Бактериальная систематика - Bacterial taxonomy - Wikipedia
Бактериальная систематика это таксономия, т.е. классифицировать -основанная классификация бактерии.
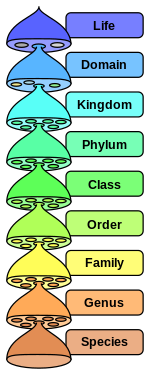
в научная классификация установлен Карл Линней,[1] каждый вид должен быть отнесен к роду (двоичная номенклатура ), что, в свою очередь, является нижним уровнем иерархии разряды (семья, подотряд, порядок, подкласс, класс, подразделение / тип, королевство и домен). классификация жизни есть три домена (Эукариоты, Бактерии и Археи ),[2] которые, с точки зрения таксономии, несмотря на следование одним и тем же принципам, имеют несколько различных соглашений между собой и между своими подразделениями, поскольку они изучаются разными дисциплинами (ботаника, зоология, микология и микробиология). Например, в зоологии есть типовые образцы, а в микробиологии - типовые.
Разнообразие
Прокариоты имеют много общих черт, таких как отсутствие ядерной мембраны, одноклеточность, деление на бинарное деление и, как правило, небольшой размер. Различные виды различаются между собой по нескольким характеристикам, что позволяет их идентифицировать и классифицировать. Примеры включают:
- Филогения: Все бактерии происходят от общего предка и с тех пор разнообразны и, следовательно, обладают разными уровнями эволюционного родства (см. Бактериальный тип и Хронология эволюции )
- Метаболизм: У разных бактерий могут быть разные метаболические способности (см. Микробный метаболизм )
- Среда: Различные бактерии процветают в разных средах, таких как высокая / низкая температура и соль (см. Экстремофилов )
- Морфология: Между бактериями существует множество структурных различий, таких как форма клеток, окраска по Граму (количество липидных бислоев) или двухслойный состав (см. Морфология бактериальных клеток, Структура бактериальной клетки )
- Патогенность: Некоторые бактерии патогенны для растений или животных (см. Патогенные бактерии )
История
Первые описания
Бактерии впервые были обнаружены Антони ван Левенгук в 1676 г., используя однообъективную микроскоп собственного дизайна.[3] Он назвал их "животные "и опубликовал свои наблюдения в серии писем в Королевское общество.[4][5][6]
Ранее описанные роды бактерий включают: Вибрион и Монас, к О. Ф. Мюллер (1773, 1786), затем классифицированный как Инфузория (однако многие виды, ранее входившие в эти роды, сегодня считаются протистами); Полиангиум, к Х. Ф. Линк (1809), первая бактерия, известная до сих пор; Серратия, к Bizio (1823 г.); и Спирилл, Спирохета и Бактерия, к Эренберг (1838).[7][8]
Период, термин Бактерия, введенный как род Эренбергом в 1838 г.,[9] стал универсальным средством для стержневых ячеек.[7]
Ранние формальные классификации
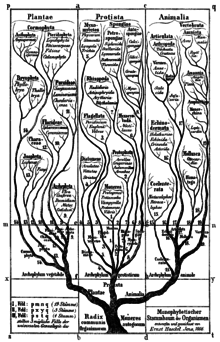
Бактерии сначала были классифицированы как растения составляющий класс Шизомицеты, который вместе с Schizophyceae (сине-зеленые водоросли/Цианобактерии) сформировал тип Шизофита.[11]
Геккель в 1866 г. поместил группу в тип Moneres (от μονήρης: простой) в королевстве Протиста и определяет их как полностью бесструктурные и однородные организмы, состоящие только из кусочка плазмы.[10] Он разделил тип на две группы:[10]
- умереть Gymnomoneren (без конверта)
- Протогены - Такие как Protogenes primordialis, теперь классифицируется как эукариот, а не как бактерия
- Протамаэба - теперь классифицируется как эукариот, а не как бактерия
- Вибрион - род бактерий в форме запятой, впервые описанный в 1854 г.[12])
- Бактерия - род палочковидных бактерий, впервые описанный в 1828 году, который позже дал свое название представителям Monera, ранее называвшимся «монерон» (множественное число «monera») на английском языке и «Эйне Монерен"(фем. пл."Moneres") на немецком
- Бациллы - род спорообразующих палочковидных бактерий, впервые описанных в 1835 году.[13]
- Спирохета – тонкие спиралевидные бактерии впервые описан в 1835 г.[13]
- Спирилл – спиралевидные бактерии впервые описан в 1832 г.[14]
- и Т. Д.
- умереть Лепомонерен (с конвертом)
- Protomonas - теперь классифицируется как эукариот, а не как бактерия. Название было повторно использовано в 1984 году для неродственного рода бактерий.[15]
- Вампирелла - теперь классифицируется как эукариот, а не как бактерия
Классификация Фердинанд Кон (1872) был влиятельным человеком в девятнадцатом веке и признал шесть родов: Микрококк, Бактерия, Бациллы, Вибрион, Спирилл, и Спирохета.[7]
Позднее группа была реклассифицирована как Прокариоты к Chatton.[16]
Классификация Цианобактерии (в просторечии «сине-зеленые водоросли») велась борьба между водорослями или бактериями (например, Геккель классифицировал Носток в типе водорослей Archephyta[10]).
в 1905 году Эрвин Ф. Смит принял 33 различных названия бактериальных родов и более 150 неправильных названий,[17] и Вюйлемен в исследовании 1913 г.[18] пришел к выводу, что все виды Бактерии должен попадать в роды Планококк, Стрептококк, Клебсиелла, Мериста, Планомериста, Neisseria, Сарцина, Planosarcina, Метабактерии, Clostridium, Серратия, Бактерия, и Спирилл.
Кон[19] признал четыре племена: Сферобактерии, микробактерии, десмобактерии и спиробактерии. Станир и ван Нил[20] признал царство Monera с двумя типами, Myxophyta и Schizomycetae, последняя включает классы Eubacteriae (три отряда), Myxobacteriae (один отряд) и Spirochetae (один отряд). Биссет[21] различали 1 класс и 4 порядка: Eubacteriales, Actinomycetales, Streptomycetales и Flexibacteriales. Вальтер Мигула система,[22] которая была наиболее широко принятой системой своего времени и включала все известные тогда виды, но была основана только на морфологии, включала три основные группы Coccaceae, Bacillaceae и Spirillaceae, а также Trichobacterinae для нитчатых бактерий. Орла-Йенсен[23] установили два отряда: Cephalotrichinae (семь семейств) и Peritrichinae (предположительно с одним семейством). Bergey et al.[24] представили классификацию, которая в целом соответствует Заключительному отчету Общества американских бактериологов за 1920 г. (Winslow et al.), в котором класс шизомицетов разделен на четыре отряда: Myxobacteriales, Thiobacteriales, Chlamydobacteriales и Eubacteriales, причем пятая группа состоит из четырех родов, считающихся промежуточными. между бактериями и простейшими: Спирохета, Кристоспира, Сапроспира, и Трепонема.
