDF). Экологическая энтомология. 30 (4): 645–655. Дои:10.1603 / 0046-225X-30.4.645. S2CID 53065271.
- ^ Callcott, A.-M .; Oi, D.H .; Collins, H.L .; Williams, D.F .; Локли, Т. (2000). «Сезонные исследования изолированной популяции завезенных красных огненных муравьев (Hymenoptera: Formicidae) в восточном Теннесси» (PDF). Экологическая энтомология. 29 (4): 788–794. Дои:10.1603 / 0046-225X-29.4.788. S2CID 53073021.
- ^ Pimm, S.L .; Бартелл, Д. (1980). «Статистическая модель для прогнозирования расширения ареала завезенного красного огненного муравья, Solenopsis invicta, в Техасе ". Экологическая энтомология. 9 (5): 653–658. Дои:10.1093 / ee / 9.5.653.
- ^ Capinera 2008 С. 2034–2035.
- ^ Моррисон, L.W .; Корзухин, д.м.н .; Портер, С. (2005). "Прогнозируемое расширение диапазона агрессивного огненного муравья, Solenopsis invicta, в восточной части США на основе сценария глобального потепления VEMAP ". Разнообразие и распределения. 11 (3): 199–204. Дои:10.1111 / j.1366-9516.2005.00142.x. S2CID 46089176.
- ^ а б Моррисон, L.W .; Porter, S.D .; Daniels, E .; Корзухин, М.Д. (2004). "Возможное глобальное расширение ареала инвазивных огненных муравьев, Solenopsis invicta" (PDF). Биологические вторжения. 6 (2): 183–191. Дои:10.1023 / B: BINV.0000022135.96042.90. S2CID 9973667.[постоянная мертвая ссылка ]
- ^ Бухс 2005, п. 5.
- ^ «Красный импортный огненный муравей - Solenopsis invicta". Департамент окружающей среды. Правительство Австралии. Архивировано из оригинал 21 сентября 2016 г.. Получено 21 марта 2015.
- ^ McCubbin, K.I .; Вайнер, Дж. М. (2002). «Огненные муравьи в Австралии: новая медицинская и экологическая опасность». Медицинский журнал Австралии. 176 (11): 518–519. Дои:10.5694 / j.1326-5377.2002.tb04547.x. PMID 12064981.
- ^ Хеншоу, Майкл Т; Kunzmann, N .; Vanderwoude, C .; Sanetra, M .; Крозье, Р. Х (2005). «Популяционная генетика и история завезенного огненного муравья», Solenopsis invicta Бурен (Hymenoptera: Formicidae) в Австралии ». Австралийский журнал энтомологии. 44 (1): 37–44. Дои:10.1111 / j.1440-6055.2005.00421.x.
- ^ Кондон, М. (27 июля 2013 г.). «Квинсленд начал войну против нашествия огненных муравьев, но 12 лет спустя они все еще в походе». Курьерская почта. Получено 30 декабря 2014.
- ^ Уиллис, П. (22 февраля 2007 г.). «Обновление огненного муравья». ABC News. Архивировано из оригинал 21 сентября 2016 г.. Получено 4 мая 2015.
- ^ «Национальная программа уничтожения огненных муравьев». Департамент сырьевых отраслей. Правительство Квинсленда. 2007. Архивировано с оригинал 21 октября 2009 г.
- ^ "Ввоз красных огненных муравьев Solenopsis invicta Бурен 1972 - профиль ». Управление окружающей среды и наследия. Правительство штата Новый Южный Уэльс. 2014. Архивировано с оригинал 21 сентября 2016 г.. Получено 5 мая 2015.
- ^ Кридон, К. (4 декабря 2014 г.). «Гонка на время по карантину в Сиднее, вспышка красных огненных муравьев». 9 Новости. Сидней, Австралия. Архивировано из оригинал 21 сентября 2016 г.. Получено 6 декабря 2014.
- ^ Австралийское агентство Associated Press (25 февраля 2016 г.). «Американские огненные муравьи найдены в аэропорту Брисбена». Новости SBS. Архивировано из оригинал 21 сентября 2016 г.. Получено 23 августа 2016.
- ^ Moloney, S .; Вандервуде, К. (2002). «Импортированные красные огненные муравьи: угроза дикой природе восточной Австралии?». Экологический менеджмент и восстановление. 3 (3): 167–175. Дои:10.1046 / j.1442-8903.2002.t01-1-00109.x. S2CID 54926916.
- ^ Rajagopal, T .; Севаркодионе, С.П .; Секар, М. (2005). «Индекс видового богатства, разнообразия и сходства муравьев в пяти выбранных населенных пунктах Саттур Талук». Индийский журнал экологического образования. 5: 7–12.
- ^ Na, J.P.S .; Ли, C.Y. (2001). «Ключ к идентификации обычных городских муравьев-вредителей в Малайзии» (PDF). Тропическая биомедицина. 18 (1): 1–17.
- ^ Куо, К. (2008). Борьба с красными инвазивными огненными муравьями и плодовыми мухами: опыт Тайваня (PDF). Тайбэй, Тайвань: Технологический центр пищевых продуктов и удобрений (FFTC). п. 1.
- ^ Bharti, H .; Guénard, B .; Bharti, M .; Экономо, Э. (2016). «Обновленный список муравьев Индии с их конкретным распространением в штатах Индии (Hymenoptera, Formicidae)». ZooKeys (551): 1–83. Дои:10.3897 / zookeys.551.6767. ЧВК 4741291. PMID 26877665.
- ^ Чинкель 2006, п. 72.
- ^ Wang, L .; Yongyue, L .; Ицзюань, X .; Линг, З. (2013). "Текущее состояние исследований по Solenopsis invicta Бурен (Hymenoptera: Formicidae) в материковом Китае " (PDF). Азиатская мирмекология. 5: 125–137. ISSN 1985-1944.
- ^ Zeng, L .; Lu, Y.Y .; Он, X.F .; Zhang, W.Q .; Лян, Г.В. (2005). «Идентификация красного завезенного огненного муравья. Solenopsis invicta вторжение в материковый Китай и заражение в Учуань, провинция Гуандун ". Китайский вестник энтомологии. 42 (2): 144–148. ISSN 0452-8255.
- ^ Zhang, R .; Li, Y .; Liu, N .; Портер, С. (2007). «Обзор красного импортированного огненного муравья (Hymenoptera: Formicidae) в материковом Китае». Флоридский энтомолог. 90 (4): 723–731. Дои:10.1653 / 0015-4040 (2007) 90 [723: aootri] 2.0.co; 2. Получено 12 апреля 2016.
- ^ Sutherst, R.W .; Мэйвальд, Г. (2005). "Климатическая модель красного импортного огненного муравья, Solenopsis invicta Бурен (Hymenoptera: Formicidae): последствия для вторжения в новые регионы, особенно в Океанию ". Экологическая энтомология. 34 (2): 317–335. Дои:10.1603 / 0046-225X-34.2.317. S2CID 85951260.
- ^ Бурен, В.Ф. (1982). «Красный импортированный огненный муравей теперь в Пуэрто-Рико». Флоридский энтомолог. 65 (1): 188–189. Дои:10.2307/3494163. JSTOR 3494163.
- ^ Wetterer, J.K .; Снеллинг, Р.Р. (2006). "Красный импортный огненный муравей, Solenopsis invictaна Виргинских островах (Hymenoptera: Formicidae) " (PDF). Флоридский энтомолог. 89 (4): 431–434. Дои:10.1653 / 0015-4040 (2006) 89 [431: TRIFAS] 2.0.CO; 2.
- ^ Дейруп, М .; Davis, L .; Бакнер, С. (1998). Состав муравьиной фауны трех Багамских островов (PDF). Сан-Сальвадор: Материалы 7-го симпозиума по естественной истории Багамских островов. п. 27. ISBN 978-0-935909-66-1.
- ^ Дэвис, Л.Р .; Vander Meer, R.K .; Портер, С. (2001). «Импортированные красные огненные муравьи расширяют свой ареал по всей Вест-Индии» (PDF). Флоридский энтомолог. 84 (4): 735. Дои:10.2307/3496416. JSTOR 3496416.
- ^ Ascunce, M.S .; Yang, C.-C .; Oakey, J .; Calcaterra, L .; Wu, W.-J .; Shih, C.-J .; Goudet, J .; Росс, К.Г .; Шумейкер, Д. (2011). «Глобальная история нашествия огненного муравья Solenopsis invicta". Наука. 331 (6020): 1066–1068. Bibcode:2011Научный ... 331.1066A. Дои:10.1126 / science.1198734. PMID 21350177. S2CID 28149214.
- ^ Млот, штат Нью-Джерси; Craig, A.T .; Ху, Д.Л. (2011). «Огненные муравьи самостоятельно собираются в водонепроницаемые плоты, чтобы пережить наводнения». Труды Национальной академии наук. 108 (19): 7669–7673. Bibcode:2011ПНАС..108.7669М. Дои:10.1073 / pnas.1016658108. ЧВК 3093451. PMID 21518911.
- ^ Моррилл, W.L. (1974). «Разгон водой красных привозных огненных муравьев». Флоридский энтомолог. 57 (1): 39–42. Дои:10.2307/3493830. JSTOR 3493830.
- ^ Hölldobler & Wilson, 1990 г., п. 171.
- ^ а б Adams, B.J .; Hooper-Bùi, L.M .; Strecker, R.M .; О'Брайен, Д. (2011). "Построение плотов привозным красным огненным муравьем Solenopsis invicta". Журнал науки о насекомых. 11 (171): 171. Дои:10.1673/031.011.17101. ЧВК 3462402. PMID 22950473.
