Физиология рыб - Fish physiology

Физиология рыб это научное исследование того, как составные части рыбы действуют вместе в живой рыбе.[2] Его можно противопоставить анатомия рыбы, что является изучением формы или морфология рыб. На практике анатомия и физиология рыбы дополняют друг друга, первая касается строения рыбы, ее органов или составных частей и того, как они устроены вместе, что можно наблюдать на препаровальном столе или под микроскопом, а более поздние. выясняет, как эти компоненты работают вместе у живых рыб.
Дыхание
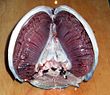


Большинство рыб обмениваются газами с использованием жабры по обе стороны от глотка (горло). Жабры - это ткани, которые состоят из нитевидных структур, называемых нити. Эти нити выполняют множество функций и «участвуют в переносе ионов и воды, а также в обмене кислорода, диоксида углерода, кислоты и аммиака.[3][4] Каждая нить содержит капилляр сеть, обеспечивающая большой площадь поверхности для обмена кислород и углекислый газ. Рыбы обмениваются газами, вытягивая богатую кислородом воду через рот и перекачивая ее через жабры. У некоторых рыб капиллярная кровь течет в направлении, противоположном воде, вызывая встречный обмен. Жабры выталкивают бедную кислородом воду через отверстия по бокам глотки.
Рыбы из нескольких групп могут жить вне воды в течение продолжительных периодов времени. Рыба-амфибия такой как грязевой прыгун могут жить и передвигаться по суше до нескольких дней или жить в стоячей или иным образом обедненной кислородом воде. Многие из таких рыб могут дышать воздухом с помощью различных механизмов. Кожа ангиллиды может напрямую поглощать кислород. В Полость рта из электрический угорь может дышать воздухом. Сом из семей Loricariidae, Callichthyidae, и Scoloplacidae впитывают воздух через пищеварительный тракт.[5] Двоякодышащий, за исключением австралийской двоякодышащей рыбы, и бихиры имеют парные легкие, похожие на легкие четвероногие и должен всплывать, чтобы глотать свежий воздух через рот и пропускать отработанный воздух через жабры. Гар и Bowfin имеют васкуляризованный плавательный пузырь, который функционирует таким же образом. Вьюны, Trahiras, и много сом дышите, пропуская воздух через кишечник. Грязевые прыгуны дышат, поглощая кислород кожей (подобно лягушкам). У ряда рыб появились так называемые вспомогательные органы дыхания которые извлекают кислород из воздуха. Лабиринтная рыба (например, гурами и петушки ) иметь лабиринтный орган над жабрами, выполняющими эту функцию. У некоторых других рыб есть структуры, напоминающие органы лабиринта по форме и функциям, в первую очередь змееголовы, щуки, а Clariidae семейство сомовых.
Воздух для дыхания в первую очередь используется для рыб, обитающих в мелководных сезонно изменчивых водах, где концентрация кислорода в воде может сезонно снижаться. Рыбы зависят исключительно от растворенного кислорода, например, окунь и цихлиды, быстро задыхаются, в то время как дышащие воздухом выживают намного дольше, в некоторых случаях в воде, которая представляет собой немного больше, чем влажная грязь. В самом крайнем случае, некоторые дышащие воздухом рыбы способны неделями выжить во влажных норах без воды, переходя в состояние праздник (летняя спячка) до возвращения воды.
Рыб, дышащих воздухом, можно разделить на обязать сапуны и факультативный воздушные сапуны. Обязательные сапуны, такие как Африканская двоякодышащая рыба, вынуждены периодически дышать воздухом или задохнуться. Факультативные дыхательные аппараты, такие как сом. Hypostomus plecostomus, дышат воздухом только в случае необходимости, иначе они могут полагаться на свои жабры для получения кислорода. Большинство дышащих воздухом рыб являются факультативными дышащими воздухом, что позволяет избежать энергетических затрат, связанных с подъемом на поверхность, и затрат на пригодность для контакта с поверхностными хищниками.[5]

системы обмена потоками
Все базальный позвоночные животные дышат жабры. Жабры расположены прямо за головой, ограничивая задние края ряда отверстий от пищевод к экстерьеру. Каждая жабра поддерживается хрящевой или костной жаберная дуга.[9] Жабры позвоночные обычно развиваются в стенах глотка вдоль ряда выходящих наружу жаберных щелей. Большинство видов используют встречный обмен система для усиления диффузии веществ в жабры и из них, при этом кровь и вода текут в противоположных направлениях друг к другу, что увеличивает эффективность поглощения кислорода из воды.[6][7][8] Свежая насыщенная кислородом вода, поступающая через рот, непрерывно «прокачивается» через жабры в одном направлении, в то время как кровь в пластинках течет в противоположном направлении, создавая противоток крови и воды, от которого зависит выживание рыбы.[8]
Жабры состоят из гребнеобразных волокон, жаберные ламели, которые помогают увеличить площадь их поверхности для кислородного обмена.[10] Когда рыба дышит, она через равные промежутки времени набирает в рот воду. Затем он сближает стороны своего горла, выталкивая воду через жаберные отверстия так, чтобы она проходила через жабры наружу. В костлявая рыба иметь три пары арок, хрящевые рыбы имеют пять-семь пар, а примитивные рыба без челюсти есть семь. У предка позвоночных, несомненно, было больше дуг, поскольку некоторые из их хордовый у родственников более 50 пар жабр.[11]
Высшие позвоночные не развиваются жабры, во время развитие плода, и заложить основу основных структур, таких как челюсти, то щитовидная железа, то гортань, то Колумелла (соответствует стремени в млекопитающие ) и у млекопитающих молоток и наковальня.[11] Жаберные щели у рыб могут быть эволюционными предками миндалины, вилочковая железа, и Евстахиевы трубы, а также многие другие структуры, происходящие от эмбрионального жаберные мешочки.[нужна цитата ]
Ученые исследовали, какая часть тела отвечает за поддержание дыхательного ритма. Они обнаружили, что нейроны расположен в мозговой ствол рыб ответственны за генезис дыхательного ритма.[12] Положение этих нейронов немного отличается от центров респираторного генеза у млекопитающих, но они расположены в одном отделе мозга, что вызвало споры по поводу гомология дыхательных центров между водными и наземными видами. Как в водном, так и в наземном дыхании точные механизмы, с помощью которых нейроны могут генерировать этот непроизвольный ритм, до сих пор полностью не изучены (см. Непроизвольный контроль дыхания ).
Другой важной особенностью дыхательного ритма является то, что он регулируется для адаптации к потреблению кислорода организмом. Как наблюдали у млекопитающих, рыбы «дышат» быстрее и тяжелее, когда они дышат. физическое упражнение. Механизмы, с помощью которых происходят эти изменения, обсуждались учеными более 100 лет.[13] Авторов можно разделить на 2 школы:
- Те, кто думают, что основная часть респираторных изменений заранее запрограммированы в мозге, что означает, что нейроны из движение центры мозга соединяются с дыхательные центры в ожидании движений.
- Те, кто думает, что основная часть респираторных изменений является результатом обнаружения сокращения мышц, а дыхание адаптируется как следствие сокращения мышц и потребления кислорода. Это означало бы, что мозг обладает какими-то механизмами обнаружения, которые запускают респираторную реакцию при сокращении мышц.
Многие теперь согласны с тем, что оба механизма, вероятно, присутствуют и дополняют друг друга или работают вместе с механизмом, который может обнаруживать изменения в насыщении крови кислородом и / или углекислым газом.
Костяная рыба

В костлявая рыба, жабры лежат в жаберной камере, покрытой костной крышечкой. Подавляющее большинство видов костистых рыб имеют пять пар жабр, хотя некоторые из них потеряли некоторые в ходе эволюции. Жаберная крышка может иметь важное значение для регулирования давления воды внутри глотки, чтобы обеспечить надлежащую вентиляцию жабр, так что костлявым рыбам не нужно полагаться на вентиляцию барана (и, следовательно, почти постоянное движение), чтобы дышать. Клапаны во рту предотвращают утечку воды.[11]
Жаберные дуги костистых рыб обычно не имеют перегородки, так что одни жабры выступают из дуги, поддерживаясь отдельными жаберными лучами. У некоторых видов сохраняются жаберные тычинки. Хотя у всех, кроме самых примитивных костистых рыб, дыхальца отсутствует, псевдожабро, связанное с ним, часто остается, находясь у основания жаберной крышки. Однако она часто значительно уменьшается, так как состоит из небольшой массы клеток без какой-либо оставшейся жаброобразной структуры.[11]