Однако разные авторы часто переклассифицировали роды из-за отсутствия видимых признаков, что приводило к плохому состоянию, которое было резюмировано в 1915 году Робертом Эрлом Бьюкененом.[25] К тому времени вся группа получила разные звания и имена разными авторами, а именно:
- Шизомицеты (Naegeli 1857)[11]
- Бактерии (Кон 1872 г.)[26]
- Бактерии (Кон 1872 б)[27]
- Schizomycetaceae (Детони и Тревизан, 1889 г.)[28]
Кроме того, семейства, на которые был разделен класс, менялись от автора к автору, и для некоторых, таких как Zipf, имена были на немецком, а не на латыни.[29]
Первое издание Бактериологического кодекса в 1947 году разрешило несколько проблем.[30][пример необходим ]
Система А. Р. Прево[31][32]) имел четыре подтипа и восемь классов, а именно:
- Eubacteriales (классы Asporulales и Sporulales)
- Mycobacteriales (классы Actinomycetales, Myxobacteriales и Azotobacteriales)
- Алгобактерии (классы Siderobacteriales и Thiobacteriales)
- Protozoobacteriales (класс Spirochetales)
Неформальные группы на основе окрашивания по Граму
Несмотря на отсутствие согласия по основным подгруппам Бактерии, Окрашивание по Граму результаты чаще всего использовались в качестве инструмента классификации. Следовательно, до появления молекулярной филогении Королевство Прокариоты был разделен на четыре подразделения,[41] Схема классификации, которой до сих пор формально следует руководство Берджи по систематической бактериологии для порядка следования книг.[42]
- Gracilicutes (грамотрицательный)
- Фотобактерии (фотосинтетический): класс Оксифотобактерии (вода как донор электронов, включает заказ Цианобактерии= сине-зеленые водоросли, теперь тип Цианобактерии) и класс Аноксифотобактерии (анаэробные фототрофы, заказы: Родоспириллы и Хлоробиалес
- Скотобактерии (нефотосинтетический, теперь Протеобактерии и другие грамотрицательные нефотосинтетические типы)
- Firmacutes [sic] (грамположительный, впоследствии исправленный на Фирмикуты[43])
- несколько заказов, таких как Бациллы и Актиномицеты (сейчас в типе Актинобактерии)
- Молликуты (переменная грамма, например Микоплазма)
- Mendocutes (неравномерное окрашивание по Граму, «метаногенные бактерии», теперь известные как Археи)
Молекулярная эра
«Архаические бактерии» и реклассификация Вёзе

Вёзе утверждал, что бактерии, археи и эукариоты представляют собой отдельные линии происхождения, которые на раннем этапе отошли от наследственной колонии организмов.[45][46] Однако некоторые биологи утверждают, что археи и эукариоты произошли от группы бактерий.[47] В любом случае считается, что вирусы и археи начали отношения примерно два миллиарда лет назад, и это совместная эволюция могли происходить между членами этих групп.[48] Возможно, что последним общим предком бактерий и архей был термофил, что повышает вероятность того, что более низкие температуры являются «экстремальной средой» в терминах архей, а организмы, живущие в более прохладной среде, появились только позже.[49] Поскольку археи и бактерии не более связаны друг с другом, чем с эукариотами, термин прокариотЕдинственное сохранившееся значение - «не эукариот», что ограничивает его ценность.[50]
С улучшенными методологиями стало ясно, что метаногенные бактерии сильно отличаются и (ошибочно) считаются реликтами древних бактерий.[51] таким образом Карл Вёзе, считающийся предшественником революции молекулярной филогении, определила три основные линии происхождения: Архебактерии, то Эубактерии, а Уркариоты, последний теперь представлен нуклеоцитоплазматическим компонентом Эукариоты.[52] Эти родословные были формализованы в Домене ранга (Regio на латыни), которые разделили жизнь на 3 области: Эукариоты, то Археи и Бактерии.[2]
Подразделения
В 1987 году Карл Вёзе разделил Эубактерии на 11 отделов по 16S рибосомная РНК (SSU) последовательности, которые с некоторыми дополнениями используются до сих пор.[53][54]
Оппозиция
Хотя трехдоменная система широко распространена,[55] некоторые авторы выступали против этого по разным причинам.
Один выдающийся ученый, выступающий против трехдоменной системы, - Томас Кавалье-Смит, который предложил Археи и Эукариоты (в Неомура ) происходят от грамположительных бактерий (Посибактерии), которые, в свою очередь, происходят от грамотрицательных бактерий (Негибактерии) на основе нескольких логических аргументов,[56][57] которые являются весьма спорными и обычно игнорируются сообществом молекулярной биологии (c.f. комментарии рецензентов,[57] например Эрик Баптесте является «агностиком» в отношении выводов) и часто не упоминается в обзорах (например.[58]) из-за субъективного характера сделанных предположений.[59]
Однако, несмотря на наличие множества статистически подтвержденных исследований по укоренению древа жизни между Бактерии и Неомура с помощью различных методов,[60] включая те, которые невосприимчивы к ускоренной эволюции, что, как утверждает Кавалье-Смит, является источником предполагаемой ошибки в молекулярных методах[56]- есть несколько исследований, в которых были сделаны разные выводы, некоторые из которых помещают корень в тип Фирмикуты с вложенными археями.[61][62][63]
Молекулярная таксономия Радхи Гупты, основанная на консервативных сигнатурных последовательностях белков, включает монофилетическую грамотрицательную кладу, монофилетическую грамположительную кладу и полифилетическую археоту, полученную из грамположительных.[64][65][66] Молекулярный анализ Хори и Осавы показал связь между метабактериями (= Archeota) и эукариотами.[67] Единственный кладистический анализ бактерий, основанный на классических доказательствах, в значительной степени подтверждает результаты Гупты (см. всеобъемлющая мега-систематика ).
Джеймс Лейк представил 2 основных устройства царства (Parkaryotae + эукариоты и эоциты + Karyotae) и предложил схему 5 основных царств (Eukaryota, Eocyta, Methanobacteria, Halobacteria и Eubacteria), основанную на структуре рибосом и схему 4 основных царств (Eukaryota, Eocyta, Метанобактерии, и Photocyta ), бактерии классифицируются по трем основным биохимическим нововведениям: фотосинтез (Photocyta), метаногенез (Метанобактерии) и серное дыхание (Eocyta ).[68][69][70] Он также обнаружил доказательства того, что грамотрицательные бактерии возникли в результате симбиоза двух грамположительных бактерий.[71]
Органы власти
Классификация - это группировка организмов в все более инклюзивные группы на основе филогении и фенотипа, а номенклатура - это применение формальных правил для наименования организмов.[72]
Номенклатурный орган
Несмотря на отсутствие официальной и полной классификации прокариот, названия (номенклатура), данные прокариотам, регулируются Международным кодексом номенклатуры бактерий (Бактериологический код ), книгу, содержащую общие соображения, принципы, правила и различные примечания, а также советы[73] аналогично номенклатурные коды других групп.
Классификационные органы
Таксоны, которые были правильно описаны, рассмотрены в Руководство Берджи по систематической бактериологии, который призван помочь в идентификации видов и считается высшим авторитетом.[42] Доступна онлайн-версия таксономического обзора бактерий и архей (ТОБА). [1].
Список названий прокариот, стоящих в номенклатуре (LPSN) - это онлайн-база данных, которая в настоящее время содержит более двух тысяч принятых имен с их ссылками, этимологией и различными примечаниями.[74]
Описание новых видов
В International Journal of Systematic Bacteriology / Международный журнал систематической и эволюционной микробиологии (IJSB / IJSEM) - это рецензируемый журнал, который действует как официальный международный форум для публикации новых таксонов прокариот. Если вид опубликован в другом рецензируемом журнале, автор может подать запрос в IJSEM с соответствующим описанием, которое, если оно верное, приведет к тому, что новый вид будет включен в Список валидации IJSEM.