- ^ Хейт, К. (2006). «Защищенность огненного муравья, Solenopsis invicta, увеличивается во время сплава по колонии ". Насекомые Sociaux. 53 (1): 32–36. Дои:10.1007 / s00040-005-0832-у. S2CID 24420242.
- ^ Howard, D.F .; Чинкель, W.R. (1976). "Аспекты некрофорного поведения у красного импортированного огненного муравья, Solenopsis invicta" (PDF). Поведение. 56 (1): 157–178. Дои:10.1163 / 156853976x00334. ISSN 0005-7959.
- ^ Qiu, H.-L .; Lu, L.-H .; Ши, Q.-X .; Tu, C.-C .; Lin, T .; Он, Я.-Р. (2015). «Дифференциальное некрофорное поведение муравья. Solenopsis invicta на зараженные грибком трупы рабочих и куколок ». Бюллетень энтомологических исследований. 105 (5): 607–614. Дои:10.1017 / S0007485315000528. PMID 26082426.
- ^ Smith, C.W .; Фредериксен, Р.А. (2000). Сорго: происхождение, история, технология и производство. Нью-Йорк, Нью-Йорк: John Wiley & Sons. п. 456. ISBN 978-0-471-24237-6.
- ^ Zettler, J.A .; Spira, T.P .; Аллен, C.R. (2001). «Взаимодействие с муравьиной семенами: могут ли красные импортированные огненные муравьи испортить отношения?». Биологическое сохранение. 101 (2): 249–253. Дои:10.1016 / S0006-3207 (01) 00074-X.
- ^ Камберленд, M.S .; Киркман, Л. (2013). «Влияние красного завезенного огненного муравья на судьбу семян в экосистеме длиннолистной сосны». Экология растений. 214 (5): 717–724. Дои:10.1007 / s11258-013-0201-2. S2CID 2648579.
- ^ Маркин, Г.П .; О'Нил, Дж .; Диллиер, Дж. (1975). "Пищевые туннели красного импортированного огненного муравья, Solenopsis invicta (Hymenoptera: Formicidae) ". Журнал Канзасского энтомологического общества. 48 (1): 83–89. JSTOR 25082717.
- ^ а б c d Porter, S.D .; Чинкель, W.R. (1987). "Собираться в Solenopsis invicta (Hymenoptera: Formicidae): влияние погоды и времени года ». Экологическая энтомология. 16 (3): 802–808. Дои:10.1093 / ee / 16.3.802.
- ^ Showler, A.T .; Knaus, R.M .; Рейган, Т. (1989). "Собирательная территория завезенного огненного муравья, Solenopsis invicta Бурен, в сургарском тростнике, как определено нейтронно-активационным анализом ". Насекомые Sociaux. 36 (3): 235–239. Дои:10.1007 / BF02226306. S2CID 36832531.
- ^ Gravish, N .; Монаенкова, Д .; Goodisman, M.A.D .; Гольдман, Д. (2013). «Подъем, падение и заклинивание во время передвижения муравья в замкнутом пространстве». Труды Национальной академии наук. 110 (24): 9746–9751. arXiv:1305.5860. Bibcode:2013PNAS..110.9746G. Дои:10.1073 / пнас.1302428110. ЧВК 3683784. PMID 23690589.
- ^ а б Ицзюань, X .; Yongyue, L .; Ling, Z .; Гуанвен, Л. (2007). "Собирательство и вербовка завезенных красных огненных муравьев. Solenopsis invicta Бурен в типичных местообитаниях Южного Китая ». Acta Ecologica Sinica. 27 (3): 855–860. Дои:10.1016 / S1872-2032 (07) 60022-5.
- ^ Чинкель, W.R. (2011). «Организация кормления огненного муравья, Solenopsis invicta". Журнал науки о насекомых. 11 (26): 26. Дои:10.1673/031.011.0126. ЧВК 3391925. PMID 21529150.
- ^ а б c d Вандер Меер, Р. (1983). «Полухимики и красный импортный огненный муравей (Solenopsis invicta Бурен) (Hymenoptera: Formicidae) ». Флоридский энтомолог. 66 (1): 139–161. Дои:10.2307/3494560. JSTOR 3494560.
- ^ Уильямс, HJ; Strand, M.R .; Винсон, С. (1981). "След феромона красного импортного огненного муравья". Solenopsis invicta (Бурен) ". Experientia. 37 (11): 1159–1160. Дои:10.1007 / BF01989893. S2CID 46572483.
- ^ Vander Meer, R.K .; Альварес, Ф .; Лофгрен, К.С. (1988). "Выделение следа феромона вербовки Solenopsis invicta". Журнал химической экологии. 14 (3): 825–838. Дои:10.1007 / BF01018776. PMID 24276134. S2CID 32388563.
- ^ Barlin, M.R .; Blum, M.S .; Бренд, М. (1976). «Феромоны следа огненного муравья: анализ видовой специфичности после газохроматографического фракционирования». Журнал физиологии насекомых. 22 (6): 839–844. Дои:10.1016/0022-1910(76)90253-5.
- ^ Capinera 2008, п. 3445.
- ^ Табер 2000, п. 41.
- ^ Vander Meer, R.K .; Preston, CA; Цой, М.-Ю. (2010). «Выделение пиразинового компонента феромона тревоги от огненного муравья, Solenopsis invicta". Журнал химической экологии. 36 (2): 163–170. Дои:10.1007 / s10886-010-9743-0. PMID 20145982. S2CID 10622781.
- ^ а б Хаббард, доктор медицины (1974). «Влияние материала гнезда и запаха колонии на раскопки муравья. Solenopsis invicta (Hymenoptera: Formicidae) ". Энтомологическое общество Джорджии. 9: 127–132.
- ^ Обин, Мартин С. (1986). "Признаки распознавания сородичей в лабораторных и полевых колониях Solenopsis invicta Бурен (Hymenoptera: Formicidae) ». Журнал химической экологии. 12 (9): 1965–1975. Дои:10.1007 / BF01041858. PMID 24305971. S2CID 19517580.
- ^ Обин, М.С.; Вандер Меер, Р. (1989). «Распознавание сородичей у огненных муравьев (Solenopsis invicta Бурен). Королевы маркируют рабочих? ». Этология. 80 (1–4): 255–264. Дои:10.1111 / j.1439-0310.1989.tb00744.x.
- ^ Vander Meer, R.K .; Preston, CA; Хефец, А. (2008). «Королева регулирует уровень биогенного амина и распознавание сородичей у рабочих муравьев-пожарных, Solenopsis invicta". Naturwissenschaften. 95 (12): 1155–1158. Bibcode:2008NW ..... 95.1155V. Дои:10.1007 / s00114-008-0432-6. PMID 18704354. S2CID 7640153.
- ^ Обин, М.С.; Вандер Меер, Р. (1988). "Источники сигналов распознавания сородичей у импортированного огненного муравья. Solenopsis invicta Бурен (Hymenoptera: Formicidae) ». Поведение животных. 36 (5): 1361–1370. Дои:10.1016 / S0003-3472 (88) 80205-7. S2CID 53205306.
- ^ а б Табер 2000, п. 37.
- ^ Табер 2000, п. 38.
- ^ Tennant, L.E .; Портер, С. (1991). «Сравнение рационов двух видов огненных муравьев (Hymenoptera: Formicidae): твердый и жидкий компоненты». Журнал энтомологической науки. 26 (4): 450–465. Дои:10.18474/0749-8004-26.4.450. ISSN 0749-8004.
- ^ а б Портер, С. (1989). «Влияние диеты на рост колоний лабораторных огненных муравьев (Hymenoptera: Formicidae)». Журнал Канзасского энтомологического общества. 62 (2): 288–291. JSTOR 25085088.
- ^ Lanza, J .; Vargo, E. L .; Pulim, S .; Чанг, Ю. З. (1993). «Предпочтения огненных муравьев Solenopsis invicta и S. geminata (Hymenoptera: Formicidae) для аминокислотных и сахарных компонентов внеклеточных нектаров ». Экологическая энтомология. 22 (2): 411–417. Дои:10.1093 / ee / 22.2.411. S2CID 17682096.
- ^ Ланца, Дж. (1991). "Ответ огненных муравьев (Formicidae: Solenopsis invicta и S.gerninata) к искусственным нектарам с аминокислотами ». Экологическая энтомология. 16 (2): 203–210. Дои:10.1111 / j.1365-2311.1991.tb00210.x.
- ^ Vogt, J.T .; Grantham, R.A .; Corbett, E .; Rice, S.A .; Райт, Р. (2002). "Диетические привычки Solenopsis invicta (Hymenoptera: Formicidae) в четырех местообитаниях Оклахомы ". Экологическая энтомология. 31 (1): 47–53. Дои:10.1603 / 0046-225X-31.1.47. S2CID 85586177.
- ^ а б Cassill, D.L .; Чинкель, W.R. (1999). «Регулирование диеты у огненного муравья, Solenopsis invicta" (PDF). Журнал поведения насекомых. 12 (3): 307–328. Дои:10.1023 / А: 1020835304713. S2CID 18581840.
- ^ Howard, D.F .; Чинкель, W.R. (1980). "Влияние размера колонии и голода на поток пищи у огненного муравья, Solenopsis invicta (Hymenoptera: Formicidae) ". Поведенческая экология и социобиология. 7 (4): 293–300. Дои:10.1007 / BF00300670. JSTOR 4599344. S2CID 18507664.