морской костистые кости также используйте жабры для выделения электролиты. Большая площадь поверхности жабр имеет тенденцию создавать проблемы для рыб, которые стремятся регулировать осмолярность их внутренних жидкостей. Морская вода менее разбавлена, чем эти внутренние жидкости, поэтому морские рыбы теряют осмотически большое количество воды через жабры. Чтобы восстановить воду, они пьют большое количество морская вода и выделять соль. Однако пресная вода более разбавлена, чем внутренние жидкости рыб, поэтому пресноводные рыбы получают воду. осмотически через жабры.[11]
У некоторых примитивных костистых рыб и амфибии, то личинки несут наружные жабры, отходящие от жаберных дуг.[14] В зрелом возрасте они сокращаются, их функцию берут на себя собственно жабры у рыб и легкие у большинства амфибий. Некоторые амфибии сохраняют внешние личиночные жабры во взрослом возрасте, сложная внутренняя жаберная система, наблюдаемая у рыб, по-видимому, безвозвратно утрачивается очень рано в эволюции четвероногие.[15]
Хрящевые рыбы
Как и другие рыбы, акулы добывают кислород от морской воды, когда она проходит через их жабры. В отличие от других рыб, жаберные щели акулы не закрыты, а лежат рядом за головой. Модифицированная щель, названная дыхальце лежит сразу за глаз, который помогает акуле взять воду во время дыхание и играет главную роль в обитающих на дне акулах. Дыхальца уменьшены или отсутствуют в активном пелагический акулы.[16] Пока акула движется, вода проходит через рот и жабры в процессе, известном как «вентиляция барана». В состоянии покоя большинство акул качает воду через жабры, чтобы обеспечить постоянный приток насыщенной кислородом воды. Небольшое количество видов потеряли способность качать воду через жабры и вынуждены плавать без отдыха. Эти виды облигатные вытяжные вентиляторы и предположительно задыхаться если не может двигаться. Облигатная вентиляция барана также характерна для некоторых пелагических костных видов рыб.[17]
Дыхание и обращение процесс начинается при дезоксигенировании кровь путешествует по акульей двухкамерной сердце. Здесь акула качает кровь к своим жабрам через брюшную полость. аорта артерия где это разветвляется в афферентный плечевой артерии. Реоксигенация происходит в жабрах, и реоксигенированная кровь течет в эфферент плечевые артерии, которые вместе образуют дорсальная аорта. Кровь течет из спинной аорты по всему телу. Дезоксигенированная кровь из организма затем течет через задние кардинальные вены и входит в задний кардинал пазухи. Оттуда кровь поступает в сердце желудочек и цикл повторяется.[18]
Акулы и лучи обычно имеют пять пар жаберных щелей, которые открываются непосредственно наружу тела, хотя у некоторых более примитивных акул их шесть или семь пар. Соседние щели разделены хрящевой жаберная дуга, из которой выступает длинный листовой перегородка, частично поддерживаемый другим куском хряща, называемым жаберный луч. Отдельные пластинки жабр лежат по обе стороны от перегородки. Основание арки также может поддерживать жаберные тычинки, небольшие выступающие элементы, которые помогают фильтровать пищу из воды.[11]
Меньшее отверстие, дыхальце, лежит в задней части первого жаберная щель. Это несет небольшой псевдожабица который по структуре напоминает жабры, но получает кровь, уже насыщенную кислородом настоящих жабр.[11] Считается, что дыхальце гомологичный к ушному отверстию у высших позвоночных.[19]
Большинство акул полагаются на вентиляцию барана, заставляя воду течь в рот и через жабры, быстро плывя вперед. У медленно передвигающихся или обитающих на дне видов, особенно среди скатов и скатов, дыхальце может быть увеличено, и рыба дышит, всасывая воду через это отверстие, а не через рот.[11]
Химеры отличаются от других хрящевых рыб тем, что лишились дыхальца и пятой жаберной щели. Остальные щели закрыты крышка, развился из перегородки жаберной дуги перед первой жаброй.[11]
Миноги и миксины
Миноги и миксина как таковых жаберных щелей нет. Вместо этого жабры заключены в сферические мешочки с круглым отверстием наружу. Как и в жаберных щелях более высокой рыбы, в каждом мешочке есть по две жабры. В некоторых случаях отверстия могут срастаться, образуя крышечку. У миног семь пар мешочков, у миксин - от шести до четырнадцати, в зависимости от вида. У миксины мешочки соединяются с глоткой изнутри. У взрослых миног отдельная дыхательная трубка развивается под собственно глоткой, отделяя пищу и воду от дыхания, закрывая клапан на его переднем конце.[11]
Тираж

