Мегафауна - Megafauna

В земном зоология, то мегафауна (от Греческий μέγας мегас "большой" и Новая латынь фауна «животный мир») включает крупных или гигантских животных определенного района, среды обитания или геологического периода. Чаще всего используются пороговые значения веса более 40 кг (90 фунтов).[1] или 44 килограмма (100 фунтов)[2][3] (т.е. имеющий массу, сравнимую или превышающую массу человек ) или более тонна, 1000 кг (2205 фунтов)[1][4][5] (т.е. имеющий массу, сравнимую или превышающую массу бык ). К первым из них относятся многие виды, которые обычно не считаются слишком большими, например белохвостый олень и красный кенгуру.
На практике наиболее распространенное использование, встречающееся в академической и популярной литературе, описывает наземных млекопитающих примерно крупнее человека, которые не (исключительно) одомашнены. Этот термин особенно ассоциируется с Мегафауна плейстоцена - наземные животные часто крупнее современных собратьев, считающихся типичными последний ледниковый период, такие как мамонты, большинство из которых в Северной Евразии, Америке и Австралия вымерли за последние сорок тысяч лет.[6] Среди живых животных термин мегафауна чаще всего используется для обозначения самых крупных животных. сохранившийся наземные млекопитающие, которые слоны, жирафы, бегемоты, носороги, и большой коровы. Из этих пяти категорий крупных травоядных только коровы в настоящее время находятся за пределами Африка и южный Азия, но все остальные раньше были более широкими. Виды мегафауны можно разделить на категории в соответствии с их типом питания: мегафауны (например, слоны ), мегакароядные (например, львы ), реже - мегаядные (например, медведи ).
Другое распространенное использование - для гигантских водных видов, особенно киты, любые крупные дикие или одомашненные наземные животные, например, более крупные антилопа и крупный рогатый скот, а также динозавры и другие вымершие гигантские рептилии.
Термин мегафауна очень редко используется для описания беспозвоночных, хотя иногда он использовался для некоторых видов вымерших беспозвоночных, которые были намного крупнее, чем все аналогичные виды беспозвоночных, существующих сегодня, например стрекозы высотой 1 м (3 фута) из Каменноугольный период.
Экологическая стратегия
Мегафауна - в смысле крупнейших млекопитающих и птиц - обычно K-стратеги, с высокой продолжительностью жизни, медленными темпами роста популяции, низким уровнем смертности и (по крайней мере, для самых крупных) небольшим количеством естественных хищников, способных убивать взрослых особей, или отсутствием их вовсе.[7] Эти характеристики, хотя и не исключительные для такой мегафауны, делают их уязвимыми для человека. чрезмерная эксплуатация отчасти из-за медленных темпов восстановления популяции.[8][9]
Эволюция большого размера тела
Одно наблюдение, которое было сделано в отношении эволюции увеличения размера тела, заключается в том, что быстрые темпы увеличения, которые часто наблюдаются в течение относительно коротких интервалов времени, не являются устойчивыми в течение более длительных периодов времени. При исследовании изменений массы тела млекопитающих с течением времени было обнаружено, что максимальное увеличение, возможное в данном временном интервале, масштабируется с увеличением длины интервала до 0,25 степени.[10] Считается, что это отражает появление во время тенденции к увеличению максимального размера тела ряда анатомических, физиологических, экологических, генетических и других ограничений, которые должны быть преодолены эволюционными инновациями, прежде чем станет возможным дальнейшее увеличение размеров. Поразительно более высокая скорость изменений была обнаружена при значительном уменьшении массы тела, что может быть связано с феноменом островная карликовость. При нормировании на длину поколения было обнаружено, что максимальная скорость уменьшения массы тела более чем в 30 раз превышает максимальную скорость увеличения массы тела при десятикратном изменении.[10]
У наземных млекопитающих