- ^ Соренсен, A.A .; Винсон, С. (1981). "Количественные исследования распределения пищи в лабораторных колониях завезенных огненных муравьев, Solenopsis invicta Бурень ". Насекомые Sociaux. 28 (2): 129–160. Дои:10.1007 / BF02223701. S2CID 28896246.
- ^ Gayahan, G.G .; Чинкель, W.R. (2008). "Огненные муравьи, Solenopsis invicta, высушите и сохраните кусочки насекомых для дальнейшего использования ». Журнал науки о насекомых. 8 (39): 39. Дои:10.1673/031.008.3901. ЧВК 3127378.
- ^ Табер 2000, п. 39.
- ^ Cassill, D.L .; Чинкель, W.R. (1995). «Распределение жидкой пищи между личинками через трофалаксис в колониях муравьев-огнедышащих, Solenopsis invicta". Поведение животных. 50 (3): 801–813. Дои:10.1016/0003-3472(95)80140-5. S2CID 18395869.
- ^ а б c d е Whitcomb, W.H .; Bhatkar, A .; Никерсон, Дж. К. (1973). "Хищники Solenopsis invicta маток до успешного создания колонии ". Экологическая энтомология. 2 (6): 1101–1103. Дои:10.1093 / ее / 2.6.1101.
- ^ Nyffeler, M .; Dean, D.A .; Стерлинг, W.L. (1988). "Южный паук черная вдова, Latrodectus mactans (Araneae, Theridiidae), как хищник красного завезенного огненного муравья, Solenopsis invicta (Hymenoptera, Formicidae) на хлопковых полях Техаса ". Журнал прикладной энтомологии. 106 (1–5): 52–57. Дои:10.1111 / j.1439-0418.1988.tb00563.x.
- ^ Дейруп, М .; Трагер, Дж. (1986). «Муравьи с биологической станции Арчболд, округ Хайлендс, Флорида (Hymenoptera: Formicidae)». Флоридский энтомолог. 69 (1): 206–228. Дои:10.2307/3494763. JSTOR 3494763.
- ^ а б Nickerson, J.C .; Whitcomb, W.H .; Bhatkar, A.P .; Навес, М.А. (1975). "Хищничество королев-основательниц Solenopsis invicta работниками Conomyrma insana". Флоридский энтомолог. 58 (2): 75–82. Дои:10.2307/3493384. JSTOR 3493384.
- ^ а б c Nichols, B.J .; Сайты, R.W. (1991). "Муравьиные хищники королев-основательниц Solenopsis invicta (Hymenoptera: Formicidae) в центральном Техасе ". Экологическая энтомология. 20 (4): 1024–1029. Дои:10.1093 / ее / 20.4.1024.
- ^ Jerome, C.A .; McInnes, D.A .; Адамс, Э. (1998). "Групповая защита королевами-основателями колонии в огненном муравье. Solenopsis invicta". Поведенческая экология. 9 (3): 301–308. Дои:10.1093 / beheco / 9.3.301.
- ^ Рао, Аша; Винсон, С. Б. (2004). "Способность местных муравьев уничтожать небольшие колонии Solenopsis invicta (Hymenoptera: Formicidae) ". Экологическая энтомология. 33 (3): 587–598. Дои:10.1603 / 0046-225X-33.3.587. S2CID 85657533.
- ^ Чен, Ли; Sharma, K.R .; Фадамиро, Х. (2009). "Алкалоиды яда огненных муравьев служат ключевыми аттрактантами для паразитических форидных мух, Псевдактеон трехстворчатый (Diptera: Phoridae) ". Naturwissenschaften. 96 (12): 1421–1429. Bibcode:2009NW ..... 96.1421C. Дои:10.1007 / s00114-009-0598-6. PMID 19714317. S2CID 24205012.
- ^ а б Williams, D.F .; Бэнкс, W.A. (1987). "Pseudacteon obtusus (Diptera: Phoridae) нападающие Solenopsis invicta (Hymenoptera: Formicidae) в Бразилии ». Психея: журнал энтомологии. 94 (1–2): 9–13. Дои:10.1155/1987/85157.
- ^ Эренберг, Р. (18 сентября 2009 г.). «Яд привлекает обезглавленных мух». Новости науки. Архивировано из оригинал 22 сентября 2016 г.. Получено 30 декабря 2014.(требуется подписка)
- ^ Моррисон, L.W .; Портер, С. (2005). «Тестирование воздействия внедренного Псевдактеон трехстворчатый мухи, форидные паразитоиды Solenopsis invicta Огненные муравьи". Биологический контроль. 33 (1): 9–19. CiteSeerX 10.1.1.542.4377. Дои:10.1016 / j.biocontrol.2005.01.004.
- ^ Шмид-Хемпель 1998, п. 94.
- ^ Kathirithamby, J .; Джонстон, Дж. (1992). «Стилопизация Solenopsis invicta (Hymenoptera: Formicidae) по Caenocholax fenyesi (Strepsiptera: Myrmecolacidae) в Техасе ". Анналы Энтомологического общества Америки. 85 (3): 293–297. Дои:10.1093 / aesa / 85.3.293.
- ^ Bruce, W.A .; ЛеКато, Г.Л. (1980). "Pyemotes tritici: Потенциальный новый агент для биологической борьбы с завезенным красным огненным муравьем. Solenopsis invicta (Acari: Pyemotidae) ". Международный журнал акарологии. 6 (4): 271–274. Дои:10.1080/01647958008683230.
- ^ Аренс, M.E .; Шумейкер, Д. (2005). «Эволюционная история Вольбахия инфекции у огненного муравья Solenopsis invicta". BMC Эволюционная биология. 5: 35. Дои:10.1186/1471-2148-5-35. ЧВК 1175846. PMID 15927071.
- ^ Шмид-Хемпель 1998, п. 34.
- ^ Capinera 2008, п. 188.
- ^ Шмид-Хемпель 1998, п. 59.
- ^ а б Briano, J .; Calcaterra, L .; Вароне, Л. (2012). "Огненные муравьи (Соленопсис spp.) и их естественных врагов в южной части Южной Америки ». Психея: журнал энтомологии. 2012: 1–19. Дои:10.1155/2012/198084.
- ^ Jouvenaz, D.P .; Эллис, Э.А. (1986). "Vairimorpha invictae N. Sp. (Microspora: Microsporida), паразит красного завезенного огненного муравья, Solenopsis invicta Бурен (Hymenoptera: Formicidae) ». Журнал протозоологии. 33 (4): 457–461. Дои:10.1111 / j.1550-7408.1986.tb05641.x.
- ^ Oi, D.H .; Briano, J. A .; Valles, S.M .; Уильямс, Д.Ф. (2005). "Передача Vairimorpha invictae (Microsporidia: Burenellidae) инфекции между колониями завезенных красных огненных муравьев (Hymenoptera: Formicidae) ". Журнал патологии беспозвоночных. 88 (2): 108–115. Дои:10.1016 / j.jip.2004.11.006. PMID 15766927.
- ^ Nickle, W.R .; Жувена, Д. (1987). "Тетрадонема соленопсис п. sp. (Nematoda: Tetradonematidae) паразитирует на завезенных красных огненных муравьях. Solenopsis invicta Бурен из Бразилии ». Журнал нематологии. 19 (3): 311–313. ЧВК 2618658. PMID 19290149.
- ^ Qiu, H.-L .; Lu, L.-H .; Zalucki, M.-P .; Он, Я.-Р. (2016). "Metarhizium anisopliae инфекция изменяет пищевое и трофалактическое поведение муравья Solenopsis invicta" (PDF). Журнал патологии беспозвоночных. 138: 24–29. Дои:10.1016 / j.jip.2016.05.005. PMID 27234423.
- ^ Дарем, С. (2010). «Коллекции паразитов ARS помогают в исследованиях и диагностике». Служба сельскохозяйственных исследований. Министерство сельского хозяйства США. Получено 25 августа 2014.
- ^ Knell, J.D .; Allen, G.E .; Азар, Э. (1977). «Световое и электронное микроскопическое исследование Thelohania solenopsae п. sp. (Microsporida: Protozoa) у красного импортного огненного муравья, Solenopsis invicta". Журнал патологии беспозвоночных. 29 (2): 192–200. Дои:10.1016/0022-2011(77)90193-8. PMID 850074.
- ^ «Юг борется с огненными муравьями с патогеном». CNN. 1 июня 1998 г.. Получено 1 ноября 2016.
- ^ Шмид-Хемпель 1998, п. 295.
- ^ Перейра, Р. (2004). "Появление Myrmicinosporidium durum в красном импортном огненном муравье, Solenopsis invicta, и другие новые муравьи-хозяева в восточной части Соединенных Штатов ". Журнал патологии беспозвоночных. 86 (1–2): 38–44. Дои:10.1016 / j.jip.2004.03.005. PMID 15145249.
- ^ Шмид-Хемпель 1998, п. 44.
- ^ Чинкель 2006, п. 632.
- ^ Wang, L .; Эллиотт, В .; Джин, X .; Zeng, L .; Чен, Дж. (2015). «Антимикробные свойства летучих компонентов гнезд красных привозных огненных муравьев, Solenopsis invicta (перепончатокрылые: formicidae) ". Naturwissenschaften. 102 (11–12): 66. Bibcode:2015SciNa.102 ... 66Вт. Дои:10.1007 / s00114-015-1316-1. PMID 26467352. S2CID 1650319.