Системы кровообращения всех позвоночные находятся закрыто, как и у людей. Тем не менее, системы рыбы, амфибии, рептилии, и птицы показать различные этапы эволюция из сердечно-сосудистая система. У рыб система имеет только один контур, в котором кровь перекачивается через капилляры жабры и капилляры тканей тела. Это известно как одиночный цикл тираж. Таким образом, сердце рыбы - это только один насос (состоящий из двух камер). У рыбы есть замкнутая система кровообращения. В сердце перекачивает кровь по единой петле по всему телу. У большинства рыб сердце состоит из четырех частей, включая две камеры, вход и выход.[20] Первая часть - это венозный синус, тонкостенный мешок, собирающий кровь из рыбьей вены прежде чем позволить ему течь во вторую часть, Атриум, который представляет собой большую мышечную камеру. Атриум служит односторонним вестибюлем, отправляет кровь в третью часть, желудочек. Желудочек - это еще одна толстостенная мышечная камера, которая перекачивает кровь из первой части в четвертую, артериальная луковица, большая трубка, а потом из сердца. Артериальная луковица соединяется с аорта, по которому кровь течет к жабры для оксигенации.
У амфибий и большинства рептилий двойная кровеносная система используется, но сердце не всегда полностью разделяется на две помпы. Амфибии имеют трехкамерный сердце.
Пищеварение
Челюсти позволяют рыбе есть разнообразную пищу, включая растения и другие организмы. Рыба глотает пищу через рот и расщепляет ее в пищевод. В желудке пища дополнительно переваривается и у многих рыб перерабатывается в мешочках в форме пальцев, называемых пилорическая слепая кишка, которые выделяют пищеварительные ферменты и впитывают питательные вещества. Такие органы, как печень и поджелудочная железа добавляйте ферменты и различные химические вещества по мере прохождения пищи по пищеварительному тракту. В кишечнике завершается процесс пищеварения и всасывания питательных веществ.
У большинства позвоночных пищеварение представляет собой четырехэтапный процесс, в котором задействованы основные структуры позвоночных. пищеварительный тракт, начиная с приема пищи, помещения пищи в рот и заканчивая выведением непереваренного материала через задний проход. Изо рта пища попадает в желудок, где по мере болюс он расщепляется химически. Затем он перемещается в кишечник, где продолжается процесс расщепления пищи на простые молекулы, и результаты абсорбируются в виде питательных веществ в кровеносную и лимфатическую систему.
Хотя точная форма и размер желудка сильно различаются у разных позвоночных, относительное положение отверстий пищевода и двенадцатиперстной кишки остается относительно постоянным. В результате орган всегда несколько изгибается влево, прежде чем повернуться назад, чтобы встретиться с пилорическим сфинктером. Тем не мение, миноги, микробы, химеры, двоякодышащие, и немного костистость у рыб вообще нет желудка, а пищевод открывается прямо в кишечник. Все эти животные потребляют диеты, которые либо требуют небольшого хранения пищи, либо не переваривают желудочный сок, либо и то, и другое.[11]
В тонкий кишечник часть пищеварительного тракта, следующая за желудком, за которым следует толстая кишка, и здесь большая часть пищеварение и происходит всасывание пищи. У рыб неясны отделы тонкой кишки, а сроки передний или же проксимальный кишечник может использоваться вместо двенадцатиперстная кишка.[21] Тонкая кишка встречается во всех костистые кости, хотя его форма и длина сильно различаются между видами. У костистых рыб он относительно короткий, обычно в полтора раза длиннее тела рыбы. Обычно он имеет ряд пилорическая слепая кишка, небольшие мешковидные структуры по длине, которые помогают увеличить общую площадь поверхности органа для переваривания пищи. У костистых кистей нет илеоцекального клапана, а граница между тонкой кишкой и кишечником отсутствует. прямая кишка отмечен только концом пищеварительного эпителия.[11]
У не костистых рыб, таких как акулы, осетровые, и двоякодышащая рыба. Вместо этого пищеварительная часть кишечника образует спиральный кишечник, соединяющий желудок с прямой кишкой. В этом типе кишечника кишечник относительно прямой, но имеет длинную складку, проходящую по внутренней поверхности по спирали, иногда с десятками витков. Этот клапан значительно увеличивает как площадь поверхности, так и эффективную длину кишечника. Выстилка спирального кишечника похожа на выстилку тонкого кишечника костистых и четвероногих не млекопитающих.[11] В миноги, спиральный клапан чрезвычайно мал, возможно, потому, что их диета требует небольшого переваривания. Hagfish не имеют вообще никакого спирального клапана, пищеварение происходит почти по всей длине кишечника, который не разделен на разные области.[11]
В толстая кишка это последняя часть пищеварительная система обычно встречается у позвоночных животных. Его функция состоит в том, чтобы поглощать воду из оставшейся неперевариваемой пищи, а затем выводить бесполезные отходы от тела.[22] У рыб нет настоящей толстой кишки, а есть просто короткая прямая кишка, соединяющая конец пищеварительной части кишечника с клоакой. В акулы, это включает ректальная железа который выделяет соль, чтобы помочь животному поддерживать осмотический балансировать с морской водой. Железа чем-то напоминает слепую кишку по строению, но не является гомологичной структурой.[11]
Как и многие другие водные животные, большинство рыб выделяют азотистые отходы в виде аммиак. Некоторые отходы размытый через жабры. Отходы крови фильтрованный посредством почки.
Морские рыбы склонны терять воду из-за осмос. Их почки возвращают воду в организм. Обратное происходит в пресноводная рыба: они имеют тенденцию осмотически набирать воду. Их почки производят разбавленную мочу для выделения. У некоторых рыб есть специально адаптированные почки, которые различаются по функциям, что позволяет им переходить из пресной в соленую.
У акул пищеварение может занять много времени. Пища перемещается изо рта в желудок J-образной формы, где она хранится и происходит начальное пищеварение.[23] Нежелательные предметы могут никогда не пройти мимо желудка, и вместо этого акула либо рвет, либо выворачивает живот наизнанку и выбрасывает нежелательные предметы изо рта. Одно из самых больших различий между пищеварительной системой акул и млекопитающих заключается в том, что у акул гораздо более короткий кишечник. Эта короткая длина достигается за счет спиральный клапан с множеством витков в одном коротком отрезке вместо длинного трубчатого кишечника. Клапан имеет большую площадь поверхности, поэтому пища должна циркулировать внутри короткого кишечника до полного переваривания, когда оставшиеся продукты жизнедеятельности попадают в кишечник. клоака.[23]
Эндокринная система
Регулирование социального поведения
Окситоцин это группа нейропептиды встречается у большинства позвоночных. Одна из форм окситоцина действует как гормон что связано с человеческой любовью. В 2012 году исследователи ввели цихлиды от социальных видов Neolamprologus pulcher либо с этой формой изотоцина, либо с контрольным физиологическим раствором. Они обнаружили, что изотоцин увеличивает «реакцию на социальную информацию», что предполагает «это ключевой регулятор социального поведения, которое развивалось и сохранялось с древних времен».[24][25]
Последствия загрязнения
Рыба может биоаккумулировать загрязняющие вещества, сбрасываемые в водные пути. Эстрогенный соединения, содержащиеся в пестицидах, противозачаточных средствах, пластмассах, растениях, грибах, бактериях и синтетических наркотиках, попадающих в реки, влияют на эндокринные системы местных видов.[26] В Боулдере, Колорадо, белый присоска рыбы, обнаруженные ниже по течению от городских очистных сооружений, демонстрируют нарушенное или ненормальное половое развитие. Рыба подверглась воздействию более высоких уровней эстрогена, что привело к появлению феминизированных рыб.[27] Самцы демонстрируют женские репродуктивные органы, и у обоих полов снижена фертильность и выше смертность при вылуплении.[28]
Пресноводные среды обитания в Соединенных Штатах широко загрязнены обычными пестицидами. атразин.[29] Существуют разногласия по поводу степени, в которой этот пестицид вредит эндокринной системе пресноводных рыб и амфибии. Исследователи, не финансируемые отраслью, постоянно сообщают о вредных последствиях, в то время как исследователи, финансируемые отраслью, постоянно не сообщают о вредных последствиях.[29][30][31]
В морской экосистеме хлорорганические соединения загрязнители, такие как пестициды, гербициды (ДДТ) и хлордан накапливаются в тканях рыб и нарушают их эндокринную систему.[32] Высокая частота бесплодия и высокий уровень хлорорганических соединений были обнаружены в капотные акулы вдоль побережья Мексиканского залива Флориды. Эти эндокринные нарушения Составы сходны по структуре с естественными гормонами рыб. Они могут модулировать гормональные взаимодействия у рыб:[33]
- связывание с клеточными рецепторами, вызывая непредсказуемую и ненормальную активность клеток
- блокирование рецепторных сайтов, ингибирование активности
- содействие созданию дополнительных рецепторных участков, усиление эффектов гормона или соединения
- взаимодействуя с естественными гормонами, изменяя их форму и влияние
- влияет на синтез или метаболизм гормонов, вызывая неправильный баланс или количество гормонов
Осморегуляция