После Меловое – палеогеновое вымирание это уничтожило нептичьих динозавров около 66 Ма (миллион лет) назад размер тела наземных млекопитающих увеличивался почти экспоненциально, поскольку они диверсифицировались, чтобы занять экологические ниши, оставшиеся вакантными.[11] Начиная с нескольких кг до события, максимальный размер достиг ~ 50 кг несколько миллионов лет спустя и ~ 750 кг к концу Палеоцен. Эта тенденция к увеличению массы тела, по-видимому, стабилизируется около 40 млн лет назад (в конце эоцен ), предполагая, что физиологические или экологические ограничения были достигнуты после увеличения массы тела более чем на три порядка.[11] Однако, если рассматривать с точки зрения скорости увеличения размера в расчете на поколение, экспоненциальное увеличение продолжалось до появления Индрикотерий 30 млн лет назад. (Поскольку время генерации масштабируется с масса тела0.259, увеличение времени генерации с увеличением размера приводит к тому, что график зависимости массы бревна от времени изгибается вниз от линейной аппроксимации.)[10]
В конце концов, мегагербоядные животные достигли массы более 10 000 кг. Самый большой из них, индрикотеры и хоботки, Был ферментеры задней кишки, которые, как считается, имеют преимущество перед ферментеры передней кишки с точки зрения способности ускорить прохождение через желудочно-кишечный тракт, чтобы приспособиться к очень большому потреблению пищи.[12] Аналогичная тенденция возникает, когда темпы увеличения максимальной массы тела за поколение у разных млекопитающих клады сравниваются (с использованием ставок, усредненных по макроэволюционный шкалы времени). Среди наземных млекопитающих самые быстрые темпы роста масса тела0.259 от времени (в млн лет) произошло в периссодактилии (наклон 2,1), за которым следует грызуны (1.2) и хоботки (1.1),[10] все из которых являются ферментерами заднего кишечника. Скорость увеличения для парнокопытные (0,74) было примерно втрое меньше, чем у периссодактилей. Ставка на плотоядные (0,65) был еще немного ниже, а приматы, возможно, сдерживаются их древесный привычек, имел самый низкий показатель (0,39) среди изученных групп млекопитающих.[10]
Плотоядные наземные млекопитающие из нескольких евтерский группы ( парнокопытный Эндрюсарх - ранее считался мезонихид, то оксианид Саркастодон, и хищники Амфицион и Arctodus ) все достигли максимального размера около 1000 кг[11] (карниворан Арктотерий и гиенодонтид Симбакубва может быть несколько больше). Самый крупный из известных метатериан плотоядное животное Проборхьяна гигантея, видимо, достигла 600 кг, что тоже близко к этому пределу.[13] Аналогичный теоретический максимальный размер для плотоядных млекопитающих был предсказан на основе скорости метаболизма млекопитающих, энергетических затрат на добычу добычи и максимального оценочного коэффициента потребления добычи.[14] Также было высказано предположение, что максимальный размер хищных млекопитающих ограничен стрессом, который плечевая кость выдерживает максимальную скорость бега.[13]
Анализ изменения максимального размера тела за последние 40 млн лет показывает, что снижение температуры и увеличение площади континентальной суши связаны с увеличением максимального размера тела. Первая корреляция будет соответствовать Правило Бергмана,[15] и может быть связано с терморегулирующий преимущество большой массы тела в прохладном климате,[11] лучшая способность более крупных организмов справляться с сезонностью пищевых продуктов,[15] или другие факторы;[15] Последняя корреляция может быть объяснена с точки зрения диапазона и ограничений ресурсов.[11] Однако эти два параметра взаимосвязаны (из-за падения уровня моря, сопровождающего усиление оледенения), что затрудняет определение движущей силы тенденций максимального размера.[11]
У морских млекопитающих

Поскольку четвероногие (первые рептилии, позже млекопитающие ) вернулись в море в поздней перми, они доминировали в верхней части диапазона размеров морских организмов из-за более эффективного поглощения кислорода, возможного с помощью легких.[16][17] Предки китообразные считается полуводным пакицетиды, не крупнее собак, около 53 миллионов лет назад.[18] К 40 млн лет назад китообразные достигли в длину 20 м и более. Базилозавр, удлиненный змеевидный кит, который во многих отношениях отличался от современных китов и не был их предком. После этого эволюция крупных размеров тела у китообразных, по-видимому, временно остановилась, а затем откатилась назад, хотя доступные летописи окаменелостей ограничены. Однако в период с 31 млн. Лет назад (в Олигоцен ) до настоящего времени у китообразных наблюдается значительно более быстрое устойчивое увеличение массы тела (скорость увеличения масса тела0.259 в 3,2 раза на миллион лет), чем достигается любой группой наземных млекопитающих.