Распределение
Коллекции микробных культур - это хранилища штаммов, целью которых является их сохранение и распространение. Основные из них:[72]
Анализирует
Эта секция нуждается в расширении. Вы можете помочь добавляя к этому. (Май 2011 г.) |
Сначала бактерии классифицировались исключительно на основании их формы (вибрионы, палочки, кокки и т. Д.), Наличия эндоспор, окраски по Граму, аэробных условий и подвижности. Эта система изменилась с изучением метаболических фенотипов, в которых использовались метаболические характеристики.[75] В последнее время, с появлением молекулярной филогении, несколько генов используются для идентификации видов, наиболее важным из которых является 16S рРНК ген, за которым следует 23S, ЕГО регион, gyrB и другие, чтобы подтвердить лучшее разрешение. Самый быстрый способ определить соответствие изолированного штамма виду или роду сегодня - это амплификация его гена 16S с универсальными праймерами, секвенирование ампликона размером 1,4 КБ и отправка его в специализированную базу данных идентификации в Интернете, а именно: Проект базы данных рибосом[2], которые выравнивают последовательность с другими последовательностями 16S, используя адское, глобальное выравнивание оснований вторичной структуры,[76][77] или же ARB SILVA, который выравнивает последовательности через SINA (инкрементный выравниватель SILVA), который выполняет локальное выравнивание семени и расширяет его [3].[78]
Существует несколько методов идентификации:[72]
- Фенотипические анализы
- анализы жирных кислот
- Условия роста (Чашка агара, Многолуночные планшеты Biolog )
- Генетический анализ
- ДНК-ДНК гибридизация
- ДНК-профилирование
- Последовательность
- Коэффициенты GC
- Филогенетические анализы
- 16S филогения на основе
- филогения на основе других генов
- Анализ мультигенной последовательности
- Анализ последовательности всего генома
Новые виды
Минимальные стандарты описания нового вида зависят от того, к какой группе принадлежит этот вид. c.f.[79]
Candidatus
Candidatus входит в состав таксономический название бактерии, которая не может содержаться в коллекции бактериологических культур. Это промежуточный таксономический статус некультивируемых организмов. например "Candidatus Pelagibacter ubique "
Понятие видов
Бактерии делятся бесполым путем и по большей части не проявляют регионализма ("Все везде "), поэтому концепция вида, которая лучше всего подходит для животных, становится полностью предметом суждения.
Количество названных видов бактерий и архей (около 13000)[80] удивительно мало, учитывая их раннюю эволюцию, генетическое разнообразие и место обитания во всех экосистемах. Причина тому - различия в представлениях о видах между бактерии и макроорганизмов, трудности выращивания / характеристики в чистой культуре (предварительное условие для определения новых видов, смотри выше) и обширный горизонтальный перенос генов стирание различия видов.[81]
Наиболее общепринятое определение - определение многофазных видов, которое учитывает как фенотипические, так и генетические различия.[82]Однако более быстрая диагностика для этого случая порог для разделения видов составляет менее 70% ДНК-ДНК-гибридизации,[83] что соответствует менее чем 97% идентичности последовательности ДНК 16S.[84] Было отмечено, что если бы это применялось к классификации животных, порядок приматы будет единым видом.[85]По этой причине были предложены более строгие определения видов на основе полных последовательностей генома.[86]
Патология против филогении
В идеале таксономическая классификация должна отражать эволюционную историю таксонов, то есть филогению. Хотя существуют некоторые исключения, когда фенотип различается в группе, особенно с медицинской точки зрения. Ниже приведены некоторые примеры проблемных классификаций.
кишечная палочка: слишком большой и полифилетический
В семье Энтеробактерии класса Гаммапротеобактерии, виды в роде Шигелла (С. dysenteriae, S. flexneri, С. бойди, S. sonnei ) с эволюционной точки зрения являются штаммами вида кишечная палочка (полифилетический), но из-за генетических различий вызывают различные заболевания в случае патогенных штаммов.[87] кишечная палочка - это плохо классифицируемый вид, так как некоторые штаммы имеют только 20% общего генома. Поскольку он настолько разнообразен, ему следует дать более высокий таксономический рейтинг. Однако из-за медицинских условий, связанных с этим видом, он не будет изменен, чтобы избежать путаницы в медицинском контексте.
Bacillus cereus группа: тесная и полифилетическая
Аналогичным образом Бациллы виды (= тип Фирмикуты ) принадлежащих к "B. cereus группа" (B. anthracis, B. cereus, Б. thuringiensis, B. mycoides, Б. pseudomycoides, Б. weihenstephanensis и Б. медуза ) имеют 99-100% схожую последовательность 16S рРНК (97% - это обычно цитируемый адекватный видовой предел) и являются полифилетическими, но по медицинским показаниям (сибирская язва и Т. Д.) остаются отдельными.[88]
Yersinia pestis: очень недавний вид
Yersinia pestis в действительности является напряжением Иерсиний псевдотуберкулез, но с островом патогенности, который вызывает совершенно иную патологию (черная чума и симптомы, подобные туберкулезу соответственно), которая возникла от 15 000 до 20 000 лет назад.[89]
Вложенные роды в Псевдомонады
В порядке гаммапротеобактерий Pseudomonadales, род Азотобактер и виды Azomonas macrocytogenes на самом деле являются членами рода Псевдомонады, но были неправильно классифицированы из-за способности фиксировать азот и большого размера рода Псевдомонады что делает классификацию проблематичной.[75][90][91] Вероятно, это будет исправлено в ближайшем будущем.
Вложенные роды в Бациллы
Другой пример большого рода с вложенными родами - это род Бациллы, в котором роды Paenibacillus и Brevibacillus являются вложенными кладами.[92] В настоящее время недостаточно геномных данных для полного и эффективного исправления таксономических ошибок в Бациллы.
Агробактерии: сопротивление смене имени
На основе молекулярных данных показано, что род Агробактерии вложен в Ризобий и Агробактерии виды переведены в род Ризобий (в результате получается следующий comp. nov .: Rhizobium radiobacter (ранее известный как A. tumefaciens), R. rhizogenes, Р. Руби, R. undicola и R. vitis)[93] Учитывая патогенную природу растений Агробактерии вида, было предложено сохранить род Агробактерии[94] и последнее было опровергнуто[95]
Номенклатура
Таксономические названия написаны курсивом (или подчеркнуты, если написаны от руки) с большой первой буквой, за исключением эпитетов для видов и подвидов. Несмотря на то, что это распространено в зоологии, тавтонимы (например. Бизон бизон) являются неприемлемыми, а названия таксонов, используемых в зоологии, ботанике или микологии, не могут быть повторно использованы для бактерий (ботаника и зоология имеют общие названия).
Номенклатура - это набор правил и условностей, регулирующих названия таксонов. Разница в номенклатуре между различными царствами / доменами рассматривается в.[96]
Для бактерий допустимые имена должны иметь латинское или неолатинское имя и могут использовать только основные латинские буквы (w и j включительно, см. История латинского алфавита для них), следовательно, дефисы, акценты и другие буквы не принимаются и должны быть правильно транслитерированы (например, ß = ss).[97] Древнегреческий язык, написанный греческим алфавитом, должен быть транслитерированный в латинский алфавит.
Когда составные слова создаются, соединяющая гласная необходима в зависимости от происхождения предыдущего слова, независимо от следующего слова, если последнее не начинается с гласной, и в этом случае соединительная гласная не добавляется. Если первое соединение латинское, то соединительная гласная будет -i-, тогда как если первое соединение греческое, соединительная гласная будет -o-.[98]
По этимологиям имен обратитесь к LPSN.
Правила для высших таксонов
Для Прокариоты (Бактерии и Археи) ранг королевства не используется[99] (хотя некоторые авторы называют филы царствами[72])
Если новый или измененный вид помещается в новые ранги, в соответствии с Правилом 9 Бактериологического кодекса название формируется путем добавления соответствующего суффикса к основе названия типового рода.[73] Для подкласса и класса рекомендация от[100] обычно следует, что приводит к нейтральному множественному числу, однако некоторые имена не следуют за ним и вместо этого учитывают греко-латинскую грамматику (например, женское множественное число Термотоги, Водные и Хламидии, мужское множественное число Хлорофлекси, Бациллы и Дейнококки и греческое множественное число Спирохеты, Gemmatimonadetes и Chrysiogentes ).[101]
| Классифицировать | Суффикс | Пример |
|---|---|---|
| Род | -ae | (Elusimicrobiae) |
| Subtribe (не используется) | -inae | (Elusimicrobiinae) |
| Племя (вышедшее из употребления) | -eae | (Elusimicrobiieae) |
| Подсемейство | -oideae | (Elusimicrobioideae) |
| Семья | -aceae | Elusimicrobiaceae |
| Подотряд | -ineae | (Elusimicrobineae) |
| Заказ | -продажи | Elusimicrobiales |
| Подкласс | -idae | (Elusimicrobidae) |
| Учебный класс | -я | Elusimicrobia |
| Тип | см текст | Elusimicrobia |
Окончания Phyla
Типы не подпадают под Бактериологический кодекс,[101] однако научное сообщество обычно следует Ncbi и Lpsn таксономия, где название типа обычно является множественным числом от типового рода, за исключением Фирмикуты, Цианобактерии и Протеобактерии, чьи имена не происходят от названия рода. Высшие таксоны, предложенные Кавалер-Смит[56] обычно игнорируются сообществом молекулярной филогении (например.[58]) (смотри выше ).