Два основных типа осморегуляция осмоконформаторы и осморегуляторы. Осмоконформеры активно или пассивно сопоставляют осмолярность своего тела с окружающей средой. Большинство морских беспозвоночных являются осмоконформерами, хотя их ионный состав может отличаться от состава морской воды.
Осморегуляторы жестко регулировать свое тело осмолярность, который всегда остается постоянным и чаще встречается в животном мире. Осморегуляторы активно контролируют концентрацию соли, несмотря на концентрацию соли в окружающей среде. Пример - пресноводная рыба. Жабры активно усваивать соль из окружающей среды за счет использования богатых митохондриями клеток. Вода будет проникать в рыбу, поэтому она выделяет очень гипотонический (разбавить) мочу, чтобы удалить всю лишнюю воду. Морской пехотинец рыбы имеет внутреннюю осмотическую концентрацию ниже, чем у окружающей морской воды, поэтому он имеет тенденцию терять воду и накапливать соль. Активно выводит из организма соль из жабры. Большинство рыб стеногалин, что означает, что они ограничены либо соленой, либо пресной водой и не могут выжить в воде с концентрацией соли, отличной от той, к которой они приспособлены. Однако некоторые рыбы демонстрируют огромную способность эффективно осморегулировать в широком диапазоне солености; рыбы с этой способностью известны как эвригалин виды, например, лосось. Было замечено, что лосось обитает в двух совершенно разных средах - морской и пресной - и ему свойственно адаптироваться к обоим, внося поведенческие и физиологические изменения.
В отличие от костистых рыб, за исключением латимерия,[34] кровь и другие ткани акул и Chondrichthyes обычно изотонический в их морскую среду из-за высокой концентрации мочевина и триметиламин N-оксид (TMAO), позволяющий им находиться в осмотический балансировать с морской водой. Эта адаптация препятствует выживанию большинства акул в пресной воде, и поэтому они ограничиваются морской среды. Существует несколько исключений, например бычья акула, который разработал способ изменить свой почка функция по выведению большого количества мочевины.[35] Когда акула умирает, мочевина расщепляется бактериями до аммиака, в результате чего мертвое тело постепенно начинает сильно пахнуть аммиаком.[36][37]
Акулы применили другой, эффективный механизм сохранения воды, а именно осморегуляцию. Они сохраняют в крови мочевину в относительно более высокой концентрации. Мочевина повреждает живые ткани, поэтому, чтобы справиться с этой проблемой, некоторые рыбы сохраняют оксид триметиламина. Это лучшее решение проблемы токсичности мочевины. Акулы, имеющие немного более высокую концентрацию растворенных веществ (то есть более 1000 мОсм, что является концентрацией растворенных веществ в море), не пьют воду, как пресноводные рыбы.
Терморегуляция
Гомеотермия и пойкилотермия относятся к тому, насколько стабильна температура организма. Наиболее эндотермический организмы гомеотермны, как млекопитающие. Однако животные с факультативным эндотермия часто пойкилотермные, то есть их температура может значительно варьироваться. Точно так же большинство рыб эктотермия, так как все их тепло исходит от окружающей воды. Однако большинство из них являются домашними животными, потому что их температура очень стабильна.
Большинство организмов имеют предпочтительный температурный диапазон, однако некоторые могут адаптироваться к температурам более холодным или более теплым, чем те, к которым они обычно привыкли. Предпочтительной температурой организма обычно является температура, при которой физиологические процессы организма могут действовать с оптимальной скоростью. Когда рыба привыкает к другим температурам, эффективность их физиологических процессов может снизиться, но она продолжит функционировать. Это называется тепловая нейтральная зона при которых организм может выжить бесконечно.[38]
H.M. Вернон провел работу по оценке температуры смерти и температуры паралича (температуры теплового окоченения) различных животных. Он обнаружил, что виды одного и того же учебный класс показали очень похожие значения температуры, Амфибия обследовано - 38,5 ° C, рыб - 39 ° C, Рептилии 45 ° C, и различные Моллюски 46 ° С.
Чтобы справиться с низкими температурами, некоторые рыбы развили способность оставаться работоспособным даже при температуре воды ниже нуля; некоторые используют натуральные антифриз или же антифризы протеины чтобы противостоять образованию кристаллов льда в тканях.
Большинство акул «хладнокровны» или, точнее, пойкилотермный, что означает, что их внутренние температура тела соответствует их окружающей среде. Члены семьи Lamnidae (такой как короткопёрая акула мако и Большая белая акула ) находятся теплый и поддерживать более высокую температуру тела, чем окружающая вода. У этих акул полоска аэробный красная мышца, расположенная недалеко от центра тела, генерирует тепло, которое тело сохраняет через встречный обмен механизм системой кровеносный сосуд называется Rete Mirabile («чудесная сеть»). В обыкновенная акула-молотильщик имеет аналогичный механизм поддержания повышенной температуры тела, который, как считается, развился независимо[неудачная проверка ].[39]
Тунец могут поддерживать температуру определенных частей своего тела выше температуры окружающей морской воды. Например, тунец поддерживайте внутреннюю температуру тела 25–33 ° C (77–91 ° F) в воде до 6 ° C (43 ° F). Однако, в отличие от типичных эндотермических существ, таких как млекопитающие и птицы, тунец не поддерживает температуру в относительно узких пределах.[40][41] Тунец достиг эндотермия за счет сохранения тепла, выделяемого обычным метаболизм. В Rete Mirabile («чудесная сеть»), переплетение вен и артерий на периферии тела, передает тепло от венозная кровь к артериальная кровь через противоточный обмен система, таким образом смягчая эффекты поверхностного охлаждения. Это позволяет тунцу повышать температуру очень аэробный ткани скелетных мышц, глаз и мозга,[40][42] который поддерживает более высокие скорости плавания и снижает расход энергии, а также позволяет им выживать в более прохладных водах в более широком диапазоне океанических сред, чем у других рыб.[41] Однако у всех тунцов сердце работает на температура окружающей среды, так как в него поступает охлажденная кровь, а коронарное кровообращение идет прямо из жабры.[42]
- Гомеотермия: Хотя большинство рыб исключительно экзотермический, бывают исключения. Некоторые виды рыб поддерживают повышенную температуру тела. Эндотермический костистые кости (костлявые рыбы) все в подотряде Scombroidi и включить рыбки, тунец, в том числе "примитивный" скумбрия разновидность, Гастерохизма мелампус. Все акулы в семье Lamnidae - мако с коротким плавником, мако с длинным плавником, белая акула, поросенок и лососевая акула - являются эндотермическими, и данные свидетельствуют о том, что эта черта существует в семье Alopiidae (акулы-молотилки ). Степень эндотермии варьирует от морская рыба, которые согревают только глаза и мозг, чтобы тунец и белоснежные акулы которые поддерживают температуру тела на 20 ° C (36 ° F) выше температуры окружающей воды.[43] Смотрите также гигантотермия. Считается, что эндотермия, хотя и требует больших затрат на метаболизм, дает такие преимущества, как увеличение мышечной силы, более высокие показатели центрального нервная система обработка, и более высокие темпы пищеварение.
У некоторых рыб Rete Mirabile допускает повышение температуры мышц в регионах, где находится эта сеть вен и артерий. Рыба умеет терморегулятор определенные области их тела. Кроме того, это повышение температуры приводит к повышению температуры основного обмена. Теперь рыба может делиться АТФ с большей скоростью и, в конечном итоге, может плавать быстрее.
Глаз рыба-меч может генерировать высокая температура чтобы лучше справляться с обнаружением их добыча на глубине 600 метров (2000 футов).[44]
Мышечная система


Рыба плавает за счет сокращения продольной красной мышцы и косо ориентированной белой мышцы. Красная мышца аэробна и нуждается в кислороде, который поступает от миоглобин. Белая мышца анаэробна и не нуждается в кислороде. Красные мышцы используются для постоянной активности, например, для плавания на малых скоростях во время миграции в океан. Белые мышцы используются для всплесков активности, таких как прыжки или резкие всплески скорости для ловли добычи.[45]
В основном у рыб белые мускулы, но у некоторых рыб, таких как скомброиды и лососевые от розового до темно-красного. Красный миотомальный мышцы получают свой цвет от миоглобин, молекула, связывающая кислород, количество которой у тунца намного выше, чем у большинства других рыб. Богатая кислородом кровь также обеспечивает доставку энергии к мышцам.[40]
Большинство рыб движутся за счет попеременного сокращения парных наборов мышц по обе стороны от позвоночника. Эти сокращения образуют S-образные кривые, которые движутся вниз по телу. Когда каждая кривая достигает задний плавник к воде прилагается обратная сила, которая вместе с плавниками перемещает рыбу вперед. В плавники рыбы работают как закрылки самолета. Плавники также увеличивают площадь поверхности хвоста, увеличивая скорость. Обтекаемое тело рыбы снижает трение о воду.
Типичной характеристикой многих животных, использующих волнообразные движения, является то, что они имеют сегментированные мышцы или блоки мышц. миомеры, идущие от головы к хвостам, которые разделены соединительной тканью, называемой миосептами. Кроме того, некоторые сегментированные группы мышц, такие как боковая гипаксиальная мускулатура у саламандры, ориентированы под углом к продольному направлению. Для этих наклонно ориентированных волокон напряжение в продольном направлении больше, чем напряжение в направлении мышечных волокон, что приводит к архитектурному передаточному отношению более 1. Более высокий начальный угол ориентации и большее дорсовентральное выпячивание вызывают более быстрое сокращение мышц, но приводят к меньшее количество производимой силы.[46] Предполагается, что животные используют механизм переменной передачи, который позволяет саморегулировать силу и скорость для удовлетворения механических требований сокращения.[47] Когда перистая мышца подвергается воздействию небольшой силы, сопротивление изменениям ширины мышцы заставляет ее вращаться, что, следовательно, обеспечивает более высокое архитектурное передаточное число (AGR) (высокая скорость).[47] Однако при воздействии большой силы перпендикулярная составляющая силы волокна преодолевает сопротивление изменениям ширины, и мышцы сжимаются, производя более низкий AGR (способный поддерживать более высокую выходную силу).[47]
Большинство рыб изгибаются как простой однородный пучок во время плавания за счет сокращений продольных красных мышечных волокон и наклонно ориентированных белых мышечных волокон в сегментированной осевой мускулатуре. Волокно напряжение (εf), испытываемая продольными красными мышечными волокнами, эквивалентна продольной деформации (εx). Более глубокие белые мышечные волокна рыб отличаются разнообразием расположения. Эти волокна организованы в конусообразные структуры и прикрепляются к листам соединительной ткани, известным как миосепта; каждое волокно показывает характерную дорсовентральную (α) и медиолатеральную (φ) траектории. Теория сегментированной архитектуры предсказывает, что εx> εf. Это явление приводит к архитектурному передаточному отношению, определяемому как продольная деформация, деленная на деформацию волокна (εx / εf), больше единицы, и усиление продольной скорости; кроме того, это возникающее усиление скорости может быть усилено изменяющейся архитектурной передачей за счет мезолатеральных и дорсовентральных изменений формы, как показано на перистая мышца схватки. Передаточное отношение красного к белому (красный εf / белый εf) отражает комбинированный эффект растяжения продольных красных мышечных волокон и косых белых мышечных волокон.[46][48]
Плавучесть