- ^ Valles, S.M .; Strong, C.A .; Dang, P.M .; Хантер, W.B .; Pereira, R.M .; Oi, D.H .; Шапиро, A.M .; Уильямс, Д.Ф. (2004). "Пикорноподобный вирус от красного завезенного огненного муравья, Solenopsis invicta: первоначальное открытие, последовательность генома и характеристика " (PDF). Вирусология. 328 (1): 151–157. Дои:10.1016 / j.virol.2004.07.016. PMID 15380366.
- ^ Valles, S.M .; Strong, C.A .; Oi, D.H .; Porter, S.D .; Pereira, R.M .; Vander Meer, R.K .; Хашимото, Й .; Hooper-Bùi, L.M .; Sánchez-Arroyo, H .; Дэвис, Т .; Karpakakunjaram, V .; Vail, K.M .; Fudd Graham, L.C .; Briano, J.A .; Calcaterra, L.A .; Gilbert, L.E .; Ward, R .; Ward, K .; Оливер, J.B .; Taniguchi, G .; Томпсон, округ Колумбия (2007). "Фенология, распространение и специфичность хозяина Solenopsis invicta вирус-1 ". Журнал патологии беспозвоночных. 96 (1): 18–27. Дои:10.1016 / j.jip.2007.02.006. PMID 17412359.
- ^ "Огненные муравьи, возможно, встретили себе пару". CNN. Ассошиэйтед Пресс. 7 мая 2007 г. Архивировано с оригинал 13 мая 2007 г.
- ^ Valles, S.M .; Хашимото, Ю. (2009). "Выделение и характеристика Solenopsis invicta вирус 3, новый вирус РНК с положительной цепью, заражающий завезенного красного огненного муравья, Solenopsis invicta". Вирусология. 388 (2): 354–61. Дои:10.1016 / j.virol.2009.03.028. PMID 19403154.
- ^ Allen, C .; Valles, S.M .; Стронг, К.А. (2011). «Множественные вирусные инфекции возникают при индивидуальной полигине и моногине. Solenopsis invicta муравьи ". Журнал патологии беспозвоночных. 107 (2): 107–111. Дои:10.1016 / j.jip.2011.03.005. PMID 21439294.
- ^ а б c Трагер 1988, п. 419.
- ^ Бухс 2005, п. 21.
- ^ Моррилл, W.L. (1974). «Добыча и лёт крыловидно-красных привозных огненных муравьев». Экологическая энтомология. 3 (2): 265–271. Дои:10.1093 / ее / 3.2.265.
- ^ а б Алонсо, L.E .; Вандер Меер, Р. (1997). "Источник крыловидных возбуждающих феромонов в красном импортном огненном муравье. Solenopsis invicta (Hymenoptera: Formicidae) ". Журнал поведения насекомых. 10 (4): 541–555. Дои:10.1007 / BF02765376. S2CID 12295884.
- ^ Маркин, Г.П .; Dillier, J.H .; Hill, S.O .; Blum, M.S .; Германн, Х.Р. (1971). «Брачный полет и дальность полета завезенного огненного муравья, Соленопсис saevissima richteri (Hymenoptera: Formicidae) ". Журнал энтомологического общества Джорджии. 6 (3): 145–156.
- ^ Обин, М.С.; Вандер Меер, Р. (1994). «Полуохимикаты Alate снимают поведение рабочих во время брачных полетов и пожаров» (PDF). Журнал энтомологической науки. 29 (1): 143–151. Дои:10.18474/0749-8004-29.1.143.
- ^ Hung, A.C.F .; Винсон, С.Б .; Саммерлин, Дж. У. (1974). «Мужское бесплодие у красного импортного огненного муравья, Solenopsis invicta". Анналы Энтомологического общества Америки. 67 (6): 909–912. Дои:10.1093 / aesa / 67.6.909.
- ^ Трагер 1988, п. 422.
- ^ Capinera 2008, п. 3444.
- ^ Capinera 2008, п. 3112.
- ^ Hölldobler & Wilson, 1990 г., п. 156.
- ^ Fritz, G.N .; Vander Meer, R.K .; Престон, К.А. (2006). "Избирательная мужская смертность красного завезенного огненного муравья, Solenopsis invicta". Генетика. 173 (1): 207–213. Дои:10.1534 / генетика.106.056358. ЧВК 1461442. PMID 16489215.
- ^ Hölldobler & Wilson, 1990 г., п. 145.
- ^ Чинкель, В. (1992). "Набег на выводок и динамика популяции основанных и зарождающихся колоний огненных муравьев, Solenopsis invicta". Экологическая энтомология. 17 (2): 179–188. Дои:10.1111 / j.1365-2311.1992.tb01176.x.
- ^ Goodisman, M .; Росс, К. (1999). "Вербовка королевы в популяции огненных муравьев с несколькими королевами. Solenopsis invicta". Поведенческая экология. 10 (4): 428–435. Дои:10.1093 / beheco / 10.4.428.
- ^ Tschinkel, W.R .; Ховард, Д.Ф. (1983). "Колония, основанная плеометрозом у огненного муравья, Solenopsis invicta" (PDF). Поведенческая экология и социобиология. 12 (2): 103–113. Дои:10.1007 / BF00343200. S2CID 3017347.
- ^ Трагер 1988, п. 217.
- ^ а б Трагер 1988, п. 218.
- ^ Manfredini, F .; Риба-Грогнуз, О .; Wurm, Y .; Keller, L .; Shoemaker, D .; Грозингер, К.; Цуцуи, Н. Д. (2013). «Социогеномика сотрудничества и конфликта при основании колонии в огненном муравье. Solenopsis invicta". PLOS Genetics. 9 (8): e1003633. Дои:10.1371 / journal.pgen.1003633. ЧВК 3738511. PMID 23950725.
- ^ Bernasconi, G .; Келлер, Л. (1996). «Репродуктивные конфликты в кооперативных ассоциациях королев-огненных муравьев (Solenopsis invicta)" (PDF). Труды Королевского общества B: биологические науки. 263 (1369): 509–513. Bibcode:1996RSPSB.263..509B. Дои:10.1098 / rspb.1996.0077. S2CID 84276777.
- ^ Табер 2000, п. 42.
- ^ Doebeli, Mi .; Abouheif, E. (2016). «Моделирование эволюционных переходов у социальных насекомых». eLife. 5: e12721. Дои:10.7554 / eLife.12721. ЧВК 4744194. PMID 26780668.
- ^ Hölldobler & Wilson, 1990 г., п. 157.
- ^ Hölldobler & Wilson, 1990 г., п. 168.
- ^ Маркин, Г. П .; Dillier, J. H .; Коллинз, Х. Л. (1973). «Рост и развитие колоний красного завезенного огненного муравья, Solenopsis invicta". Анналы Энтомологического общества Америки. 66 (4): 803–808. Дои:10.1093 / aesa / 66.4.803.
- ^ а б Hölldobler & Wilson, 1990 г., п. 163.
- ^ а б Табер 2000, п. 32.
- ^ Шмид-Хемпель 1998, п. 10.
- ^ Шмид-Хемпель 1998, п. 16.
- ^ Портер, С. (1988). «Влияние температуры на рост колонии и темпы развития муравья, Solenopsis invicta". Журнал физиологии насекомых. 34 (12): 1127–1133. Дои:10.1016/0022-1910(88)90215-6.
- ^ Helms, K.R .; Винсон, С. (2008). «Ресурсы растений и рост колоний у инвазивного муравья: важность производящих пади Hemiptera в передаче углеводов на трофических уровнях». Экологическая энтомология. 37 (2): 487–493. Дои:10.1093 / ee / 37.2.487. PMID 18419921.
- ^ Росс, К.Г .; Флетчер, D.J.C. (1986). «Диплоидная продукция самцов - значительный фактор смертности колоний у огненных муравьев. Solenopsis invicta (Hymenoptera: Formicidae) ". Поведенческая экология и социобиология. 19 (4): 283–291. Дои:10.1007 / BF00300643. S2CID 25932018.
- ^ Wang, L .; Xu, Y .; Di, Z .; Рёнер, Б. (2013). «Как групповое взаимодействие и его разрыв влияют на продолжительность жизни?» (PDF). Библиотека Корнельского университета: 1–28. arXiv:1304.2935. Bibcode:2013arXiv1304.2935W. S2CID 14848627.
- ^ Calabi, P .; Портер, С. (1989). «Рабочее долголетие в огненном муравье. Solenopsis invicta: Эргономические аспекты взаимосвязи между температурой, размером и скоростью метаболизма ». Журнал физиологии насекомых. 35 (8): 643–649. Дои:10.1016/0022-1910(89)90127-3.
- ^ а б Hölldobler & Wilson, 1990 г., п. 170.
- ^ Davies, N.B .; Krebs, J.R .; Уэст, С.А. (2012). Введение в поведенческую культуру (4-е изд.). Чичестер: Вили-Блэквелл. п. 385. ISBN 978-1-4051-1416-5.
- ^ Passera, L .; Aron, S .; Vargo, E.L .; Келлер, Л. (2001). "Королева контроля над соотношением полов у огненных муравьев" (PDF). Наука. 293 (5533): 1308–1310. Bibcode:2001Научный ... 293.1308П. Дои:10.1126 / science.1062076. PMID 11509728. S2CID 7483158.
- ^ Vargo, E.L .; Флетчер, D.J.C. (1986). "Свидетельства того, что королева феромонов контролирует воспроизводство самцов и самок у огненных муравьев, Solenopsis invicta". Журнал сравнительной физиологии А. 159 (6): 741–749. Дои:10.1007 / BF00603727. ISSN 1432-1351. S2CID 1165247.