Для Археи суффикс -археота используется.[102] За бактериальный тип было предложено, чтобы суффикс -бактерии использоваться для типов.[103]
Следовательно, для основного типа название типа такое же, как у первого описанного класса:
- Ацидобактерии (из Ацидобактерии )
- Актинобактерии (из Актиномицеты )
- Caldisericia (из Caldisericum )
- Elusimicrobia (из Elusimicrobium )
- Фузобактерии (из Фузобактерии )
- Термодесульфобактерии (из Термодесульфобактерии )
- Термотоги (из Thermotoga )
- Водные (из Aquifex )
- Хламидии (из Хламидиоз )
- Хлорофлекси (из Хлорофлексус )
- Chrysiogentes (из Хризиоген )
- Gemmatimonadetes (из Гемматимонас )
- Deferribacteres (из Deferribacter )
В то время как для других, где суффикс -ia для класса используется независимо от грамматики, они различаются:
- филюм Bacteroidetes против класса Бактероидии из Бактероиды
- филюм Хлороби против класса Хлоробия из Хлоробий
- филюм Веррукомикробия против класса Веррукомикробии из Веррукомикробий (аномальное название класса)
- филюм Диктиогломи против класса Диктиогломия из Диктиогломус
- филюм Фибробактерии против класса Фибробактерии из Фибробактер (ср. суффикс -бактер обратите внимание на разницу с Deferribacteres )
- филюм Lentisphaerae против класса Лентисфаерия из Lentisphaera
- филюм Нитроспира или же Нитроспиры против класса Нитроспира из Нитроспира
- филюм Спирохеты против класса Спирохеты из Спирохета
- филюм Synergistetes против класса Синергистетия из Synergistes
- филюм Планктомицеты против Planctomycea из Planctomyces
Исключение составляет тип Дейнококк – Термус, который несет пару родов через дефис - только латинские буквы без ударения принимаются для правильных имен, но типы официально не признаются.[102]Совсем недавно было предложено внести поправки в Бактериологический код указать -aeota в качестве окончания для бактериального типа и чтобы имена были производными от класса типа внутри этого типа.[104] Для этого потребуются следующие изменения:
- Ацидобактерии → Acidobacteraeota
- Актинобактерии → Actinobacteraeota
- Протеобактерии → Alphaproteobacteraeota
- Водные → Aquificaeota
- Арматимонадеты → Armatimonadaeota
- Фирмикуты → Bacillaeota
- Bacteroidetes → Bacteroidaeota
- Caldiserica → Caldisericaeota
- Хламидии → Chlamydaeota
- Хлороби → Хлоробаеота
- Хлорофлекси → Хлорофлексаеота
- Chrysiogentes → Chrysiogenaeota
- Deferribacteres → Deferribacteraeota
- Deinococcus-Thermus → Deinococcaeota
- Диктиогломи → Диктиогломеота
- Elusimicrobia → Elusimicrobaeota
- Фибробактерии → Fibrobacteraeota
- Фузобактерии → Fusobacteraeota
- Gemmatimonadetes → Gemmatimonadaeota
- Игнавибактерии → Игнавибактераэота
- Lentisphaerae → Lentisphaeraeota
- Euryarchaeota → Methanobacteraeota
- Tenericutes → Mollicutaeota
- Таумархеота → Nitrososphaeraeota
- Нитроспира → Нитроспиреота
- Планктомицеты → Planctomycetaeota
- Спирохеты → Спирохеты
- Synergistetes → Synergistaeota
- Термодесульфобактерии → Thermodesulfobacteraeota
- Термомикробия → Термомикробаэота
- Crenarchaeota → Thermoproteaeota
- Термотоги → Thermotogaeota
- Веррукомикробия → Веррукомикробаэота
Имена в честь людей
Некоторые виды названы в честь людей, например, первооткрывателя или известного человека в области микробиологии. Сальмонелла после D.E. Лосось, который его открыл (хотя и как «Bacillus typhi»[105]).[106]
В случае родового эпитета все имена, производные от людей, должны быть в женском именительном падеже, путем изменения окончания на -a или уменьшительного -ella, в зависимости от имени.[98]
Для конкретного эпитета имена могут быть преобразованы в форму прилагательного (добавление -nus (m.), -Na (f.), -Num (n.) В соответствии с полом названия рода) или в родительный падеж родительного падежа. латинизированное имя.[98]
Имена после мест
Многие виды (специфический эпитет) названы в честь места, где они присутствуют или встречаются (например, Thiospirillum jenense ). Их имена создаются путем образования прилагательного путем соединения названия местности с окончанием -ensis (m. Или f.) Или ense (n.) В соответствии с полом названия рода, если для места не существует классического латинского прилагательного. . Однако названия мест не следует употреблять как существительные в родительном падеже.[98]
Народные имена
Несмотря на то, что некоторые гетеро / гомогенные колонии или биопленки бактерий имеют названия на английском языке (например, зубной налет или же Звездное желе ), ни один вид бактерий не имеет общеупотребительного / банального / общепринятого названия на английском языке.
Для имен в единственном числе нельзя использовать множественное число (singulare tantum ), что подразумевает наличие нескольких групп с одним и тем же ярлыком, а не нескольких членов этой группы (по аналогии, в английском языке стулья и столы - это типы мебели, которые нельзя использовать во множественном числе «мебель» для описания обоих членов), и наоборот имена во множественном числе - тантум множественного числа. Тем не менее, частичное исключение из этого составляет использование местных названий. Однако, чтобы избежать повторения таксономических названий, которые нарушают поток прозы, часто используются и рекомендуются местные названия представителей рода или более высоких таксонов, они образуются написав название таксона в нижнем регистре предложения ("стандартный" в MS Office), таким образом, обрабатывая имя собственное как английский имя нарицательное (например, сальмонеллы), хотя есть некоторые споры о грамматике множественного числа, которое может быть либо правильным множественным числом, добавив - (e) s (сальмонеллы), либо с использованием древнегреческой или латинской формы множественного числа (неправильные множественные числа ) существительного (сальмонеллы); последнее проблематично, поскольку множественное число - бактерия будет -bacteres, тогда как множественное число myces (N.L. masc. n. от греч. masc. n. mukes) будет mycetes.[107]
Обычаи присутствуют для определенных имен, например, тех, которые заканчиваются на -монас преобразуются в -монаду (одну псевдомонаду, две аэромонады и немонады).
Бактерии, являющиеся этиологическая причина болезни часто обозначаются названием болезни, за которым следует описывающее существительное (бактерия, бацилла, кокк, агент или название их типа), например бактерия холеры (Холерный вибрион ) или спирохеты болезни Лайма (Borrelia burgdorferi ), обратите внимание также на риккетсиоз (Риккетсия акари ) (подробнее см.[108]).
Трепонема превращается в трепонему, а множественное число - трепонемы, а не трепонемы.
У некоторых необычных бактерий есть особые названия, например, овал Куина (Quinella ovalis ) и квадрат Уолсби (Haloquadratum walsbyi ).
До появления молекулярной филогении многие более высокие таксономические группы имели только тривиальные названия, которые используются до сих пор, некоторые из которых являются полифилетическими, например Rhizobacteria. Некоторые более высокие таксономические тривиальные имена:
- Сине-зеленые водоросли являются членами филума Цианобактерии
- Зеленые бактерии, не содержащие серы, являются членами филума Хлорофлекси
- Зеленые серные бактерии являются членами Хлороби
- Пурпурные бактерии - это некоторые, но не все, члены этого типа. Протеобактерии
- Пурпурные серные бактерии являются членами отряда Chromatiales
- грамположительные бактерии с низким содержанием G + C являются членами филума Фирмикуты, независимо от содержания GC
- грамположительные бактерии с высоким содержанием G + C являются членами филума Актинобактерии, независимо от содержания GC
- Ризобактерии являются представителями различных родов протеобактерий.