[10] Эта тенденция привела к появлению самого большого животного всех времен, современного синий кит. Возможны несколько причин более быстрой эволюции крупных размеров тела у китообразных. Меньше биомеханический ограничения на увеличение размеров тела могут быть связаны с подвешиванием в воде, а не противостоянием силе тяжести, и с плавательные движения в отличие от наземное движение. Кроме того, большая теплоемкость и теплопроводность воды по сравнению с воздухом может увеличить терморегулирующий преимущество большого размера тела в морской эндотермы, хотя применяется убывающая отдача.[10]
У зубатых китов максимальный размер тела ограничен доступностью пищи. Больший размер, как в сперма и клюв китов, облегчает более глубокое погружение для доступа к относительно легко пойманной крупной добыче головоногих моллюсков в менее конкурентной среде. По сравнению с зубатыми китами эффективность усатых китов фильтрующая подача когда планктонная пища густая, она более выгодно масштабируется при увеличении размера, что делает более крупный размер более предпочтительным. В выпад кормления техника Rorquals оказывается более энергоэффективным, чем кормление барана из балаенид киты; последний метод используется с менее плотным и неоднородным планктоном.[19] Тенденция к похолоданию в новейшей истории Земли, возможно, привела к появлению большего количества мест с высоким содержанием планктона за счет ветряных апвеллинги, способствуя эволюции гигантских китов.[19]
Китообразные - не единственные морские млекопитающие, достигающие огромных размеров. Самый большой плотоядные всех времен морские ластоногие, крупнейшим из которых является южный морской слон, который может достигать 6 метров в длину и весить до 5 000 кг (11 000 фунтов). Другие крупные ластоногие включают северный морской слон при 4000 кг (8800 фунтов), морж при 2000 кг (4400 фунтов), и Морской лев Стеллер при 1135 кг (2502 фунта). В сирены - еще одна группа морских млекопитающих, которые адаптировались к полноценной водной жизни примерно в то же время, что и китообразные. Сирены тесно связаны со слонами. Самой большой сиреной была Стеллерова морская корова, который достигал 10 метров в длину и весил от 8000 до 10000 килограммов (от 18000 до 22000 фунтов) и был истреблен в 18 веке. Полуводный бегемот, которое является наземным млекопитающим, наиболее близким к китообразным, может достигать 3200 килограммов (7 100 фунтов).
У нелетающих птиц

Из-за небольшого начального размера всех млекопитающих после вымирания нептичьих динозавров у позвоночных, не являющихся млекопитающими, было окно возможностей продолжительностью около десяти миллионов лет (в течение палеоцена) для эволюции гигантизма без особой конкуренции.[20] В течение этого интервала хищник вершины ниши часто занимали рептилии, например, наземные крокодилы (например. Pristichampsus ), большие змеи (например, Титанобоа ) или варанидские ящерицы, или нелетающими птицами[11] (например. Палеопсилоптер В Южной Америке). Это также период, когда мегафауна нелетающие травоядные гасторнитид птицы эволюционировали в Северном полушарии, а нелетающие палеогнаты развился до больших размеров на Гондванец суши и Европа. Гасторнитиды и по крайней мере одна линия нелетающих палеогнатных птиц произошли в Европе, причем обе линии доминируют в нишах крупных травоядных, в то время как вес млекопитающих остается ниже 45 кг (в отличие от других массивов суши, таких как Северная Америка и Азия, которые видели более раннюю эволюцию более крупных млекопитающих) и были самыми крупными европейскими четвероногими в Палеоцен.[21]
Нелетающие палеогнаты, получившие название ратиты, традиционно рассматривались как представители рода, отличного от линии их маленьких летучих родственников, Неотропный жестяной. Однако недавние генетические исследования показали, что оловянные гнезда хорошо гнездятся внутри ратитового дерева и являются сестринская группа вымерших моа Новой Зеландии.[20][22][23] Точно так же малые киви Новой Зеландии были признаны сестринской группой вымерших слон птицы Мадагаскара.[20] Эти данные показывают, что нелетающая способность и гигантизм возникал независимо несколько раз среди ратитов через параллельная эволюция.