- ^ а б Чинкель 2006, п. 95.
- ^ Hölldobler & Wilson, 1990 г., п. 187.
- ^ Гудисман, Майкл А. Д .; Росс, К. (1998). "Тестирование моделей набора королевы с использованием ядерных и митохондриальных маркеров у огненного муравья. Solenopsis invicta" (PDF). Эволюция. 52 (5): 1416–1422. Дои:10.2307/2411311. JSTOR 2411311. PMID 28565383. Архивировано из оригинал (PDF) 5 мая 2015 г.. Получено 4 мая 2015.
- ^ Morel, L .; Meer, R.K.V .; Лофгрен, C.S. (1990). "Сравнение распознавания сородичей между моногинными и полигинными популяциями Solenopsis invicta (Hymenoptera: Formicidae) ". Анналы Энтомологического общества Америки. 83 (3): 642–647. Дои:10.1093 / aesa / 83.3.642.
- ^ Шмид-Хемпель 1998, п. 191.
- ^ Трагер 1988, п. 204.
- ^ Fletcher, D.J.C .; Blum, M.S .; Whitt, T.V .; Темпл, Н. (1980). «Моногиния и многоженство в огненном муравье, Solenopsis invicta". Анналы Энтомологического общества Америки. 73 (6): 658–661. Дои:10.1093 / aesa / 73.6.658.
- ^ Greenberg, L .; Fletcher, D.J.C .; Винсон, С. (1985). «Различия в размерах рабочих и распределении насыпей в моногинных и полигинных колониях огненных муравьев. Solenopsis invicta Бурень ". Журнал Канзасского энтомологического общества. 58 (1): 9–18. JSTOR 25084596.
- ^ Харрисон, Р. (1980). «Дисперсные полиморфизмы у насекомых». Ежегодный обзор экологии и систематики. 11 (1): 95–118. Дои:10.1146 / annurev.es.11.110180.000523. JSTOR 2096904.
- ^ DeHeer, C.J .; Goodisman, M.A.D .; Росс, К. (1999). "Стратегии распространения королевы в форме огненного муравья с множеством королев. Solenopsis invicta" (PDF). Американский натуралист. 153 (6): 660–675. Дои:10.1086/303205. ISSN 0003-0147. JSTOR 303205. Архивировано из оригинал (PDF) 24 сентября 2015 г.. Получено 4 мая 2015.
- ^ Келлер, Л. (1995). «Социальная жизнь: парадокс колоний с несколькими королевами». Тенденции в экологии и эволюции. 10 (9): 355–360. Дои:10.1016 / S0169-5347 (00) 89133-8. PMID 21237068.
- ^ а б Росс, Кеннет Г. (1993). «Система размножения огненного муравья. Solenopsis invicta: Влияние на генетическую структуру колонии ". Американский натуралист. 141 (4): 554–76. Дои:10.1086/285491. JSTOR 2462750. PMID 19425999.
- ^ Goodisman, M.A.D .; Санкович, К.А .; Ковач, Дж. Л. (2006). «Генетические и морфологические изменения в пространстве и времени у инвазивных огненных муравьев». Solenopsis invicta" (PDF). Биологические вторжения. 9 (5): 571–584. Дои:10.1007 / s10530-006-9059-8. S2CID 25341015. Архивировано из оригинал (PDF) 24 сентября 2015 г.. Получено 4 мая 2015.
- ^ Adams, E.S .; Балас, М. (1999). "Рабочая дискриминация маток в недавно основанных колониях огненных муравьев. Solenopsis invicta". Поведенческая экология и социобиология. 45 (5): 330–338. Дои:10.1007 / s002650050568. S2CID 23876496.
- ^ Чирино, М.Г .; Gilbert, L.E .; Фольгарайт, П.Дж. (2012). «Поведенческое различие между моногинными и полигинными красными огненными муравьями (Hymenoptera: Formicidae) в их естественном ареале». Анналы Энтомологического общества Америки. 105 (5): 740–745. Дои:10.1603 / AN11073. S2CID 86326110.
- ^ Goodisman, M .; Росс, К. (1997). "Взаимосвязь количества маток и родства маток в колониях с несколькими матками огненного муравья. Solenopsis invicta". Экологическая энтомология. 22 (2): 150–157. Дои:10.1046 / j.1365-2311.1997.t01-1-00052.x.
- ^ Adams, E.S .; Чинкель, W.R. (2001). «Механизмы регуляции численности огненного муравья. Solenopsis invicta: экспериментальное исследование ". Журнал экологии животных. 70 (3): 355–369. Дои:10.1046 / j.1365-2656.2001.00501.x. S2CID 18657087.
- ^ Pracana, R .; Приям, А .; Левантис, I .; Nichols, R.A .; Вурм, Ю. (2017). «Супергенный вариант социальной хромосомы огненного муравья Sb демонстрирует низкое разнообразие, но высокое расхождение с SB». Молекулярная экология. 26 (11): 2864–2879. Дои:10.1111 / mec.14054. ЧВК 5485014. PMID 28220980.
- ^ Wang, J .; Wurm, Y .; Nipitwattanaphon, M .; Риба-Грогнуз, О .; Huang, Y.-C .; Shoemaker, D .; Келлер, Л. (2013). «Y-образная социальная хромосома вызывает альтернативную организацию колоний у огненных муравьев» (PDF). Природа. 493 (7434): 664–668. Bibcode:2013Натура 493..664Вт. Дои:10.1038 / природа11832. PMID 23334415. S2CID 4359286.
- ^ Росс, К.Г .; Krieger, M.J .; Шумейкер, Д.Д. (2003). «Альтернативные генетические основы ключевого социального полиморфизма огненных муравьев». Генетика. 165 (4): 1853–1867. ЧВК 1462884. PMID 14704171.
- ^ "Уменьшение биоразнообразия местной фауны и флоры Австралии из-за завезенного красного огненного муравья, Solenopsis invicta (огненный муравей) ". Департамент окружающей среды и энергетики. Правительство Австралии. Архивировано из оригинал 22 сентября 2016 г.. Получено 10 августа 2016.
- ^ а б c d Wojcik, D .; Allen, C.R .; Brenner, R.J .; Forys, E.A .; Jouvenaz, D.P .; Лутц, Р. (2001). «Красный завезенный огненный муравей: влияние на биоразнообразие» (PDF). Американский энтомолог. 47 (1): 16–23. Дои:10.1093 / ae / 47.1.16. Архивировано из оригинал (PDF) 16 декабря 2010 г.
- ^ Porter, S.D .; Савиньяно, Д.А. (1990). «Нашествие огненных муравьев-полигинных истребляет местных муравьев и разрушает сообщество членистоногих». Экология. 71 (6): 2095–2106. Дои:10.2307/1938623. JSTOR 1938623. S2CID 56244004.
- ^ Стокер, Р.Л .; Grant, W.E .; Брэдли Винсон, С. (1995). "Solenopsis invicta (Hymenoptera: Formicidae) влияние на беспозвоночных, разлагающих падаль в центральном Техасе ». Экологическая энтомология. 24 (4): 817–822. Дои:10.1093 / ee / 24.4.817.
- ^ Ricks, B.L .; Винсон, Б. (1970). «Приемлемость кормления некоторыми насекомыми и различными водорастворимыми соединениями для двух разновидностей импортированного огненного муравья». Журнал экономической энтомологии. 63 (1): 145–148. Дои:10.1093 / jee / 63.1.145.
- ^ Кук, Дж. Л. (2003). "Сохранение биоразнообразия в районе, пораженном завезенным красным огненным муравьем, Solenopsis invicta (Hymenoptera: Formicidae) ". Биоразнообразие и сохранение. 12 (2): 187–195. Дои:10.1023 / А: 1021986927560. S2CID 28425331.
- ^ Массер, М.П .; Грант, W.E. (1986). «Смертность мелких млекопитающих, вызванная пожарными муравьями, в восточно-центральном Техасе». Юго-западный натуралист. 31 (4): 540–542. Дои:10.2307/3671712. JSTOR 3671712.
- ^ Фликингер, Э. (1989). «Наблюдения хищничества красных импортированных огненных муравьев на пойманных в ловушку диких хлопковых крысах». Техасский научный журнал. 41 (2): 223–224.
- ^ Хилл, Э. (1970). «Наблюдения за хищничеством завезенных муравьев на птенцов хлопчатника». Материалы ежегодной конференции Юго-Восточной ассоциации уполномоченных по дичи и рыболовству. 23: 171–181.
- ^ Thawley, C.J .; Лангкильде, Т. (2016). «Инвазивный огненный муравей (Solenopsis invicta) хищничество восточной ограды ящерицы (Sceloporus undulatus) яйца ". Журнал герпетологии. 50 (2): 284–288. Дои:10.1670/15-017. S2CID 85855854.
- ^ Mount, R.H .; Trauth, S.E .; Мейсон, W.H. (1981). "Хищничество красного завезенного огненного муравья, Solenopsis invicta (Hymenoptera: Formicidae), на яйцах ящерицы Cnemidophorus sexlineatus (Squamata: Teiidae) ". Журнал Академии наук Алабамы. 52: 66–70.
- ^ Бартлетт, Д. (1997). «40 лет раздумий о Пейнс-Прери». Рептилии. 5 (7): 68, 70–73.
- ^ Лангкильде, Т. (2009). «Инвазивные огненные муравьи меняют поведение и морфологию местных ящериц». Экология. 90 (1): 208–217. Дои:10.1890/08-0355.1. PMID 19294926. S2CID 670780.