- Ризобии являются членами отряда Ризобиальные
- Молочнокислые стрептококки являются представителями рода Лактококк
- Коринеформные бактерии являются членами семейства Коринебактерии
- Плодоносящие планирующие бактерии или миксобактерии являются членами отряда Myxococcales
- Энтерии являются членами ордена Энтеробактерии, хотя этого термина следует избегать, если они не живут в кишечнике, например Пектобактерии
- Бактерии уксусной кислоты являются членами семейства Acetobacteraceae
Терминология
- Сокращенное обозначение вида sp. (множественное число виды) и используется после родового эпитета для обозначения вида этого рода. Часто используется для обозначения штамма рода, для которого вид неизвестен, либо потому, что этот организм еще не был описан как вид, либо было проведено недостаточно тестов для его идентификации. Например Halomonas sp. GFAJ-1
- Если бактерия известна и хорошо изучена, но не подлежит культивированию, в ее названии используется термин Candidatus.
- Базоним - это исходное название новой комбинации, а именно первое название, данное таксону перед его переклассификацией.
- Синоним - это альтернативное название таксона, т.е. таксон дважды ошибочно описан
- Когда таксон переносится, он становится новой комбинацией (comb. Nov.) Или nomina nova (nom. Nov.)
- парафилия, монофилия и полифилия
Смотрите также
- Список латинских и греческих слов, обычно используемых в систематических именах
- Список родов бактерий
- Список бактериальных заказов
- Список секвенированных геномов прокариот
- Проект человеческого микробиома
- Микробная экология
- Список клинически важных бактерий
- Список секвенированных геномов архей
- Список родов архей
- Видовая проблема
- Эволюционный класс
- Комплекс загадочных видов
- Синоним (таксономия)
- Таксономия
- LPSN, список допустимых названий бактерий и архей
- Цианобактерии, тип обычных бактерий, но плохо классифицированных в настоящее время
- Бактериальный тип, сложная классификация
Рекомендации
- ^ Линней, Карл (1735). Systemae Naturae, sive regna tria naturae, систематика предложений по классам, порядкам, родам и видам.
- ^ а б Woese, C. R .; Kandler, O .; Уилис, М. Л. (1990). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев». Труды Национальной академии наук. 87 (12): 4576–4579. Bibcode:1990PNAS ... 87,4576 Вт. Дои:10.1073 / pnas.87.12.4576. ЧВК 54159. PMID 2112744.
- ^ Портер-младший (1976). «Энтони ван Левенгук: трехсотлетие открытия бактерий». Бактериологические обзоры. 40 (2): 260–9. Дои:10.1128 / ммбр.40.2.260-269.1976. ЧВК 413956. PMID 786250.
- ^ ван Левенгук А (1684). "Отрывок из письма мистера Энтони Леввенгука из Делфта от 17 сентября 1683 г., содержащего некоторые микроскопические наблюдения, о животных в зубном налете, о червях в носу, о кутикуле, состоящей из чешуек. ". Философские труды. 14 (155–166): 568–574. Дои:10.1098 / рстл.1684.0030.
- ^ ван Левенгук А (1700 г.). "Часть письма г-на Антония ван Левенгука о червях в печени овец, о комарах и животных в экскрементах лягушек". Философские труды. 22 (260–276): 509–518. Bibcode:1700РСПТ ... 22..509В. Дои:10.1098 / рстл.1700.0013.
- ^ ван Левенгук А (1702 г.). «Часть письма г-на Энтони ван Левенгука, Ф. Р. С. о зеленых сорняках, растущих в воде, и некоторых животных, обнаруженных вокруг них». Философские труды. 23 (277–288): 1304–11. Bibcode:1702РСПТ ... 23,1304В. Дои:10.1098 / рстл.1702.0042. S2CID 186209549.
- ^ а б c Мюррей, Р.Г.Е., Холт, Дж. (2005). История Руководство Берджи. В: Гаррити, Г.М., Бун, Д.Р. И Кастенхольц, Р.В. (ред., 2001). Руководство Берджи по систематической бактериологии, 2-е изд., Т. 1, Springer-Verlag, New York, p. 1-14. связь. [См. Стр. 2.]
- ^ Пот Б., Гиллис М. и Де Лей Дж. Род Aquaspirillum. В: Balows, A., Trüper, H.G., Dworkin, M., Harder, W., Schleifer, K.-H. (Ред.). Прокариоты, 2-е изд., Т. 3. Springer-Verlag. Нью-Йорк. 1991 г.
- ^ "Этимология слова" бактерии """. Интернет-словарь этимологии. В архиве из оригинала 18 ноября 2006 г.. Получено 23 ноября 2006.
- ^ а б c d Геккель, Эрнст (1867). Generelle Morphologie der Organismen (на немецком). Берлин: Реймер. ISBN 978-1-144-00186-3 - через Интернет-архив.
- ^ а б Фон Нэгели, К. (1857). Каспари, Р. (ред.). "Bericht über die Verhandlungen der 33. Versammlung deutscher Naturforscher und Aerzte, gehalten в Бонне от 18 до 24 сентября 1857 г." [Отчет о переговорах на 33-м совещании немецких естествоиспытателей и врачей, состоявшемся в Бонне 18-24 сентября 1857 г.]. Botanische Zeitung (на немецком). 15: 749–776 - через Интернет-архив.
- ^ Пачини, Ф .: Osservazione microscopiche e deduzioni patologiche sul cholera asiatico. Gazette Medicale de Italiana Toscano Firenze, 1854, 6, 405-412.
- ^ а б Эренберг, К. Г. (1835). "Dritter Beitrag zur Erkenntniss grosser Organization in der Richtung des kleinsten Raumes". Physikalische Abhandlungen der Koeniglichen Akademie der Wissenschaften zu Berlin aus den Jahren 1833-1835С. 143-336.
- ^ Эренберг, К. Г. (1830). "Beiträge zur Kenntnis der Organization der Infusorien und ihrer geographischen Verbreitung besonders в Сибири". Abhandlungen der Koniglichen Akademie der Wissenschaften zu Berlin 1832 г., стр. 1–88.
- ^ Вход Protomonas в LPSN; Euzéby, J.P. (1997). «Список названий бактерий с позиции в номенклатуре: папка, доступная в Интернете». Международный журнал систематической и эволюционной микробиологии. 47 (2): 590–2. Дои:10.1099/00207713-47-2-590. PMID 9103655.
- ^ Чаттон, Э. (1925). "Панспорелла perplexa: Réflexions sur la biologie et la phylogénie des protozoaires ". Анналы естественных наук. 10 (VII): 1–84.
- ^ Смит, Эрвин Ф. (1905). Номенклатура и классификация бактерий по отношению к болезням растений, Vol. 1.
- ^ Вюйлемен (1913). «Рода Schizomycetum». Annales Mycologici, Vol. 11. С. 512–527.
- ^ Кон, Фердинанд (1875). "Untersuchungen uber Bakterien, II". Beitrage zur Biologie der Pfanzen, Vol. 1. С. 141–207.
- ^ Станье и ван Нил (1941). «Основные положения бактериальной классификации». Журнал бактериологии, Vol. 42. С. 437–466.
- ^ Биссет (1962). Бактерии2-е изд .; Лондон: Ливингстон.
- ^ Мигула, Уолтер (1897). System der Bakterien. Йена, Германия: Густав Фишер.
- ^ Орла-Йенсен (1909). "Die Hauptlinien des naturalischen Bakteriensystems nebst einer Ubersicht der Garungsphenomene". Центр. Бакт. Паразитенк., Vol. II, № 22, стр. 305–346
- ^ Руководство Берджи по детерминантной бактериологии (1925). Балтимор: Williams & Wilkins Co. (со многими последующими изданиями).