Хищные нелетающие птицы мегафауны часто могли конкурировать с млекопитающими в ранние годы жизни. Кайнозойский. Позже, в кайнозое, они были вытеснены развитыми хищниками и вымерли. В Северной Америке баторнитиды Паракракс и Баторнис были высшими хищниками, но вымерли из-за Ранний миоцен. В Южной Америке родственные форусрациды разделял доминирующие хищные ниши с метатерией Спарассодонты на протяжении большей части кайнозоя, но сократилась и в конечном итоге вымерла после того, как эвтерианские хищники прибыли из Северной Америки (как часть Великий американский обмен ) в течение Плиоцен. Напротив, до наших дней сохранились крупные травоядные нелетающие ратиты.
Однако ни одна из нелетающих птиц кайнозоя, в том числе и хищная Бронторнис, возможно всеядный Дроморнис[24] или травоядные Воромбе, когда-либо вырастали до массы, намного превышающей 500 кг, и, таким образом, никогда не достигали размеров крупнейших хищных млекопитающих, не говоря уже о размерах крупнейших травоядных млекопитающих. Было высказано предположение, что увеличение толщины птичьей яичной скорлупы пропорционально массе яйца с увеличением размера яйца накладывает верхний предел на размер птицы.[25][примечание 1] Самый крупный вид Дроморнис, Д. стиртони, возможно, вымерли после того, как достигли максимальной массы тела птицы, а затем уступили место сумчатым дипротодонты которые выросли до размеров в несколько раз больше.[28]
В гигантских черепахах
Гигантские черепахи были важными компонентами в последнее время Кайнозойский мегафауны, присутствовавшие на всех неполярных континентах до прибытия гоминины.[29][30] Самая крупная из известных наземных черепах была Атлас мегалохели, животное, которое, вероятно, весило около 1000 кг.
Некоторые более ранние водные Testudines, например морской Архелон мелового периода и пресноводных Ступендемис миоцена, были значительно крупнее и весили более 2000 кг.
Массовые вымирания мегафауны
Сроки и возможные причины
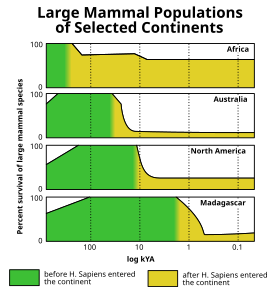
В Голоценовое вымирание (смотрите также Событие четвертичного вымирания ), произошедший в конце последний ледниковый период ледниковый период (он же Вюрмское оледенение ) когда многие гигантские млекопитающие ледникового периода, такие как шерстистые мамонты, вымерли в Америка и северный Евразия. Анализ случая вымирания в Северной Америке показал, что оно является уникальным среди кайнозойских импульсов вымирания по своей избирательности для крупных животных.[31](Рис. 10) Различные теории связывают волну вымирания с человеческая охота, изменение климата, болезнь, а предполагаемое внеземное столкновение, или по другим причинам. Однако это вымирание ближе к концу Плейстоцен был лишь одним из серии импульсов мегафауны вымирания, которые произошли в течение последних 50 000 лет на большей части поверхности Земли, с Африка и южный Азия (где местная мегафауна имела шанс развиваться вместе с современными людьми) пострадала сравнительно меньше. Последние районы действительно пострадали от постепенного истощения мегафауны, особенно более медленных видов (класс уязвимой мегафауны, олицетворяемый гигантские черепахи ) за последние несколько миллионов лет.[32][33]
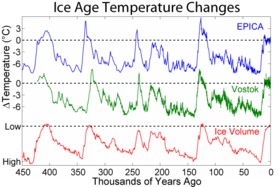
За пределами материковой части Афро-Евразия эти вымирания мегафауны происходили по очень характерной схеме для каждого отдельного участка суши, которая близко соответствует распространению людей в ранее необитаемые регионы мира и не показывает общей корреляции с историей климата (которую можно визуализировать с помощью графиков за последние геологические периоды климатических маркеров, таких как морские изотопы кислорода или уровни двуокиси углерода в атмосфере ).