- ^ Брайнер, Дж. (26 января 2009 г.). «Танец ящериц избегает смертельных муравьев». LiveScience. Получено 20 августа 2016.
- ^ Freed, P.S .; Нейтман, К. (1988). "Заметки о хищничестве хьюстонской жабы, находящейся под угрозой исчезновения, Буфо хьюстоненсис". Техасский научный журнал. 40: 454–456.
- ^ Allen, C.R .; Rice, K.G .; Wojcik, D.P .; Персиваль, Х.Ф. (1997). «Эффект отравления импортных красных огненных муравьев на новорожденных американских аллигаторов». Журнал герпетологии. 31 (2): 318–321. Дои:10.2307/1565408. JSTOR 1565408.
- ^ а б Capinera 2008, п. 1080.
- ^ Кастелланос, Адриан А .; Medeiros, Matthew C.I .; Hamer, Gabriel L .; Морроу, Майкл Э .; Юбэнкс, Микки Д .; Тил, Пит Д .; Хамер, Сара А .; Свет, Джессика Э. (2016). "Уменьшение численности мелких млекопитающих и клещей у хозяев в связи с инвазивными завезенными красными огненными муравьями (Solenopsis invicta)". Письма о биологии. 12 (9): 20160463. Дои:10.1098 / rsbl.2016.0463. ЧВК 5046925. PMID 27651533.
- ^ Moloney, S.D .; Вандервуде, К. (2003). «Возможное экологическое воздействие завезенных красных огненных муравьев в восточной Австралии» (PDF). Журнал сельскохозяйственной и городской энтомологии. 20 (3): 131–142.
- ^ Локли, Т. (1995). «Влияние завезенного хищничества огненных муравьев на популяцию наименьшей крачки: исчезающий вид». Юго-западный энтомолог. 20: 517–519.
- ^ Сайкс, П.Дж .; Арнольд, К. (1986). «Красный импортный огненный муравей (Solenopsis invicta) хищник на скале ласточка (Хирундо пирронота) птенцы в восточной части центрального Техаса ". Юго-западный натуралист. 31 (1): 105–106. Дои:10.2307/3670967. JSTOR 3670967.
- ^ Campomizzi, A.J .; Моррисон, M.L .; Farrell, S.L .; Wilkins, R..N .; Drees, B.M .; Паккард, Дж. М. (2009). «Импортированные красные огненные муравьи могут снизить выживаемость гнезд певчих птиц». Кондор. 111 (3): 534–537. Дои:10.1525 / cond.2009.090066. S2CID 84552544.
- ^ Morrow, M.E .; Chester, R.E .; Lehnen, S.E .; Drees, B.M .; Топфер, Дж. Э. (2015). «Косвенное влияние красных импортированных огненных муравьев на выживаемость выводка прерийных кур Аттуотер». Журнал управления дикой природой. 79 (6): 898–906. Дои:10.1002 / jwmg.915. ЧВК 4745021. PMID 26900176.
- ^ Yosef, R .; Лорер, Ф.Э. (1995). "Сорокопуты, красные огненные муравьи и красная сельдь?" (PDF). Кондор. 97 (4): 1053–1056. Дои:10.2307/1369544. JSTOR 1369544.
- ^ «Окружающая среда: борьба с огненным муравьем». Время. 2 ноября 1970 г. Архивировано с оригинал 22 сентября 2016 г.. Получено 28 августа 2016.
- ^ Capinera 2008, п. 1771.
- ^ King, J.R .; Чинкель, W.R. (2006). «Экспериментальные доказательства того, что занесенный огненный муравей, Solenopsis invicta, не подавляет в конкурентной борьбе сопутствующих муравьев в нарушенной среде обитания ". Журнал экологии животных. 75 (6): 1370–1378. Дои:10.1111 / j.1365-2656.2006.01161.x. JSTOR 4125078. PMID 17032369. S2CID 14719552.
- ^ Gao, Y .; Лу, Л-Н .; Он, Y-R .; Qi, GJ .; Чжан, Джей-Кью. (2011). «Помехи конкуренции между завезенным красным огненным муравьем (Solenopsis invicta Buren) и два местных вида муравьев (Hymenoptera: Formicidae) ». Acta Entomologica Sinica. 54 (5): 602–608.
- ^ LeBrun, E.G .; Abbott, J .; Гилберт, Л. (2013). «Импортированный сумасшедший муравей вытесняет импортного огненного муравья, сокращает и гомогенизирует сообщества пастбищных муравьев и членистоногих». Биологические вторжения. 15 (11): 2429–2442. Дои:10.1007 / s10530-013-0463-6. S2CID 15260861.
- ^ Vitone, T .; Лаки, А. (2014). «Тротуарный муравей - Tetramorium caespitum". Избранные существа UF / IFAS. Университет Флориды. Получено 4 апреля 2016.
- ^ King, T.G .; Филлипс, С.А. (1992). «Уничтожение брусчатым муравьем молодых колоний красного завезенного огненного муравья». Энтомологические новости. 103 (3): 72–77. ISSN 0013-872X.
- ^ Chen, Y.C .; Kafle, L .; Ши, Си Джей (2011). "Межвидовая конкуренция между Solenopsis invicta и два местных вида муравьев, Фейдол Фервенс и Monomorium chinense". Журнал экономической энтомологии. 104 (2): 614–621. Дои:10.1603 / ec10240. PMID 21510213. S2CID 44631810.
- ^ Calcaterra, L.A .; Livore, J.P .; Delgado, A .; Бриано, Дж. (2008). «Экологическое доминирование красного завезенного огненного муравья, Solenopsis invicta, в его родном диапазоне ". Oecologia. 156 (2): 411–421. Bibcode:2008Oecol.156..411C. Дои:10.1007 / s00442-008-0997-y. PMID 18305962. S2CID 21842183.[постоянная мертвая ссылка ]
- ^ LeBrun, E.G .; Tillberg, C.V .; Suarez, A.V .; Folgarait, P.J .; Smith, C.R .; Холуэй, Д.А. (2007). «Экспериментальное исследование конкуренции между огненными муравьями и аргентинскими муравьями в их естественном ареале» (PDF). Экология. 88 (1): 63–75. Дои:10.1890 / 0012-9658 (2007) 88 [63: AESOCB] 2.0.CO; 2. PMID 17489455.
- ^ Чжоу, А .; Liang, G .; Lu, Y .; Zeng, L .; Сюй, Ю. (2014). «Межвидовая конкуренция между завезенными красными огненными муравьями, Solenopsis invicta Бурен и муравей-призрак, Tapinoma melanocephalum Фабрициус для ресурсов пади, продуцируемых инвазивным мучнистым червецом, Фенакокк соленопсис Тинсилей ". Взаимодействие членистоногих и растений. 8 (5): 469–474. Дои:10.1007 / s11829-014-9324-1. S2CID 18102670.
- ^ Wu, B .; Wang, L .; Liang, G .; Lu, Y .; Цзэн, Л. (2014). "Механизм продовольственной конкуренции между Solenopsis invicta Бурен и Tapinoma melanocephalum Фабрициус ». Социобиология. 61 (3): 265–273. Дои:10.13102 / socialbiology.v61i3.265-273. Архивировано из оригинал 22 сентября 2016 г.
- ^ Cassill, D.L .; Во, К .; Беккер, Б. (2008). «Молодые пожарные муравьи симулируют смерть и переживают агрессивных соседей». Naturwissenschaften. 95 (7): 617–624. Bibcode:2008NW ..... 95..617C. Дои:10.1007 / s00114-008-0362-3. PMID 18392601. S2CID 2942824.
- ^ Kabashima, J.N .; Greenberg, L .; Rust, M.K .; Пейн, Т. Д. (2007). "Агрессивные взаимодействия между Solenopsis invicta и Линепитема униженная (Hymenoptera: Formicidae) в лабораторных условиях » (PDF). Журнал экономической энтомологии. 100 (1): 148–154. Дои:10.1603 / 0022-0493 (2007) 100 [148: AIBSIA] 2.0.CO; 2. PMID 17370822.
- ^ Capinera 2008, п. 290.
- ^ а б Чжоу, А .; Lu, Y .; Zeng, L .; Xu, Y .; Лян, Г. (2012). "Мутуализм способствует вторжению двух чужеродных видов? Solenopsis invicta и Фенакокк соленопсис". PLOS ONE. 7 (7): e41856. Bibcode:2012PLoSO ... 741856Z. Дои:10.1371 / journal.pone.0041856. ЧВК 3402455. PMID 22911859.
- ^ Каплан, I .; Юбэнкс, доктор медицины (2002). «Разрушение хлопковой тли (Homoptera: Aphididae) - естественная динамика врага завезенными красными огненными муравьями (Hymenoptera: Formicidae)». Экологическая энтомология. 31 (6): 1175–1183. Дои:10.1603 / 0046-225X-31.6.1175. S2CID 34957731.
- ^ Coppler, L.B .; Мерфи, Дж. Ф .; Юбэнкс, доктор медицины (2007). «Красные импортные огненные муравьи (Hymenoptera: Formicidae) увеличивают численность тли в томате». Флоридский энтомолог. 90 (3): 419–425. Дои:10.1653 / 0015-4040 (2007) 90 [419: RIFAHF] 2.0.CO; 2.
- ^ Diaz, R .; Knutson, A .; Бернал, Дж. (2004). «Влияние красного завезенного огненного муравья на плотность популяции хлопковой тли и хищничество яиц совки и совки». Журнал экономической энтомологии. 97 (2): 222–229. Дои:10.1603/0022-0493-97.2.222. PMID 15154439. S2CID 198130004.