- ^ Бьюкенен, Р. Э. (1916). «Исследования по номенклатуре и классификации бактерий: проблема бактериальной номенклатуры». Журнал бактериологии. 1 (6): 591–596. Дои:10.1128 / jb.1.6.591-596.1916. ЧВК 378679. PMID 16558720 - через Национальный центр биотехнологической информации.
- ^ Кон, Фердинанд (1872 г.). "Организаторы Покенлимфе". Вирхов (ред.), Архив, Vol. 55, стр. 237.
- ^ Кон, Фердинанд (1872 б) "Untersuchungen ilber Bakterien". Beitrage zur Biologie der Pflanzen Vol. 1, № 1, с. 136.
- ^ Detoni, J. B .; Тревизан, В. (1889). «Schizomycetaceae». Саккардо (ред.), Sylloge Fungorum, Vol. 8, стр. 923.
- ^ Бьюкенен, Р. Э. (март 1917 г.). «Исследования по номенклатуре и классификации бактерий: II. Основные подразделения шизомицетов». Журнал бактериологии. 2 (2): 155–64. Дои:10.1128 / JB.2.2.155-164.1917. ЧВК 378699. PMID 16558735.
- ^ «Международный кодекс бактериологической номенклатуры». Журнал бактериологии. 55 (3): 287–306. Март 1948 г. Дои:10.1128 / JB.55.3.287-306.1948. ЧВК 518444. PMID 16561459.
- ^ Прево, А. Р. (1958). Мануэль де классификации и определения анаэробных бакетов. Париж: Массон.
- ^ Прево, А. Р. (1963). Бактерии в Précis de Botanique. Париж: Массон.
- ^ Линней, К. (1735). Systemae Naturae, sive regna tria naturae, систематика предложений по классам, порядкам, родам и видам.
- ^ Геккель, Э. (1866). Generelle Morphologie der Organismen. Реймер, Берлин.
- ^ Чаттон, Э. (1925). "Панспорелла perplexa. Réflexions sur la biologie et la phylogénie des protozoaires ". Annales des Sciences Naturelles - Zoologie et Biologie Animale. 10-VII: 1–84.
- ^ Коупленд, Х. (1938). «Царства организмов». Ежеквартальный обзор биологии. 13: 383–420. Дои:10.1086/394568.
- ^ Уиттакер, Р. Х. (январь 1969 г.). «Новые представления о царствах организмов». Наука. 163 (3863): 150–60. Bibcode:1969Sci ... 163..150W. Дои:10.1126 / science.163.3863.150. PMID 5762760.
- ^ Woese, C .; Kandler, O .; Уилис, М. (1990). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев». Труды Национальной академии наук Соединенных Штатов Америки. 87 (12): 4576–9. Bibcode:1990PNAS ... 87,4576 Вт. Дои:10.1073 / pnas.87.12.4576. ЧВК 54159. PMID 2112744.
- ^ Кавальер-Смит, Т. (1998). «Пересмотренная система жизни шести царств». Биологические обзоры. 73 (03): 203–66. Дои:10.1111 / j.1469-185X.1998.tb00030.x. PMID 9809012.
- ^ Ruggiero, Michael A .; Гордон, Деннис П .; Оррелл, Томас М .; Байи, Николас; Бургуэн, Тьерри; Бруска, Ричард С .; Кавальер-Смит, Томас; Guiry, Michael D .; Кирк, Пол М .; Туэзен, Эрик В. (2015). «Классификация всех живых организмов более высокого уровня». PLOS ONE. 10 (4): e0119248. Bibcode:2015PLoSO..1019248R. Дои:10.1371 / journal.pone.0119248. ЧВК 4418965. PMID 25923521.
- ^ GIBBONS N.E .; МЮРРЕЙ Р.Г.Е. (1978). «Предложения по высшим таксонам бактерий». Международный журнал систематической бактериологии. 28: 1–6. Дои:10.1099/00207713-28-1-1.
- ^ а б Джордж М. Гаррити: Руководство Берджи по систематической бактериологии. 2. Auflage. Springer, Нью-Йорк, 2005 г., том 2: Proteobacteria, часть B: Gammaproteobacteria
- ^ МЮРРЕЙ (R.G.E.): Высшие таксоны или место для всего ...? В: Н. KRIEG и J.G. Холт (ред.) Bergey's Manual of Systematic Bacteriology, vol. 1, The Williams & Wilkins Co., Балтимор, 1984, стр. 31–34
- ^ Ciccarelli FD, Doerks T., von Mering C, Creevey CJ, Snel B, Bork P (2006). «К автоматической реконструкции дерева жизни с высоким разрешением». Наука. 311 (5765): 1283–7. Bibcode:2006Научный ... 311.1283C. CiteSeerX 10.1.1.381.9514. Дои:10.1126 / science.1123061. PMID 16513982. S2CID 1615592.
- ^ Woese CR, Gupta R (1981). «Архебактерии - просто производные от прокариотов?». Природа. 289 (5793): 95–6. Bibcode:1981Натура 289 ... 95Вт. Дои:10.1038 / 289095a0. PMID 6161309. S2CID 4343245.
- ^ Woese C (1998). «Вселенский предок». Труды Национальной академии наук Соединенных Штатов Америки. 95 (12): 6854–9. Bibcode:1998PNAS ... 95,6854 Вт. Дои:10.1073 / пнас.95.12.6854. ЧВК 22660. PMID 9618502.
- ^ Гупта Р.С. (2000). «Естественные эволюционные отношения между прокариотами». Крит. Rev. Microbiol. 26 (2): 111–31. CiteSeerX 10.1.1.496.1356. Дои:10.1080/10408410091154219. PMID 10890353. S2CID 30541897.
- ^ С. Майкл Хоган. 2010 г. Вирус. Энциклопедия Земли. Редакторы: Катлер Кливленд и Сидни Драгган
- ^ Грибальдо С., Брошье-Армане С. (2006). «Происхождение и эволюция архей: современное состояние». Философские труды Королевского общества B. 361 (1470): 1007–22. Дои:10.1098 / rstb.2006.1841. ЧВК 1578729. PMID 16754611.
- ^ Woese CR (1 марта 1994 г.). «Где-то должен быть прокариот: микробиология ищет себя». Microbiol. Rev. 58 (1): 1–9. Дои:10.1128 / MMBR.58.1.1-9.1994. ЧВК 372949. PMID 8177167.
- ^ Balch, W .; Magrum, L .; Fox, G .; Wolfe, R .; Woese, C. (1977). «Древнее расхождение среди бактерий». Журнал молекулярной эволюции. 9 (4): 305–311. Bibcode:1977JMolE ... 9..305B. Дои:10.1007 / BF01796092. PMID 408502. S2CID 27788891.
- ^ Francolini, C .; Эгет, Х. (1979). «Перцепционная селективность зависит от задачи: всплывает эффект всплывающего окна». Восприятие и психофизика. 25 (2): 99–110. Дои:10.3758 / bf03198793. PMID 432104.
- ^ Холланд Л. (22 мая 1990 г.). «Везе, Карл в авангарде революции бактериальной эволюции». Ученый. 4 (10).
- ^ Woese, C. R. (1987). «Бактериальная эволюция». Микробиологические обзоры. 51 (2): 221–271. Дои:10.1128 / MMBR.51.2.221-271.1987. ЧВК 373105. PMID 2439888.
- ^ Уорд, Б. Б. (2002). "Сколько существует видов прокариот?". Труды Национальной академии наук. 99 (16): 10234–10236. Bibcode:2002PNAS ... 9910234W. Дои:10.1073 / pnas.162359199. ЧВК 124894. PMID 12149517.
- ^ а б c Кавальер-Смит, Т. (2002). «Неомуранское происхождение архебактерий, негибактериальный корень универсального дерева и бактериальная мегаклассификация». Международный журнал систематической и эволюционной микробиологии. 52 (Чт 1): 7–76. Дои:10.1099/00207713-52-1-7. PMID 11837318.
- ^ а б Кавальер-Смит, Т. (2006). «Укоренение древа жизни с помощью анализа переходов». Биология Директ. 1: 19. Дои:10.1186/1745-6150-1-19. ЧВК 1586193. PMID 16834776.