Тело рыбы плотнее воды, поэтому рыба должна компенсировать разницу, иначе она утонет. Много костлявые рыбы иметь внутренний орган, называемый плавательный пузырь, или газовый пузырь, который регулирует их плавучесть путем манипулирования газами. Таким образом, рыба может оставаться на текущей глубине воды, подниматься или опускаться, не тратя энергию на плавание. Мочевой пузырь есть только у костистых рыб. В более примитивных группах, как некоторые пескарей, бихиры и двоякодышащая рыба мочевой пузырь открыт для пищевод и вдвое больше легкое. Он часто отсутствует у быстро плавающих рыб, таких как семейства тунца и скумбрии. Состояние мочевого пузыря, открытого в пищевод, называется физостом, закрытое состояние физоклист. В последнем случае газосодержание мочевого пузыря регулируется через rete mirabilis, сеть кровеносных сосудов, влияющих на газообмен между мочевым пузырем и кровью.[49]
У некоторых рыб ретомирабиль наполняет плавательный пузырь кислородом. А система противоточного обмена используется между венозными и артериальными капиллярами. Понизив pH уровни в венозных капиллярах, кислород не связывает кровь гемоглобин. Это вызывает повышение концентрации кислорода в венозной крови, позволяя кислороду диффундировать через капиллярную мембрану в артериальные капилляры, где кислород все еще блокируется в гемоглобине. Цикл диффузии продолжается до тех пор, пока концентрация кислорода в артериальных капиллярах не станет равной. перенасыщенный (больше, чем концентрация кислорода в плавательном пузыре). В этот момент свободный кислород из артериальных капилляров диффундирует в плавательный пузырь через газовую железу.[50]
В отличие от костистых рыб, у акул нет плавучего пузыря, наполненного газом. Вместо этого акулы полагаются на большую печень, наполненную маслом, содержащим сквален, и их хрящ, который составляет примерно половину нормальной плотности кости.[51] Их печень составляет до 30% от общей массы их тела.[35] Эффективность печени ограничена, поэтому акулы используют динамический подъем для поддержания глубины, когда не плаваете. Песчаные тигровые акулы хранят воздух в желудке, используя его как плавательный пузырь. Большинству акул необходимо постоянно плавать, чтобы дышать, и они не могут долго спать, не утонув (если вообще). Однако некоторые виды, такие как акула-медсестра, способны перекачивать воду через жабры, позволяя им отдыхать на дне океана.[52]
Сенсорные системы
Большинство рыб обладают высокоразвитыми органами чувств. Почти все дневные рыбы имеют цветовое зрение, по крайней мере, такое же хорошее, как у человека (см. зрение у рыб ). У многих рыб есть хеморецепторы, отвечающие за необычные чувства вкуса и запаха. Хотя у них есть уши, многие рыбы могут плохо слышать. У большинства рыб есть чувствительные рецепторы, которые формируют система боковой линии, который обнаруживает слабые течения и вибрации, а также движение ближайших рыб и добычи.[53] Акулы могут воспринимать частоты в диапазоне от 25 до 50.Гц через их боковую линию.[54]
Рыбы ориентируются по ориентирам и могут использовать мысленные карты, основанные на нескольких ориентирах или символах. Поведение рыб в лабиринтах показывает, что они обладают пространственной памятью и зрительным различением.[55]
Зрение
Зрение это важный сенсорная система для большинства видов рыб. Рыбьи глаза похожи на глаза земной позвоночные подобно птицы и млекопитающие, но сферический линза. Их сетчатка обычно есть оба стержневые клетки и конические клетки (за скотопический и фотопическое зрение ), и у большинства видов цветовое зрение. Некоторые рыбы могут видеть ультрафиолетовый и некоторые могут видеть поляризованный свет. Среди рыба без челюсти, то минога имеет хорошо развитые глаза, а миксина имеет только примитивный пятна.[56] Шоу рыбного зрения приспособление к их визуальной среде, например глубоководные рыбы глаза подходят для темной среды.
Слух
Слух является важной сенсорной системой для большинства видов рыб. Порог слуха и способность локализовать источники звука снижаются под водой, где скорость звука выше, чем в воздухе. Подводный слух костная проводимость, а локализация звука, по-видимому, зависит от различий в амплитуде, обнаруживаемых костной проводимостью.[57] Однако у водных животных, таких как рыбы, есть более специализированный слуховой аппарат, который эффективен под водой.[58]
Рыба может чувствовать звук через боковые линии и их отолиты (уши). Некоторые рыбы, например, некоторые виды карп и сельдь, слышат через плавательный пузырь, который функционирует как слуховой аппарат.[59]
Слух хорошо развит в карп, которые имеют Веберовский орган, три специализированных позвоночных отростка, передающих колебания плавательного пузыря внутреннему уху.
Хотя слух акул проверить сложно, у них может быть острый чувство слуха и, возможно, слышит добычу за много миль.[60] Маленькие отверстия на каждой стороне головы (не дыхальце ) ведет прямо в внутреннее ухо через тонкий канал. В боковая линия показывает аналогичное расположение и открыт для окружающей среды через ряд отверстий, называемых боковой линией поры. Это напоминание об общем происхождении этих двух органов, обнаруживающих вибрацию и звук, которые сгруппированы вместе в акустико-латеральную систему. У костистой рыбы и четвероногие внешнее отверстие во внутреннее ухо потеряно.
Хеморецепция

Акулы увлеклись обонятельный органы чувств, расположенные в коротком протоке (который не слит, в отличие от костистых рыб) между передним и задним носовыми отверстиями, причем некоторые виды способны обнаружить всего лишь один часть на миллион крови в морской воде.[61]
У акул есть способность определять направление данного запаха в зависимости от времени обнаружения запаха каждой ноздрей.[62] Это похоже на метод, который используют млекопитающие для определения направления звука.
Их больше привлекают химические вещества, содержащиеся в кишечнике многих видов животных, и в результате они часто задерживаются рядом или в сточные воды водопады. Некоторые виды, такие как акулы-медсестры, иметь внешние усики которые значительно увеличивают их способность чувствовать добычу.
Магнитоцепция
Электрорецепция