- ^ Tedders, W.L .; Reilly, C.C .; Wood, B.W .; Моррисон, Р.К .; Лофгрен, C.S. (1990). "Поведение Solenopsis invicta (Hymenoptera: Formicidae) в садах орехов пекан ". Экологическая энтомология. 19 (1): 44–53. Дои:10.1093 / ee / 19.1.44.
- ^ Табер, Стивен (август 2000 г.). Огненные муравьи (1-е изд.). Издательство Техасского университета A&M. п. 308. ISBN 0890969450
- ^ Подробнее, Daniel R .; Kohlmeier, R E .; Хоффман, Дональд Р. (март 2008 г.). «Смертельная анафилаксия к укусам местных местных огненных муравьев у младенца». Американский журнал судебной медицины и патологии. 29 (1): 62–63. Дои:10.1097 / PAF.0b013e3181651b53. PMID 19749619.
- ^ Фитцджеральд, Кевин Т .; Флуд, Арын А. (ноябрь 2006 г.). «Укусы перепончатокрылых». Клинические методы в практике мелких животных. 21 (4): 194–204. Дои:10.1053 / j.ctsap.2006.10.002. PMID 17265905.
- ^ Роудс, Роберт Б .; Стаффорд, Честер Т .; Джеймс, Фрэнк К. (август 1989 г.). «Исследование фатальных анафилактических реакций на укусы импортных огненных муравьев». Журнал аллергии и клинической иммунологии. 84 (2): 159–162. Дои:10.1016/0091-6749(89)90319-9. PMID 2760357.
- ^ Замиф-Миранда, Дэниел; Fox, Eduardo G.P .; Монтейро, Ана Паула; Гама, Диого; Poublan, Луис Э .; де Араужу, Алмайр Феррейра; Араухо, Мария Ф. К .; Ателла, Джорджия C .; Machado, Ednildo A .; Диас, Бруно Л. (26 сентября 2018 г.). «Аллергический ответ, опосредованный белками яда огненных муравьев». Научные отчеты. 8 (1): 14427. Bibcode:2018НатСР ... 814427Z. Дои:10.1038 / s41598-018-32327-z. ЧВК 6158280. PMID 30258210.
- ^ Фокс E.G.P. (2016) Ядовитые токсины огненных муравьев. В: Gopalakrishnakone P., Calvete J. (eds) Venom Genomics and Proteomics. Токсинология. Спрингер, Дордрехт
- ^ душ Сантуш Пинту, Хосе Р. А .; Fox, Eduardo G.P .; Saidemberg, Daniel M .; Santos, Lucilene D .; да Силва Менегассо, Anally R .; Коста-Мансо, Элиуд; Machado, Ednildo A .; Bueno, Odair C .; Пальма, Марио С. (22 августа 2012 г.). "Протеомный взгляд на яд из огненного муравья Бурена". Журнал протеомных исследований. 11 (9): 4643–4653. Дои:10.1021 / пр300451г. PMID 22881118.
- ^ Baer, H .; Liu, T.-Y .; Anderson, M.C .; Блюм, М .; Schmid, W.H .; Джеймс, Ф.Дж. (1979). «Белковые компоненты яда огненных муравьев (Solenopsis invicta)". Токсикон. 17 (4): 397–405. Дои:10.1016/0041-0101(79)90267-8. PMID 494321.
- ^ Хоффман, Д.Р .; Dove, DE; Якобсон, RS (ноябрь 1988 г.). «Аллергены в яде перепончатокрылых. XX. Выделение четырех аллергенов из импортного яда огненных муравьев (Solenopsis invicta)». Журнал аллергии и клинической иммунологии. 82 (5, Пет. 1): 818–27. Дои:10.1016 / 0091-6749 (88) 90084-X. PMID 3192865.
- ^ Макдональд, М. (2006). «Красные под ногами». Новый ученый. 189 (2538): 50. Архивировано с оригинал 25 апреля 2016 г.
- ^ а б c d «Красный импортированный огненный муравей: объявлен вредителем в Западной Австралии». Департамент сельского хозяйства и продовольствия. Правительство Западной Австралии. 2014. Архивировано с оригинал 22 сентября 2016 г.. Получено 21 августа 2016.
- ^ Пиментель, Д .; Lach, L .; Zuniga, R .; Моррисон, Д. (2000). «Экологические и экономические издержки некоренных видов в Соединенных Штатах». Бионаука. 50 (1): 53–65. Дои:10.1641 / 0006-3568 (2000) 050 [0053: EAECON] 2.3.CO; 2.
- ^ Дай, Дж. (8 декабря 2014 г.). «Вспышка красных огненных муравьев в Сиднее может стоить экономике миллиардов». Sydney Morning Herald. Получено 8 декабря 2014.
- ^ а б c "Огненные муравьи". Сельское хозяйство Виктория. Правительство Виктории. 2004. Архивировано с оригинал 22 сентября 2016 г.. Получено 20 августа 2016.
- ^ Смит, М.Р. (1965). Муравьи-насекомые на востоке США: их признание, биология и экономическое значение (PDF). Вашингтон, округ Колумбия: Министерство сельского хозяйства США. п. 43. OCLC 6078460.
- ^ Stiles, J.H .; Джонс, Р.Х. (1998). «Раздача красного импортного огненного муравья, форма Solenopsis invicta, в местах обитания дорог и линий электропередач ". Ландшафтная Экология. 13 (6): 335–346. Дои:10.1023 / А: 1008073813734. S2CID 21131952.
- ^ "США импортируют врагов огненных муравьев". CNN. 30 ноября 1999 г.. Получено 1 ноября 2016.
- ^ Вандер Меер, Р. К. Вандер; Slowik, T. J .; Торвилсон, Х. Г. (2002). "Семиохимические вещества, выделяемые электрически стимулированными красными импортными огненными муравьями, Solenopsis invicta". Журнал химической экологии. 28 (12): 2585–2600. Дои:10.1023 / А: 1021448522147. PMID 12564802. S2CID 4705569.
- ^ Mackay, W.P .; Винсон, С.Б .; Irving, J .; Majdi, S .; Мессер, К. (1992). «Влияние электрических полей на красного завезенного огненного муравья (Hymenoptera: Formicidae)». Экологическая энтомология. 21 (4): 866–870. Дои:10.1093 / ee / 21.4.866.
- ^ Slowik, T.J .; Green, B.L .; Торвилсон, Х.Г. (1997). «Обнаружение магнетизма у красного импортного огненного муравья (Solenopsis invicta) с помощью магнитно-резонансной томографии ». Биоэлектромагнетизм. 18 (5): 396–399. Дои:10.1002 / (SICI) 1521-186X (1997) 18: 5 <396 :: AID-BEM7> 3.0.CO; 2-Y. PMID 9209721.
- ^ Андерсон, J.B .; Вандер Меер, Р. (1993). «Магнитная ориентация в огненном муравье, Solenopsis invicta". Naturwissenschaften. 80 (12): 568–570. Bibcode:1993NW ..... 80..568A. Дои:10.1007 / BF01149274. S2CID 41502040.
- ^ а б Jetter, K.M .; Гамильтон, Дж .; Klotz, J.H. (2002). «Подсчитаны затраты на ликвидацию: завезенные красные огненные муравьи угрожают сельскому хозяйству, дикой природе и домам» (PDF). Калифорнийское сельское хозяйство. 56 (1): 26–34. Дои:10.3733 / ca.v056n01p26.
- ^ «Удары огненных муравьев - почему они являются проблемой». Департамент сельского хозяйства и рыболовства. Правительство Квинсленда. 2016. Архивировано с оригинал 22 сентября 2016 г.. Получено 20 августа 2016.
- ^ Lofgren, C.S .; Адамс, К. (1981). "Снижение урожайности сои на полях, зараженных красным завезенным огненным муравьем, Solenopsis invicta". Флоридский энтомолог. 64 (1): 199–202. Дои:10.2307/3494619. JSTOR 3494619.
- ^ Бэнкс, W.A .; Adams, C.T .; Lofgren, C.S .; Войчик, Д. (1990). «Заражение соевых бобов на полях сои на юге США».. Флоридский энтомолог. 73 (3): 503–504. Дои:10.2307/3495467. JSTOR 3495467.
- ^ Adams, C.T .; Plumley, J.K .; Lofgren, C.S .; Бэнкс, W.A. (1976). «Экономическое значение красного импортного огненного муравья, Solenopsis invicta Бурен. I. Предварительные исследования влияния на урожай сои ». Журнал энтомологического общества Джорджии. 11: 165–169.
- ^ Adams, C.T .; Plumley, J.K .; Бэнкс, W.A .; Лофгрен, К.С. (1977). «Удар красного импортного огненного муравья, Solenopsis invicta Бурен (Hymenoptera: Formicidae) на сборе сои в Северной Каролине ». Журнал Общества Элиша Митчелла. 93: 150–153.
- ^ Apperson, C.S .; Адамс, К. (1983). «Медицинское и сельскохозяйственное значение привозного красного огненного муравья». Флоридский энтомолог. 66 (1): 121–126. Дои:10.2307/3494558. JSTOR 3494558.
- ^ Capinera 2008, п. 903.
- ^ Бэнкс, W.A .; Adams, C.T .; Лофгрен, C.S. (1991). «Повреждение молодых цитрусовых деревьев завезенным красным огненным муравьем (Hymenoptera: Formicidae)». Журнал экономической энтомологии. 84 (1): 241–246. Дои:10.1093 / jee / 84.1.241.