- ^ а б Пейс, Н. Р. (2009). «Картирование Древа Жизни: прогресс и перспективы». Обзоры микробиологии и молекулярной биологии. 73 (4): 565–576. Дои:10.1128 / MMBR.00033-09. ЧВК 2786576. PMID 19946133.
- ^ Fournier, G.P .; Гогартен, Дж. П. (2010). «Укоренение рибосомного древа жизни». Молекулярная биология и эволюция. 27 (8): 1792–1801. Дои:10.1093 / molbev / msq057. PMID 20194428.
- ^ Fournier, G.P .; Гогартен, Дж. П. (2010). «Укоренение рибосомного древа жизни». Молекулярная биология и эволюция. 27 (8): 1792–1801. Дои:10.1093 / molbev / msq057. PMID 20194428.
- ^ Lake, J .; Servin, J .; Herbold, C .; Скопхаммер, Р. (2008). «Свидетельства нового корня древа жизни». Систематическая биология. 57 (6): 835–843. Дои:10.1080/10635150802555933. PMID 19085327.
- ^ Skophammer, R.G .; Сервин, Дж. А .; Herbold, C.W .; Лейк, Дж. А. (2007). «Доказательства наличия грамположительных эубактериальных корней древа жизни». Молекулярная биология и эволюция. 24 (8): 1761–1768. Дои:10.1093 / молбев / msm096. PMID 17513883.
- ^ Valas, R.E .; Борн, П. Э. (2011). «Происхождение производного суперсарства: как грамположительная бактерия пересекла пустыню, чтобы стать археоном». Биология Директ. 6: 16. Дои:10.1186/1745-6150-6-16. ЧВК 3056875. PMID 21356104.
- ^ Гупта, Р. С. (1998). «Филогения белков и сигнатурные последовательности: переоценка эволюционных взаимоотношений между архебактериями, эубактериями и эукариотами». Обзоры микробиологии и молекулярной биологии. 62 (4): 1435–1491. Дои:10.1128 / MMBR.62.4.1435-1491.1998. ЧВК 98952. PMID 9841678.
- ^ Гупта, Р. С. (1998). «Третья область жизни (археи): установленный факт или парадигма, находящаяся под угрозой исчезновения?». Теоретическая популяционная биология. 54 (2): 91–104. Дои:10.1006 / tpbi.1998.1376. PMID 9733652.
- ^ Гупта, Р. С. (2000). «Естественные эволюционные отношения между прокариотами». Критические обзоры в микробиологии. 26 (2): 111–131. CiteSeerX 10.1.1.496.1356. Дои:10.1080/10408410091154219. PMID 10890353. S2CID 30541897.
- ^ Hori, H .; Осава, С. (1987). «Происхождение и эволюция организмов на основе последовательностей 5S рибосомной РНК». Молекулярная биология и эволюция. 4 (5): 445–472. Дои:10.1093 / oxfordjournals.molbev.a040455. PMID 2452957.
- ^ Лейк, Дж. А. (1986). «Альтернатива архебактериальной догме». Природа. 319 (6055): 626. Bibcode:1986Натура.319..626л. Дои:10.1038 / 319626b0. S2CID 4273843.
- ^ Lake, JA; Хендерсон, Э; Оукс, М; Кларк, MW (1984). «Эоциты: новая структура рибосом указывает на царство, тесно связанное с эукариотами». Труды Национальной академии наук Соединенных Штатов Америки. 81 (12): 3786–3790. Bibcode:1984PNAS ... 81.3786L. Дои:10.1073 / pnas.81.12.3786. ЧВК 345305. PMID 6587394.
- ^ Lake, J. A .; Хендерсон, Э .; Clark, M. W .; Scheinman, A .; Оукс, М. И. (1986). «Картирование эволюции с трехмерной структурой рибосом». Систематическая и прикладная микробиология. 7: 131–136. Дои:10.1016 / S0723-2020 (86) 80135-7.
- ^ Лейк, Дж. (2009). «Доказательства раннего прокариотического эндосимбиоза». Природа. 460 (7258): 967–971. Bibcode:2009Натурал.460..967л. Дои:10.1038 / природа08183. PMID 19693078. S2CID 4413304.
- ^ а б c d Мэдиган, Майкл (2009). Биология микроорганизмов Брока. Сан-Франциско: Пирсон / Бенджамин Каммингс. ISBN 978-0-13-232460-1.
- ^ а б Lapage, S .; Sneath, P .; Lessel, E .; Скерман, В .; Seeliger, H .; Кларк, В. (1992). Международный кодекс номенклатуры бактерий: Бактериологический кодекс, редакция 1990 г.. Вашингтон, округ Колумбия: ASM Press. PMID 21089234.
- ^ Вступительная запись в LPSN; Euzéby, J.P. (1997). «Список названий бактерий с позиции в номенклатуре: папка, доступная в Интернете». Международный журнал систематической и эволюционной микробиологии. 47 (2): 590–2. Дои:10.1099/00207713-47-2-590. PMID 9103655.
- ^ а б Паллерони, Н. Дж. (2010). «История псевдомонады». Экологическая микробиология. 12 (6): 1377–1383. Дои:10.1111 / j.1462-2920.2009.02041.x. PMID 20553550.
- ^ Olsen, G .; Larsen, N .; Woese, C. (1991). «Проект базы данных рибосомных РНК». Исследования нуклеиновых кислот. 19 Приложение: 2017–2021. Дои:10.1093 / nar / 19.suppl.2017. ЧВК 331344. PMID 2041798.
- ^ Cole, J. R .; Wang, Q .; Cardenas, E .; Fish, J .; Chai, B .; Farris, R.J .; Kulam-Syed-Mohideen, A. S .; McGarrell, D.M .; Марш, Т .; Гаррити, Г. М .; Тидже, Дж. М. (2009). «Проект базы данных рибосом: улучшенное выравнивание и новые инструменты для анализа рРНК». Исследования нуклеиновых кислот. 37 (Проблема с базой данных): D141 – D145. Дои:10.1093 / nar / gkn879. ЧВК 2686447. PMID 19004872.
- ^ Pruesse, E .; Quast, C .; Knittel, K .; Fuchs, B.M .; Ludwig, W .; Peplies, J .; Глокнер, Ф. О. (2007). «SILVA: всеобъемлющий онлайн-ресурс для проверенных и согласованных данных о последовательностях рибосомных РНК, совместимых с ARB». Исследования нуклеиновых кислот. 35 (21): 7188–7196. Дои:10.1093 / нар / гкм864. ЧВК 2175337. PMID 17947321.
- ^ минимальный ввод стандартов в LPSN; Euzéby, J.P. (1997). «Список названий бактерий с позиции в номенклатуре: папка, доступная в Интернете». Международный журнал систематической и эволюционной микробиологии. 47 (2): 590–2. Дои:10.1099/00207713-47-2-590. PMID 9103655.
- ^ «Количество опубликованных имен».
- ^ Стейли, Дж. Т. (2006). «Дилемма бактериального вида и концепция геномно-филогенетического вида». Философские труды Королевского общества B: биологические науки. 361 (1475): 1899–1909. Дои:10.1098 / rstb.2006.1914. ЧВК 1857736. PMID 17062409.
- ^ Геверс, Д .; Cohan, F.M .; Lawrence, J. G .; Spratt, B.G .; Coenye, T .; Feil, E. J .; Stackebrandt, E .; Van De Peer, Y. V .; Vandamme, P .; Томпсон, Ф. Л .; Качели, Дж. (2005). «Мнение: Переоценка прокариотических видов». Обзоры природы Микробиология. 3 (9): 733–739. Дои:10.1038 / nrmicro1236. PMID 16138101. S2CID 41706247.
- ^ Уэйн, Л. (1988). «Международный комитет по систематической бактериологии: Объявление отчета специального комитета по согласованию подходов к бактериальной систематике». Zentralblatt für Bakteriologie, Mikrobiologie und Hygiene. Серия A: Медицинская микробиология, инфекционные болезни, вирусология, паразитология. 268 (4): 433–434. Дои:10.1016 / s0176-6724 (88) 80120-2. PMID 3213314.