У некоторых рыб, таких как сомы и акулы, есть органы, которые обнаруживают слабые электрические токи порядка милливольт.[63] Другая рыба, такая как южноамериканские электрические рыбы Gymnotiformes, могут создавать слабые электрические токи, которые они используют в навигации и социальных сетях. У акул ампулы Лоренцини являются электрорецепторными органами. Они исчисляются от сотен до тысяч. Акулы используют ампулы Лоренцини для обнаружения электромагнитные поля что производят все живые существа.[64] Это помогает акулам (особенно Акула-молот ) найти добычу. Акула обладает наибольшей электрической чувствительностью из всех животных. Акулы находят добычу, спрятанную в песке, обнаруживая электрические поля они производят. Океанские течения движется в магнитное поле Земли также генерируют электрические поля, которые акулы могут использовать для ориентации и, возможно, навигации.[65]
- В ампулы Лоренцини позволить акулам почувствовать электрические разряды.
- Электрическая рыба способны создавать электрические поля с помощью модифицированных мышц своего тела.
Боль
Эксперименты, проведенные Уильямом Таволгой, доказывают, что у рыб боль и бояться ответов. Например, в опытах Таволги жаба grunted when electrically shocked and over time they came to grunt at the mere sight of an electrode.[66]
In 2003, Scottish scientists at the Эдинбургский университет and the Roslin Institute concluded that rainbow trout exhibit behaviors often associated with боль у других животных. Пчела яд и уксусная кислота injected into the lips resulted in fish rocking their bodies and rubbing their lips along the sides and floors of their tanks, which the researchers concluded were attempts to relieve pain, similar to what mammals would do.[67][68][69] Neurons fired in a pattern resembling human neuronal patterns.[69]
Professor James D. Rose of the Университет Вайоминга claimed the study was flawed since it did not provide proof that fish possess "conscious awareness, particularly a kind of awareness that is meaningfully like ours".[70] Rose argues that since fish brains are so different from human brains, fish are probably not conscious in the manner humans are, so that reactions similar to human reactions to pain instead have other causes. Rose had published a study a year earlier arguing that fish cannot feel pain because their brains lack a неокортекс.[71] However, animal behaviorist Темпл Грандин утверждает, что рыбы все еще могут иметь сознание без неокортекса, потому что «разные виды могут использовать разные структуры и системы мозга для выполнения одних и тех же функций».[69]
Animal welfare advocates raise concerns about the possible страдания of fish caused by angling. Some countries, such as Germany have banned specific types of fishing, and the British RSPCA now formally prosecutes individuals who are cruel to fish.[72]
Reproductive processes
Оогония development in teleosts fish varies according to the group, and the determination of oogenesis dynamics allows the understanding of maturation and fertilisation processes. Изменения в ядро, ooplasm, and the surrounding layers characterize the oocyte maturation process.[73]
Postovulatory follicles are structures formed after oocyte release; they do not have эндокринный function, present a wide irregular lumen, and are rapidly reabsorbed in a process involving the апоптоз of follicular cells. A degenerative process called follicular atresia reabsorbs vitellogenic oocytes not spawned. This process can also occur, but less frequently, in oocytes in other development stages.[73]
Некоторые рыбы гермафродиты, having both testes and ovaries either at different phases in their life cycle or, as in деревушки, have them simultaneously.
Over 97% of all known fish are яйцекладущий,[74] that is, the eggs develop outside the mother's body. Examples of oviparous fish include лосось, золотая рыбка, цихлиды, тунец, и угри. In the majority of these species, fertilisation takes place outside the mother's body, with the male and female fish shedding their гаметы into the surrounding water. However, a few oviparous fish practice internal fertilisation, with the male using some sort of интромитентный орган to deliver sperm into the genital opening of the female, most notably the oviparous sharks, such as the horn shark, and oviparous rays, such as коньки. In these cases, the male is equipped with a pair of modified тазовый fins known as застежки.
Marine fish can produce high numbers of eggs which are often released into the open water column. The eggs have an average diameter of 1 millimetre (0.039 in). The eggs are generally surrounded by the extraembryonic membranes but do not develop a shell, hard or soft, around these membranes. Some fish have thick, leathery coats, especially if they must withstand physical force or desiccation. These type of eggs can also be very small and fragile.

Яйцо минога

Яйцо catshark (mermaids' purse )