- ^ Capinera 2008, п. 1084.
- ^ Стерлинг, W.L. (1978). «Случайное биологическое подавление долгоносика красным завезенным огненным муравьем». Экологическая энтомология. 7 (4): 564–568. Дои:10.1093 / ее / 7.4.564.
- ^ Fillman, D.A .; Стерлинг, W.L. (1983). «Убийственная сила красного завезенного огненного муравья [Hym .: Formicidae]: ключевой хищник долгоносика [Col .: Curculionidae]». Энтомофага. 28 (4): 339–344. Дои:10.1007 / BF02372186. S2CID 38550501.
- ^ Sterling, W.L .; Dean, D.A .; Fillman, D.A .; Джонс, Д. (1984). «Естественный биологический контроль над долгоносиком [Col .: Curculionidae]». Энтомофага. 29 (1): 1–9. Дои:10.1007 / BF02372203. S2CID 33045477.
- ^ Negm, A.A .; Хенсли, С. (1967). «Отношение хищников членистоногих к ущербу урожаю, нанесенному мотыльком сахарного тростника». Журнал экономической энтомологии. 60 (6): 1503–1506. Дои:10.1093 / jee / 60.6.1503.
- ^ Reagan, T.E .; Coburn, G .; Хенсли, С. (1972). «Воздействие мирекса на фауну членистоногих в поле сахарного тростника Луизианы». Экологическая энтомология. 1 (5): 588–591. Дои:10.1093 / ee / 1.5.588.
- ^ Capinera 2008, п. 4173.
- ^ Lemke, L.A .; Киссам, Дж. Б. (1988). «Воздействие хищничества красных завезенных огненных муравьев (Hymenoptera: Formicidae) на рогатых мух (Diptera: Muscidae) на пастбищах крупного рогатого скота, обработанных протруном». Журнал экономической энтомологии. 81 (3): 855–858. Дои:10.1093 / jee / 81.3.855. PMID 3403766.
- ^ Lee, J.-H .; Johnson, S.J .; Райт, В. (1990). «Количественный анализ выживаемости куколок бархатных гусениц (Lepidoptera: Noctuidae) на соевых полях в Луизиане». Экологическая энтомология. 19 (4): 978–986. Дои:10.1093 / ee / 19.4.978.
- ^ Моррилл, W.L. (1977). «Красный импортный огненный муравей, добывающий пищу в теплице». Экологическая энтомология. 6 (3): 416–418. Дои:10.1093 / ее / 6.3.416.
- ^ а б c Юбэнкс, доктор медицины (2001). «Оценки прямого и косвенного воздействия красных завезенных огненных муравьев на биологический контроль над полевыми культурами». Биологический контроль. 21 (1): 35–43. Дои:10.1006 / bcon.2001.0923. S2CID 86682078.
- ^ Wilson, N.L .; Олив, AD (1969). «Пищевые привычки завезенных огненных муравьев на пастбищах и в сосновых лесах на юго-востоке Луизианы». Журнал экономической энтомологии. 62 (6): 1268–1271. Дои:10.1093 / jee / 62.6.1268.
- ^ Hu, G.Y .; Франк, Дж. (1996). «Воздействие красного импортированного огненного муравья (Hymenoptera: Formicidae) на членистоногих, населяющих навоз во Флориде». Экологическая энтомология. 25 (6): 1290–1296. Дои:10.1093 / ee / 25.6.1290.
- ^ Лопес, Дж. Д. (1982). "Схема появления зимующей популяции Cardiochiles nigriceps в центральном Техасе ". Экологическая энтомология. 11 (4): 838–842. Дои:10.1093 / ee / 11.4.838.
- ^ Эллиот, Д. (1 июля 2013 г.). ""Безумные муравьи «кишат в юго-восточных штатах». CBS Новости. Получено 22 ноября 2016.
- ^ Lofgren, C.S .; Бэнкс, W.A .; Гланси, Б. (1975). «Биология и борьба с завезенными огненными муравьями». Ежегодный обзор энтомологии. 20 (1): 1–30. Дои:10.1146 / annurev.en.20.010175.000245. PMID 1090234.
- ^ Бухс 2005, стр. 1–2.
- ^ Williams, D.F .; деШазо, Р.Д. (2004). «Биологическая борьба с огненными муравьями: обновленная информация о новых методах». Анналы аллергии, астмы и иммунологии. 93 (1): 15–22. Дои:10.1016 / S1081-1206 (10) 61442-1. PMID 15281467.
- ^ Mottern, J.L; Heinz, K.M; Ода, П.Дж. (2004). «Оценка биологического контроля над огненными муравьями с использованием форидных мух: влияние на конкурентные взаимодействия». Биологический контроль. 30 (3): 566–583. Дои:10.1016 / j.biocontrol.2004.02.006.
- ^ Drees, B.M .; Пакетт, Р. «Потенциальные средства биологической борьбы с завезенным красным огненным муравьем» (PDF). Техасский импортный проект по исследованию и управлению огненных муравьев. Техасский университет A&M. Получено 21 августа 2016.
- ^ Buren, W.F .; Allen, G.E .; Уильямс, Р. (1978). «Подходы к возможному контролю над завезенными огненными муравьями». Бюллетень энтомологического общества Америки. 24 (4): 418–421. Дои:10.1093 / besa / 24.4.418.
- ^ а б c «Интегрированное руководство по борьбе с вредителями: огнестрельное оружие». Служба национальных парков. Министерство внутренних дел США. 2010. Архивировано с оригинал 22 сентября 2016 г.. Получено 21 августа 2016.
- ^ Bonnefoy, X .; Kampen, H .; Суини, К. (2008). Значение городских вредителей для общественного здравоохранения. Копенгаген, Дания: Всемирная организация здравоохранения, Европейское региональное бюро. п. 193. ISBN 978-92-890-7188-8.
- ^ Calixto, A.A .; Харрис, М.К .; Knutson, A .; Барр, К. (2007). "Родные муравьи реагируют на Solenopsis invicta Уменьшение бурена с помощью трансляционных приманок ». Экологическая энтомология. 36 (5): 1112–1123. Дои:10.1603 / 0046-225X (2007) 36 [1112: НАРТСИ] 2.0.CO; 2. PMID 18284735.
- ^ Klotz, J.H .; Vail, K.M .; Уильямс, Д.Ф. (1997). "Токсичность водной приманки борная кислота-сахароза для Solenopsis invicta (Hymenoptera: Formicidae) ". Журнал экономической энтомологии. 90 (2): 488–491. Дои:10.1093 / jee / 90.2.488.
- ^ Benson, E.P .; Zungoli, P.A .; Райли, М. (2003). "Воздействие загрязнителей на прием наживки Solenopsis invicta (Hymenoptera: Formicidae) ". Журнал экономической энтомологии. 96 (1): 94–97. Дои:10.1093 / jee / 96.1.94. PMID 12650350.
Библиография
- Бухс, Дж. Б. (2005). Войны огненных муравьев: природа, наука и общественная политика в Америке двадцатого века. Чикаго, Иллинойс: Издательство Чикагского университета. ISBN 978-0-226-07981-3.CS1 maint: ref = harv (связь)
- Capinera, J.L. (2008). Энциклопедия энтомологии (2-е изд.). Дордрехт, Нидерланды: Springer. ISBN 978-1-4020-6242-1.CS1 maint: ref = harv (связь)
- Hölldobler, B .; Уилсон, Э. (1990). Муравьи. Кембридж, Массачусетс: Belknap Press of Harvard University Press. ISBN 978-0-674-04075-5.CS1 maint: ref = harv (связь)
- Шмид-Хемпель, П. (1998). Паразиты у социальных насекомых. Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-05924-2.CS1 maint: ref = harv (связь)
- Табер, С.В. (2000). Огненные муравьи. Колледж-Стейшн, Техас: Издательство Техасского университета A&M. ISBN 978-1-60344-711-9.CS1 maint: ref = harv (связь)
- Трэджер, Дж. К. (1988). Достижения в мирмекологии (1-е изд.). Лейден, Нидерланды: E.J. Брилл. ISBN 978-0-916846-38-1.CS1 maint: ref = harv (связь)
- Чинкель, В. (2006). Огненные муравьи. Кембридж, Массачусетс: Belknap Press of Harvard University Press. ISBN 978-0-674-02207-2.CS1 maint: ref = harv (связь)
внешняя ссылка
| Библиотечные ресурсы о Красный импортный огненный муравей |
Интернет сайт
- Книги и журналы о красном привозном огненном муравье в Библиотека наследия биоразнообразия
- Книги и журналы о красном привозном огненном муравье в Интернет-архив
- Национальная программа искоренения завезенных красных огненных муравьев - Министерство сельского хозяйства и рыболовства Квинсленда
- Импортные огненные муравьи - Министерство сельского хозяйства США
- Solenopsis invicta в AntWiki - Муравьи в мир
- В S. invicta Геномный проект - Проект базы данных генома перепончатокрылых
- Профиль вида - Красный импортный огненный муравей (Solenopsis invicta), Национальный информационный центр по инвазивным видам, Национальная сельскохозяйственная библиотека США.
Средства массовой информации
- Кадры форидных мух, паразитирующих на огненных муравьях - Захвачено BBC, опубликовано Техасским университетом в Остинском колледже естественных наук.
- Красные огненные муравьи - опубликовано National Geographic на YouTube
- Заливка колонии огненных муравьев из расплавленного алюминия на YouTube
Освещение новостей
- Невероятный летающий огненный муравей – Вашингтон Пост, 2011