- ^ Stackebrandt, E; Гебель, Б. (1994). «Таксономическое примечание: место для реассоциации ДНК-ДНК и анализа последовательности 16S рДНК в настоящем определении видов в бактериологии». Int. J. Syst. Бактериол. 44 (4): 846–849. Дои:10.1099/00207713-44-4-846.
- ^ Стейли, Дж. (1997).«Биоразнообразие: угрожают ли виды микробов?». Текущее мнение в области биотехнологии. 8 (3): 340–345. Дои:10.1016 / s0958-1669 (97) 80014-6. PMID 9206017.
- ^ Wright, E .; Баум, Д. (2018). «Эксклюзивность предлагает надежный, но практичный критерий вида бактерий, несмотря на обильный поток генов». BMC Genomics. 19 (724): 724. Дои:10.1186 / s12864-018-5099-6. ЧВК 6171291. PMID 30285620.
- ^ Lan, R; Ривз, PR (2002). «Замаскированная Escherichia coli: молекулярное происхождение шигелл». Микробы и инфекции / Institut Pasteur. 4 (11): 1125–32. Дои:10.1016 / S1286-4579 (02) 01637-4. PMID 12361912.
- ^ Бавыкин, С.Г .; Лысов, Ю.П .; Захариев, В .; Kelly, J. J .; Jackman, J .; Stahl, D. A .; Черни, А. (2004). «Использование 16S рРНК, 23S рРНК и анализа последовательности гена gyrB для определения филогенетических отношений микроорганизмов группы Bacillus cereus». Журнал клинической микробиологии. 42 (8): 3711–30. Дои:10.1128 / JCM.42.8.3711-3730.2004. ЧВК 497648. PMID 15297521.
- ^ Ахтман, М .; Zurth, K .; Морелли, G .; Torrea, G .; Guiyoule, A .; Карниэль, Э. (1999). «Yersinia pestis, вызывающая чуму, является недавно появившимся клоном Yersinia pseudotuberculosis». Труды Национальной академии наук Соединенных Штатов Америки. 96 (24): 14043–14048. Bibcode:1999PNAS ... 9614043A. Дои:10.1073 / пнас.96.24.14043. ЧВК 24187. PMID 10570195.
- ^ Young, J.M .; Парк, Д. -К. (2007). «Вероятная синонимия азотфиксирующего рода Azotobacter и рода Pseudomonas». Международный журнал систематической и эволюционной микробиологии. 57 (12): 2894–2901. Дои:10.1099 / ijs.0.64969-0. PMID 18048745.
- ^ Rediers, H; Vanderleyden, J; Де Мот, Р. (2004). «Azotobacter vinelandii: замаскированная псевдомонада?». Микробиология. 150 (Пт 5): 1117–9. Дои:10.1099 / мик. 0.27096-0. PMID 15133068.
- ^ Сюй, Д; Côté, JC (2003). «Филогенетические отношения между видами Bacillus и родственными родами, выведенные из сравнения нуклеотидных последовательностей 3'-конца 16S рДНК и 5'-конца 16S-23S ITS». Международный журнал систематической и эволюционной микробиологии. 53 (Pt 3): 695–704. Дои:10.1099 / ijs.0.02346-0. PMID 12807189.
- ^ Young, J .; Kuykendall, L .; Martínez-Romero, E .; Kerr, A .; Савада, Х. (2001). "Пересмотр Rhizobium Frank 1889 с исправленным описанием рода и включением всех видов Agrobacterium Conn 1942 и Allorhizobium undicola de Lajudie et al. 1998 в качестве новых комбинаций: Rhizobium radiobacter, R. Rhizogenes, R. Rubi, Р. Ундикола и Р. Витис ". Международный журнал систематической и эволюционной микробиологии. 51 (Чт 1): 89–103. Дои:10.1099/00207713-51-1-89. PMID 11211278.
- ^ Farrand, S .; Van Berkum, P .; Огер, П. (2003). «Agrobacterium - определяемый род семейства Rhizobiaceae». Международный журнал систематической и эволюционной микробиологии. 53 (Pt 5): 1681–1687. Дои:10.1099 / ijs.0.02445-0. PMID 13130068.
- ^ Young, J .; Kuykendall, L .; Martínez-Romero, E .; Kerr, A .; Савада, Х. (2003). «Классификация и номенклатура Agrobacterium и Rhizobium». Международный журнал систематической и эволюционной микробиологии. 53 (Pt 5): 1689–1695. Дои:10.1099 / ijs.0.02762-0. PMID 13130069.
- ^ К. Джеффри. 1989. Биологическая номенклатура, 3-е изд. Эдвард Арнольд, Лондон, 86 стр.
- ^ приложение. 9 г. до н.э. https://www.ncbi.nlm.nih.gov/books/NBK8808/
- ^ а б c d Помощь! Латинский! Как избежать самых распространенных ошибок при присвоении латинских имен вновь обнаруженным прокариотам. Microbiología (Sociedad Española de Microbiología), 1996, 12, 473-475. «Архивная копия». Архивировано из оригинал 7 октября 2011 г.. Получено 14 апреля 2011.CS1 maint: заархивированная копия как заголовок (связь)
- ^ Классификация статьи Phyla в LPSN; Euzéby, J.P. (1997). «Список названий бактерий с позиции в номенклатуре: папка, доступная в Интернете». Международный журнал систематической и эволюционной микробиологии. 47 (2): 590–2. Дои:10.1099/00207713-47-2-590. PMID 9103655.
- ^ STACKEBRANDT, E .; Дождливый; УОРД-РЕЙНИ, Н.Л. (1997). «Предложение по новой иерархической системе классификации Actinobacteria classis nov». Int. J. Syst. Бактериол. 47 (2): 479–491. Дои:10.1099/00207713-47-2-479.
- ^ а б Классификационная запись в LPSN; Euzéby, J.P. (1997). «Список названий бактерий с позиции в номенклатуре: папка, доступная в Интернете». Международный журнал систематической и эволюционной микробиологии. 47 (2): 590–2. Дои:10.1099/00207713-47-2-590. PMID 9103655.
- ^ а б Иерархическая классификация записи прокариот в LPSN; Euzéby, J.P. (1997). «Список названий бактерий с позиции в номенклатуре: папка, доступная в Интернете». Международный журнал систематической и эволюционной микробиологии. 47 (2): 590–2. Дои:10.1099/00207713-47-2-590. PMID 9103655.
- ^ Мюррей РГЭ. (1984). «Высшие таксоны или место всему…?». В Krieg NR, Holt JG (ed.). Руководство по систематической бактериологии Берджи. 1. Балтимор: Williams & Wilkins Co., стр. 33.
- ^ Орен А., Да Коста М.С., Гаррити ГМ, Рейни Ф.А., Росселло-Мора Р., Шинк Б., Сатклифф I, Трухильо М.Э., Уитман В.Б. (2015). «Предложение включить ранг типа в Международный кодекс номенклатуры прокариот». Int. J. Syst. Evol. Микробиол. 65 (11): 4284–4287. Дои:10.1099 / ijsem.0.000664. PMID 26654112.
- ^ ШРОЭТЕР (Дж.). В: F. COHN (ред.), Kryptogamenflora von Schlesien. Band 3, Heft 3, Pilze. J.U. Kern's Verlag, Бреслау, 1885–1889, стр. 1–814.
- ^ Поступление сальмонеллы в LPSN; Euzéby, J.P. (1997). «Список названий бактерий с позиции в номенклатуре: папка, доступная в Интернете». Международный журнал систематической и эволюционной микробиологии. 47 (2): 590–2. Дои:10.1099/00207713-47-2-590. PMID 9103655.
- ^ Р. Э. БУКАНАН, Таксономия, Annu. Rev. Microbiol. 1955.9: 1-20. http://www.annualreviews.org/doi/pdf/10.1146/annurev.mi.09.100155.000245
- ^ ftp://ftp.ncbi.nih.gov/pub/taxonomy/