[34][35] Австралия[36] и близлежащие острова (например, Флорес[37]) были впервые обнаружены около 46000 лет назад, а затем Тасмания около 41000 лет назад (после образования сухопутного моста в Австралию около 43000 лет назад),[38][39][40] Япония очевидно, около 30 000 лет назад,[41] Северная Америка 13000 лет назад[заметка 2] Южная Америка примерно 500 лет спустя,[43][44] Кипр 10000 лет назад[45][46] то Антильские острова 6000 лет назад[47][48] Новая Каледония[49] и близлежащие острова[50] 3000 лет назад Мадагаскар 2000 лет назад[51] Новая Зеландия 700 лет назад[52] то Маскарены 400 лет назад[53] и Командорские острова 250 лет назад.[54] Почти все изолированные острова мира могут служить подобными примерами вымирания, происходящего вскоре после прибытия люди, хотя большинство этих островов, таких как Гавайские острова, никогда не было наземной мегафауны, поэтому их вымершая фауна были меньше.[34][35]
Анализ сроков Голарктический вымирания мегафауны и исчезновения за последние 56000 лет выявили тенденцию к группированию таких событий внутри межстадиальные, периоды резкого потепления, но только когда присутствовали люди. Люди могли препятствовать процессам миграции и повторного заселения, которые в противном случае позволили бы мегафаунам адаптироваться к изменению климата.[55] По крайней мере, в некоторых областях интерстадиалы были периодами увеличения человеческого населения.[56]
Анализ Спорормиелла грибковые споры (которые происходят в основном из навоза крупных травоядных) в кернах болотных отложений за последние 130000 лет от Кратер Линча в Квинсленд, Австралия, показали, что мегафауна этого региона практически исчезла около 41000 лет назад, в то время, когда изменения климата были минимальными; изменение сопровождалось увеличением количества древесного угля, за которым последовал переход от тропических лесов к огнестойким склерофилл растительность. Хронология изменений с высоким разрешением поддерживает гипотезу о том, что человеческая охота устранила мегафауну, и что последующее изменение флоры, скорее всего, было следствием отказа от браузеров и увеличения количества пожаров.[57][58][59][60] Увеличение количества пожаров отставало от исчезновения мегафауны примерно на столетие и, скорее всего, было результатом накопления топлива после прекращения просмотра. В течение следующих нескольких столетий трава увеличилась; растительность склерофиллов увеличилась с отставанием еще в одно столетие, и примерно через тысячу лет образовался склерофилловый лес.[59] В течение двух периодов изменения климата, примерно 120 000 и 75 000 лет назад, растительность склерофиллов также увеличилась на этом участке в ответ на переход к более прохладным и сухим условиям; ни один из этих эпизодов не оказал значительного влияния на численность мегафауны.[59] Аналогичные выводы относительно виновности людей-охотников в исчезновении плейстоценовой мегафауны были сделаны на основе хронологий высокого разрешения, полученных путем анализа большой коллекции фрагментов яичной скорлупы нелетающих австралийских птиц. Genyornis newtoni,[61][62][60] из анализа Спорормиелла грибковые споры из озера на востоке Северной Америки[63][64] и от изучения месторождений Шаста наземный ленивец навоз остался в более чем полдюжине пещер на юго-западе Америки.[65][66]
Продолжающаяся охота на людей и нарушение окружающей среды привели к дополнительным вымирания мегафауны в недавнем прошлом, и создал серьезная опасность дальнейшего исчезновения в ближайшем будущем (см. примеры ниже). Прямое убийство людей, в первую очередь ради мяса, является наиболее значительным фактором современного упадка мегафауны.[67][68]
Ряд других массовые вымирания произошел раньше в геологической истории Земли, когда вымерла часть или вся мегафауна того времени. Как известно, в Меловое – палеогеновое вымирание нептичьи динозавры и большинство других гигантских рептилий были уничтожены. Однако более ранние массовые вымирания были более глобальными и не столь избирательными для мегафауны; то есть многие виды других типов, включая растения, морские беспозвоночные[69] и планктон также вымерли. Таким образом, более ранние события должны были быть вызваны более общими типами нарушений биосфера.