Яйцо bullhead shark

Яйцо химера

The newly hatched young of oviparous fish are called личинки. They are usually poorly formed, carry a large желточный мешок (for nourishment) and are very different in appearance from juvenile and adult specimens. The larval period in oviparous fish is relatively short (usually only several weeks), and larvae rapidly grow and change appearance and structure (a process termed метаморфоза ) to become juveniles. During this transition larvae must switch from their yolk sac to feeding on зоопланктон prey, a process which depends on typically inadequate zooplankton density, starving many larvae.
В яйцекладущие fish the eggs develop inside the mother's body after internal fertilisation but receive little or no nourishment directly from the mother, depending instead on the желток. Each embryo develops in its own egg. Familiar examples of ovoviviparous fish include гуппи, angel sharks, и coelacanths.
Some species of fish are живородящий. In such species the mother retains the eggs and nourishes the embryos. Typically, viviparous fish have a structure analogous to the плацента видел в млекопитающие connecting the mother's blood supply with that of the embryo. Examples of viviparous fish include the surf-perches, splitfins, и лимонная акула. Some viviparous fish exhibit оофагия, in which the developing embryos eat other eggs produced by the mother. This has been observed primarily among sharks, such as the короткопёрый мако и сельдевая акула, but is known for a few bony fish as well, such as the полуклюв Nomorhamphus ebrardtii.[75] Intrauterine cannibalism is an even more unusual mode of vivipary, in which the largest embryos eat weaker and smaller siblings. This behavior is also most commonly found among sharks, such as the grey nurse shark, but has also been reported for Nomorhamphus ebrardtii.[75]
In many species of fish, fins have been modified to allow Внутреннее оплодотворение.
Аквариумисты commonly refer to ovoviviparous and viviparous fish as livebearers.
- Many fish species are гермафродиты. Synchronous hermaphrodites possess both яичники и яички в то же время. Последовательные гермафродиты have both types of tissue in their гонады, with one type being predominant while the fish belongs to the corresponding gender.
Социальное поведение
Fish social behaviour called ‘shoaling’ involves a group of fish swimming together. This behaviour is a defence mechanism in the sense that there is safety in large numbers, where chances of being eaten by predators are reduced. Shoaling also increases mating and foraging success. Schooling on the other hand, is a behaviour within the shoal where fish can be seen performing various manoeuvres in a synchronised manner.[76] The parallel swimming is a form of ‘social copying’ where fish in the school replicate the direction and velocity of its neighbouring fishes.[77]
Experiments done by D.M. Steven, on the shoaling behaviour of fish concluded that during the day, fish had a higher tendency to stay together as a result of a balance between single fish leaving and finding their own direction and the mutual attraction between fishes of the same species. It was found that at night the fish swam noticeably faster however, often singly and in no co-ordination. Groups of two or three could be seen frequently formed although were dispersed after a couple of seconds.[78]
Theoretically, the amount of time that a fish stays together in a shoal should represent their cost of staying instead of leaving.[79] A past laboratory experiment done on cyprinids has established that the time budget for social behaviour within a shoal varies proportionally to the quantity of fishes present. This originates from the cost/benefit ratio which changes accordingly with group size, measured by the risk of predation versus food intake.[80] When the cost/benefit ratio is favourable to shoaling behaviour then decisions to stay with a group or join one is favourable. Depending on this ratio, fish will correspondingly decide to leave or stay. Thus, shoaling behaviour is considered to be driven by an individual fish's constant stream of decisions.[76]
Смотрите также
Рекомендации
- ^ Schwab, Ivan R (2002). "More than just cool shades". British Journal of Ophthalmology. 86 (10): 1075. Дои:10.1136/bjo.86.10.1075. ЧВК 1771297. PMID 12349839.
- ^ Prosser, C. Ladd (1991). Сравнительная физиология животных, экологическая и метаболическая физиология животных (4-е изд.). Хобокен, Нью-Джерси: Wiley-Liss. С. 1–12. ISBN 978-0-471-85767-9.
- ^ Хоар WS и Рэндалл ди-джей (1984) Физиология рыб: Жабры: Часть A - Анатомия, газообмен и кислотно-щелочная регуляция Академическая пресса. ISBN 978-0-08-058531-4.
- ^ Хоар WS и Рэндалл ди-джей (1984) Физиология рыб: Жабры: Часть B - Перенос ионов и воды Академическая пресса. ISBN 978-0-08-058532-1.
- ^ а б Армбрустер, Джонатан В. (1998). «Модификации пищеварительного тракта для удержания воздуха у лорикариевых и сколоплацидных сомов» (PDF). Копея. 1998 (3): 663–675. Дои:10.2307/1447796. JSTOR 1447796. Получено 25 июн 2009.
- ^ а б Campbell, Neil A. (1990). Биология (Второе изд.). Redwood City, California: Benjamin/Cummings Publishing Company, Inc. pp.836 –838. ISBN 0-8053-1800-3.
- ^ а б Hughes GM (1972). "Morphometrics of fish gills". Физиология дыхания. 14 (1–2): 1–25. Дои:10.1016/0034-5687(72)90014-x. PMID 5042155.
- ^ а б c Storer, Tracy I.; Usinger, R. L.; Stebbins, Robert C.; Nybakken, James W. (1997). Общая зоология (шестое изд.). Нью-Йорк: Макгроу-Хилл. стр.668 –670. ISBN 0-07-061780-5.
- ^ Скотт, Томас (1996). Краткая энциклопедия биологии. Вальтер де Грюйтер. п.542. ISBN 978-3-11-010661-9.
- ^ Эндрюс, Крис; Адриан Экселл; Невилл Кэррингтон (2003). Руководство по здоровью рыб. Книги Светлячка.
- ^ а б c d е ж грамм час я j k л м п о Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного. Филадельфия, Пенсильвания: Holt-Saunders International. С. 316–327. ISBN 978-0-03-910284-5.
- ^ Rovainen, 1985
- ^ Waldrop, Iwamoto and Haouzi, 2006
- ^ Сарский, Хенрик (1957). «Происхождение личинки и метаморфозы у амфибии». Американский натуралист. Институт Эссекса. 91 (860): 287. Дои:10.1086/281990. JSTOR 2458911.
- ^ Клак, Дж. А. (2002): Завоевание позиций: происхождение и эволюция четвероногих. Издательство Индианского университета, Блумингтон, Индиана. 369 стр.
- ^ Gilbertson, Lance (1999). Руководство зоологической лаборатории. New York: McGraw-Hill Companies, Inc. ISBN 978-0-07-237716-3.
- ^ William J. Bennetta (1996). "Deep Breathing". Архивировано из оригинал 14 августа 2007 г.. Получено 28 августа 2007.
- ^ "SHARKS & RAYS, SeaWorld/Busch Gardens ANIMALS, CIRCULATORY SYSTEM". Busch Entertainment Corporation. Архивировано из оригинал 24 апреля 2009 г.. Получено 3 сентября 2009.
- ^ Лаурин М. (1998): Важность глобальной экономичности и исторической предвзятости в понимании эволюции четвероногих. Часть I. Систематика, эволюция среднего уха и подвешивание челюсти. Annales des Sciences Naturelles, Zoologie, Париж, 13e Série 19: pp 1-42.
- ^ Setaro, John F. (1999). Circulatory System. Microsoft Encarta 99.
- ^ Guillaume, Jean; Praxis Publishing; Sadasivam Kaushik; Pierre Bergot; Robert Metailler (2001). Nutrition and Feeding of Fish and Crustaceans. Springer. п. 31. ISBN 978-1-85233-241-9. Получено 9 января 2009.
- ^ Национальный институт рака (2 февраля 2011 г.). "NCI Dictionary of Cancer Terms — large intestine". Получено 16 сентября 2012.
- ^ а б Мартин, Р. Эйдан. "Кто не рискует, тот не пьет шампанское". Центр исследования акул ReefQuest. Получено 22 августа 2009.
- ^ Reddon, AR; O'Connor, CM; Marsh-Rollo, SE; Balcshine, S (2012). "Effects of isotocin on social responses in a cooperatively breeding fish". Поведение животных. 84 (4): 753–760. Дои:10.1016/j.anbehav.2012.07.021. S2CID 13037173.
- ^ Fish Version Of Oxytocin Drives Their Social Behavior Наука 2.0, 10 октября 2012 г.
- ^ Pinto, Patricia; Estevao, Maria; Power, Deborah (August 2014). "Effects of Estrogens and Estrogenic Disrupting Compounds on Fish Mineralized Tissues". Морские препараты. 12 (8): 4474–94. Дои:10.3390/md12084474. ЧВК 4145326. PMID 25196834.
- ^ Waterman, Jim. "Research into wastewater treatment effluent impact on Boulder Creek fish sexual development". Boulder Area Sustainability Information Network.
- ^ Arukwe, Augustine (August 2001). "Cellular and Molecular Responses to Endocrine-Modulators and The Impact on Fish Reproduction". Бюллетень загрязнения морской среды. 42 (8): 643–655. Дои:10.1016/S0025-326X(01)00062-5. PMID 11525282.
- ^ а б Reeves, C (2015). "Of Frogs & Rhetoric: The Atrazine wars". Ежеквартальная техническая коммуникация. 24 (4): 328–348. Дои:10.1080/10572252.2015.1079333. S2CID 53626955.
- ^ Suter, Glenn; Cormier, Susan (2015). "The problem of biased data and potential solutions for health and environmental assessments". Оценка рисков для человека и окружающей среды: международный журнал. 21 (7): 1–17. Дои:10.1080/10807039.2014.974499. S2CID 84723794.
- ^ Rohr, J.R. (2018) "Atrazine and Amphibians: A Story of Profits, Controversy, and Animus". In: D. A. DellaSala, and M. I. Goldstein (eds.) Encyclopedia of the Anthropocene, volume 5, pages 141–148. Оксфорд: Эльзевир.
- ^ Fowler, Sarah; Cavanagh, Rachel. "Sharks, Rays, and Chimaeras: The Status of Chondrichthyan Fishes" (PDF). Всемирный союз охраны природы. Получено 3 мая 2017.