Последствия истощения мегафауны
Влияние на транспорт питательных веществ
Мегафауна играет важную роль в боковом переносе питательных минеральных веществ в экосистеме, стремясь перемещать их из областей с высокой численностью в районы с меньшей численностью. Они делают это за счет своего движения между моментом, когда они потребляют питательное вещество, и временем, когда они высвобождают его путем выведения (или, в гораздо меньшей степени, путем разложения после смерти).[70] В Южной Америке Бассейн Амазонки, по оценкам, такая боковая диффузия сократилась более чем на 98% после мегафаунистических вымираний, произошедших примерно 12 500 лет назад.[71][72] При условии фосфор считается, что доступность ограничивает продуктивность в большей части региона, снижение его переноса из западной части бассейна и из пойм (оба из которых получают свои поставки в результате подъема Анды ) в другие районы, как полагают, значительно повлияли на экологию региона, и последствия, возможно, еще не достигли своих пределов.[72] Считается, что в море китообразные и ластоногие, питающиеся на глубине, переносят азот с глубины на мелководье, увеличивая тем самым продуктивность океана, и противодействуя активности зоопланктон, которые, как правило, делают наоборот.[73]
Влияние на выбросы метана
Большие популяции крупных травоядных могут внести значительный вклад в концентрацию в атмосфере метан, что является важным парниковый газ. Современный жвачный травоядные животные производить метан как побочный продукт ферментация передней кишки в пищеварении и высвободить его через отрыжку или метеоризм. Сегодня около 20% годовых выбросы метана происходят из выброса метана в животноводстве. в Мезозойский, было подсчитано, что зауроподы мог бы ежегодно выбрасывать в атмосферу 520 миллионов тонн метана,[74] способствуя более теплому климату того времени (на 10 ° C теплее, чем сейчас).[74][75] Этот большой выброс следует из огромной оценочной биомассы зауроподов, а также потому, что производство метана отдельными травоядными, как полагают, почти пропорционально их массе.[74]
Недавние исследования показали, что исчезновение мегафаунальных травоядных животных могло вызвать сокращение атмосферный метан. Эта гипотеза относительно нова.[76] В одном исследовании изучались выбросы метана из зубр что занимало Великие равнины Северной Америки до контакта с европейскими поселенцами. По оценкам исследования, вырубка зубров привела к сокращению численности на 2,2 миллиона тонн в год.[77] В другом исследовании изучалось изменение концентрации метана в атмосфере в конце периода Эпоха плейстоцена после исчезновения мегафауны в Америке. После того, как первые люди мигрировали в Америку, около 13000 человек BP, их охота и другие связанные с ними экологические воздействия привели к исчезновению там многих видов мегафауны. Расчеты показывают, что это вымирание уменьшило производство метана примерно на 9,6 млн тонн в год. Это предполагает, что отсутствие выбросов метана в мегафаунах могло способствовать резкому похолоданию климата в начале Младший дриас.[76] Произошедшее в то время уменьшение атмосферного метана, зафиксированное в ледяные керны, было в 2-4 раза быстрее, чем любое другое уменьшение за последние полмиллиона лет, что свидетельствует о работе необычного механизма.[76]
Примеры
Ниже приведены некоторые примечательные примеры животных, которые часто считаются мегафауной (в смысле определения «крупных животных»). Этот список не является исчерпывающим:
- Clade Synapsida
- Класс Млекопитающие (филогенетически, а клады внутри Терапсиды; см. ниже)
- Инфракласс Метатерия
- порядок Дипротодонтия
- В красный кенгуру (Macropus rufus) самый большой из живущих Австралийский млекопитающее и сумчатое животное при весе до 85 кг (187 фунтов). Однако его вымерший родственник гигантский короткомордый кенгуру Прокоптодон голия достиг 230 кг (510 фунтов),[78] в то время как вымершие дипротодонты достигла самого большого размера из всех сумчатых в истории, примерно до 2750 кг (6060 фунтов). Вымерший сумчатый лев (Thylacoleo carnifex ), весом до 160 кг (350 фунтов) был намного больше любого современного хищного сумчатого животного.
- порядок Дипротодонтия
- Инфракласс Евтерия
- Суперзаказ Афротерия
- порядок Хоботок
- Слонов являются самыми крупными из ныне живущих наземных животных. Они и их родственники возникли в Африка, но до недавнего времени имел почти мировое распространение. В Африканский слон (Loxodonta africana) имеет высоту плеча до 4,3 м (14 футов) и весит до 10,4 тонны (11,5 коротких тонн).[79] Среди недавно вымерших хоботков, мамонты (Маммутус) были близкими родственниками слонов, а мастодонты (Маммут) были гораздо более дальними родственниками. В степной мамонт (M. trogontherii) обычно весил около 10 тонн, что делает его, возможно, самым большим хоботок, что сделало бы его вторым по величине наземным млекопитающим после индрикотерины.
- порядок Сирения
- Самая большая сирена весом до 1500 кг (3300 фунтов) - это Вест-индийский ламантин (Trichechus manatus). Стеллерова морская корова (Hydrodamalis gigas), вероятно, был примерно в пять раз массивнее, но был истреблен людьми в течение 27 лет после его открытия за пределами Командорские острова в 1741 году. В доисторические времена эта морская корова также обитала на побережьях северо-востока Азия и северо-запад Северная Америка; очевидно, он был удален из этих более доступных мест охотниками-аборигенами.
- порядок Хоботок
- Суперзаказ Ксенартра
- порядок Cingulata
- В глиптодонты были группой больших, тяжело бронированных анкилозавр -любить ксенартранс связанные с жизнью броненосцы. Они возникли в Южная Америка, вторглись в Северную Америку во время Великий американский обмен, и вымерли в конце Плейстоцен эпоха.[80]
- порядок Pilosa
- Наземные ленивцы были еще одной группой медленных земных ксенартранов, родственных современным ленивцы. У них была похожая история, хотя они достигли Северной Америки раньше и распространились дальше на север (например, Мегалоникс ). Самые крупные роды, Мегатерий и Эремотерий достигли размеров, сопоставимых со слонами.[80]
- порядок Cingulata
- Суперзаказ Euarchontoglires
- порядок Приматы
- Самый большой из ныне живущих приматов весом до 266 кг (586 фунтов) - это горилла (Горилла берингей и Горилла горилла, причем три из четырех подвидов находящихся под угрозой исчезновения ). Вымерший малагасийский ленивый лемур Археоиндрис достиг такого же размера, в то время как вымершие Гигантопитек черный из Юго-Восточная Азия считается, что он был еще больше, хотя, вероятно, меньше, чем в два раза, вопреки ранним оценкам (отсутствие посткраниальных останков затрудняет оценку его размера).[81] Некоторые популяции архаичный Гомо были значительно больше в среднем, чем последние Homo sapiens;[82][83] Например, Homo heidelbergensis в южной части Африки обычно достигал 7 футов (2,1 м) в высоту,[84] в то время как Неандертальцы были примерно на 30% массивнее.[85]
- порядок Rodentia
- Сохранившиеся водосвинка (Hydrochoerus hydrochaeris) Южной Америки, самый крупный из ныне живущих грызунов, весит до 80 кг (180 фунтов).[86] Несколько недавно вымерших североамериканских форм были крупнее: капибара Neochoerus pinckneyi (еще один Неотропный мигрант) был в среднем примерно на 40% тяжелее; то гигантский бобр (Castoroides ohioensis) было похоже. Вымершие тупозубый гигант хутия (Amblyrhiza inundata) нескольких Карибский бассейн острова могли быть еще больше. Однако несколько миллионов лет назад в Южной Америке жили гораздо более массивные грызуны. Phoberomys pattersoni, известный из почти полного скелета, вероятно, достиг 700 кг (1500 фунтов). Фрагментарные остатки предполагают, что Josephoartigasia monesi выросла до более чем 1000 кг (2200 фунтов).