- ^ Arukwe, Augustine (2001). "Cellular and Molecular Responses to Endocrine-Modulators and the Impact on Fish Reproduction". Бюллетень загрязнения морской среды. 42 (8): 643–655. Дои:10.1016/s0025-326x(01)00062-5. PMID 11525282.
- ^ Chemistry of the body fluids of the coelacanth, Latimeria chalumnae
- ^ а б Компаньо, Леонард; Dando, Marc; Fowler, Sarah (2005). Sharks of the World. Collins Field Guides. ISBN 978-0-00-713610-0. OCLC 183136093.
- ^ John A. Musick (2005). "Management techniques for elasmobranch fisheries: 14. Shark Utilization". FAO: Fisheries and Aquaculture Department. Получено 16 марта 2008.
- ^ Thomas Batten. "MAKO SHARK Isurus oxyrinchus". Delaware Sea Grant, University of Delaware. Архивировано из оригинал 11 марта 2008 г.. Получено 16 марта 2008.
- ^ Уилмер, Пэт; Стоун, Грэм; Джонстон, Ян (2009). Экологическая физиология животных. Вайли. стр.198–200. ISBN 978-1-4051-0724-2.
- ^ Martin, R. Aidan (April 1992). "Fire in the Belly of the Beast". Центр исследования акул ReefQuest. Получено 21 августа 2009.
- ^ а б c Sepulveda, C.A.; Dickson, K.A.; Bernal, D.; Graham, J. B. (1 July 2008). "Elevated red myotomal muscle temperatures in the most basal tuna species, Аллотуннус фаллай" (PDF). Журнал биологии рыб. 73 (1): 241–249. Дои:10.1111/j.1095-8649.2008.01931.x. Архивировано из оригинал (PDF) 7 февраля 2013 г.. Получено 2 ноября 2012.
- ^ а б "Tuna - Biology Of Tuna". Получено 12 сентября 2009.
- ^ а б Landeira-Fernandez, A.M.; Morrissette, J.M.; Blank, J.M.; Block, B.A. (16 October 2003). "Temperature dependence of the Ca2+-ATPase (SERCA2) in the ventricles of tuna and mackerel". AJP: Regulatory, Integrative and Comparative Physiology. 286 (2): 398R–404. Дои:10.1152/ajpregu.00392.2003. PMID 14604842.
- ^ Block, BA; Finnerty, JR (1993). "Endothermy in fishes: a phylogenetic analysis of constraints, predispositions, and selection pressures" (PDF). Экологическая биология рыб. 40 (3): 283–302. Дои:10.1007/BF00002518. S2CID 28644501.[постоянная мертвая ссылка ]
- ^ David Fleshler(10-15-2012) Южная Флорида Sun-Sentinel,
- ^ Kapoor 2004
- ^ а б Brainerd, E. L.; Azizi, E. (2005). "Muscle Fiber Angle, Segment Bulging and Architectural Gear Ratio in Segmented Musculature". Журнал экспериментальной биологии. 208 (17): 3249–3261. Дои:10.1242/jeb.01770. PMID 16109887.
- ^ а б c Azizi, E.; Brainerd, E. L.; Roberts, T. J. (2008). "Variable Gearing in Pennate Muscles". PNAS. 105 (5): 1745–1750. Bibcode:2008PNAS..105.1745A. Дои:10.1073/pnas.0709212105. ЧВК 2234215. PMID 18230734.
- ^ Brainerd, E. L.; Azizi, E. (2007). "Architectural Gear Ratio and Muscle Fiber Strain Homogeneity in Segmented Musculature". Журнал экспериментальной зоологии. 307 (A): 145–155. Дои:10.1002/jez.a.358. PMID 17397068.
- ^ Kardong, K. (2008). Vertebrates: Comparative anatomy, function, evolution (5-е изд.). Бостон: Макгроу-Хилл. ISBN 978-0-07-304058-5.
- ^ Kardong, K. (2008). Vertebrates: Comparative anatomy, function, evolution, (5th ed.). Бостон: Макгроу-Хилл.
- ^ Мартин, Р. Эйдан. «Важность хрящевой ткани». Центр исследования акул ReefQuest. Получено 29 августа 2009.
- ^ "Do sharks sleep". Flmnh.ufl.edu. Получено 23 сентября 2010.
- ^ Orr, James (1999). Рыбы. Microsoft Encarta 99. ISBN 978-0-8114-2346-5.
- ^ Popper, A.N.; C. Platt (1993). "Inner ear and lateral line". Физиология рыб. CRC Press (1st ed).
- ^ Journal of Undergraduate Life Sciences. "Appropriate maze methodology to study learning in fish" (PDF). Архивировано из оригинал (PDF) 6 июля 2011 г.. Получено 28 мая 2009.
- ^ N. A. Campbell и J. B. Reece (2005). Биология, Seventh Edition. Benjamin Cummings, San Francisco, California.
- ^ Shupak A. Sharoni Z. Yanir Y. Keynan Y. Alfie Y. Halpern P. (January 2005). "Underwater Hearing and Sound Localization with and without an Air Interface". Отология и невротология. 26 (1): 127–130. Дои:10.1097/00129492-200501000-00023. PMID 15699733. S2CID 26944504.
- ^ Graham, Michael (1941). "Sense of Hearing in Fishes". Природа. 147 (3738): 779. Bibcode:1941Natur.147..779G. Дои:10.1038/147779b0. S2CID 4132336.
- ^ Williams, C (1941). "Sense of Hearing in Fishes". Природа. 147 (3731): 543. Bibcode:1941Natur.147..543W. Дои:10.1038/147543b0. S2CID 4095706.
- ^ Мартин, Р. Эйдан. "Hearing and Vibration Detection". Получено 1 июня 2008.
- ^ Мартин, Р. Эйдан. "Smell and Taste". Центр исследования акул ReefQuest. Получено 21 августа 2009.
- ^ The Function of Bilateral Odor Arrival Time Differences in Olfactory Orientation of Sharks В архиве 8 марта 2012 г. Wayback Machine, Jayne M. Gardiner, Jelle Atema, Current Biology - 13 July 2010 (Vol. 20, Issue 13, pp. 1187-1191)
- ^ Albert, J.S., and W.G.R. Crampton. 2005. Electroreception and electrogenesis. pp. 431–472 in The Physiology of Fishes, 3rd Edition. D.H. Evans and J.B. Claiborne (eds.). CRC Press.
- ^ Kalmijn AJ (1982). "Electric and magnetic field detection in elasmobranch fishes". Наука. 218 (4575): 916–8. Bibcode:1982Sci...218..916K. Дои:10.1126/science.7134985. PMID 7134985.
- ^ Meyer CG, Holland KN, Papastamatiou YP (2005). "Sharks can detect changes in the geomagnetic field". Journal of the Royal Society, Interface. 2 (2): 129–30. Дои:10.1098/rsif.2004.0021. ЧВК 1578252. PMID 16849172.
- ^ Dunayer, Joan, "Fish: Sensitivity Beyond the Captor's Grasp," The Animals' Agenda, July/August 1991, pp. 12–18
- ^ Vantressa Brown, "Fish Feel Pain, British Researchers Say," Agence France-Presse, 1 May 2003
- ^ Kirby, Alex (30 April 2003). "Рыбы действительно чувствуют боль, - говорят ученые". Новости BBC. Получено 4 января 2010.
- ^ а б c Grandin, Temple; Джонсон, Кэтрин (2005). Животные в переводе. New York, New York: Scribner. стр.183–184. ISBN 978-0-7432-4769-6.
- ^ "Rose, J.D. 2003. A Critique of the paper: "Do fish have nociceptors: Evidence for the evolution of a vertebrate sensory system"" (PDF). Архивировано из оригинал (PDF) 8 июня 2011 г.. Получено 21 мая 2011.
- ^ James D. Rose, Чувствуют ли рыбы боль? В архиве 20 января 2013 г. Wayback Machine, 2002. Retrieved 27 September 2007.
- ^ Leake, J. "Anglers to Face RSPCA Check," The Sunday Times – Britain, 14 March 2004
- ^ а б Guimaraes-Cruz, Rodrigo J., Rodrigo J.; Santos, José E. dos; Santos, Gilmar B. (July–September 2005). "Gonadal structure and gametogenesis of Loricaria lentiginosa Isbrücker (Pisces, Teleostei, Siluriformes)". Rev. Bras. Zool. 22 (3): 556–564. Дои:10.1590/S0101-81752005000300005. ISSN 0101-8175.
- ^ Peter Scott: Живородящие рыбы, п. 13. Tetra Press 1997. ISBN 1-56465-193-2
- ^ а б Meisner, A.; Burns, J. (December 1997). "Viviparity in the Halfbeak Genera Дермогенис и Nomorhamphus (Teleostei: Hemiramphidae)". Журнал морфологии. 234 (3): 295–317. Дои:10.1002/(SICI)1097-4687(199712)234:3<295::AID-JMOR7>3.0.CO;2-8. PMID 29852651.
- ^ а б Pitcher, T.J. (1998). Shoaling and Schooling in Fishes. Garland, New York, USA: Greenberg, G. and Hararway. М.М. pp. 748–760.
- ^ Pitcher, T.J. (1983). "Heuristic definitions of fish shoaling behaviour". Поведение животных. 31 (2): 611–613. Дои:10.1016/S0003-3472(83)80087-6. S2CID 53195091.
- ^ Steven, D.M. (1959). "Studies on the shoaling behaviour of fish: I. Responses of two species to changes of illumination and to olfactory stimuli". Журнал экспериментальной биологии. 36 (2): 261–280.
- ^ Partridge, B.L. (1982). "Rigid definitions of schooling behaviour are inadequate". Поведение животных. 30: 298–299. Дои:10.1016/S0003-3472(82)80270-4. S2CID 53170935.
- ^ Pitcher, T.J (1982). "Evidence for position preferences in schooling mackerel". Поведение животных. 30 (3): 932–934. Дои:10.1016/S0003-3472(82)80170-X. S2CID 53186411.
дальнейшее чтение
- Bernier NJ, Van Der Kraak G, Farrell AP and Brauner CJ (2009) Fish Physiology: Fish Neuroendocrinology Академическая пресса. ISBN 978-0-08-087798-3.
- Eddy FB and Handy RD (2012) Ecological and Environmental Physiology of Fishes Издательство Оксфордского университета. ISBN 978-0-19-954095-2.
- Evans DH, JB Claiborne and S Currie (Eds) (2013) Физиология рыб 4th edition, CRC Press. ISBN 978-1-4398-8030-2.
- Grosell M, Farrell AP and Brauner CJ (2010) Fish Physiology: The Multifunctional Gut of Fish Академическая пресса. ISBN 978-0-08-096136-1.
- Hara TJ and Zielinski B (2006) Fish Physiology: Sensory Systems Neuroscience Академическая пресса. ISBN 978-0-08-046961-4.
- Kapoor BG and Khanna B (2004) "Ichthyology handbook" Pages 137–140, Springer. ISBN 978-3-540-42854-1.
- McKenzie DJ, Farrell AP and Brauner CJ (2007) Физиология рыб: примитивные рыбы Академическая пресса. ISBN 978-0-08-054952-1.
- Sloman KA, Wilson RW and Balshine S (2006) Behaviour And Physiology of Fish Gulf Professional Publishing. ISBN 978-0-12-350448-7.
- Wood CM, Farrell AP and Brauner CJ (2011) Fish Physiology: Homeostasis and Toxicology of Non-Essential Metals Академическая пресса. ISBN 978-0-12-378634-0.
внешняя ссылка
Физиология типы | ||
|---|---|---|
| Животные |  | |
| Растения | ||
| Клетки | ||
| похожие темы | ||
| О рыбы |  | ||||
|---|---|---|---|---|---|
| Анатомия и физиология | |||||
| Сенсорный системы | |||||
| Размножение | |||||
| Передвижение | |||||
| Другой поведение | |||||
| К среда обитания | |||||
| Другие типы | |||||
| Коммерческий |
| ||||
| Основные группы | |||||
| Списки | |||||
| |||||