- порядок Приматы
- Суперзаказ Лавразиатерия
- порядок Хищник
- Самые крупные из существующих кошек относятся к роду Пантера, в том числе тигр (P. tigris) и лев (P. leo).[87] В Сибирский тигр (П. т. алтайский) должна быть самой большой дикой кошкой по мнению Правило Бергмана, и считался таковым некоторыми[88][89] но это спорный вопрос.[90] Исторически сложилось так, что дикие сибирские тигры уменьшились в размерах, и теперь они меньше, чем бенгальские тигры (П. т. тигр);[91] тем не менее, амурские тигры по-прежнему остаются самыми крупными тиграми в неволе, достигая веса около 320 кг (710 фунтов).[92] Пантера виды отличаются морфологический особенности, которые позволяют им рычать. Более крупные вымершие кошки включают Американский лев (П. атрокс) и южноамериканский саблезубый кот (Популятор смилодона ).
- Медведи крупные хищники пловидный подотряд. Самые крупные живые формы - это Полярный медведь (Ursus maritimus), с массой тела до 800 кг (1800 фунтов),[93] и почти такой же большой Кадьяк медведь (Ursus arctos middendorffi),[94] в соответствии с правилом Бергмана. Арктотерий августейцы, вымерший короткомордый медведь из Южной Америки, был самым крупным хищным наземным млекопитающим, когда-либо имеющим оценочный средний вес 1600 кг (3500 фунтов).[95]
- Тюлени, морские львы и моржи морские хищники-амфибии, которые произошли от медвежьих предков. В южный морской слон (Мироунга леонина) из Антарктика и субантарктический Waters является крупнейшим хищником всех времен: самцы-быки достигают максимальной длины 6–7 м (20–23 футов) и максимального веса 5 000 кг (11 000 фунтов).
- порядок Периссодактиля
- Тапиры - пасущиеся животные с короткой цепкой мордой и похожей на свинью формой, которая, кажется, мало изменилась за 20 миллионов лет. Они населяют тропические леса Юго-Восточной Азии и Южной и Центральной Америки, и включают в себя крупнейших выживших наземных животных двух последних регионов. Всего четыре вида.
- Носороги находятся однопалые копытные с рогами из кератин, того же типа белок составление волос. Они относятся к вторым по величине наземным млекопитающим с массой 850-3 800 кг. Три из пяти существующих видов являются находящихся под угрозой исчезновения. Их вымершие Центральная Азия родственники индрикотерины были самыми крупными наземными млекопитающими всех времен.
 Носорог, из Ксилография Дюрера
Носорог, из Ксилография Дюрера
- порядок Парнокопытные
- Жирафы (Giraffa spp.) - самые высокие из ныне живущих наземных животных, достигающие в высоту почти 6 м (20 футов). Средний вес составляет 1192 кг (2628 фунтов) для взрослого мужчины и 828 кг (1825 фунтов) для взрослой женщины с максимальным весом 1930 кг (4250 фунтов) и 1180 кг (2600 фунтов), зарегистрированным для мужчин и женщин, соответственно.
- Коровы копытные включают крупнейших сохранившихся наземных животных Европа и Северная Америка. В буйвол (Bubalis arnee), зубр (Бизон бизон и B. bonasus ), и гаур (Bos gaurus) могут вырасти до веса более 1000 кг (2200 фунтов).
- В полуводный бегемот (Бегемот амфибий) - самый тяжелый живой член отряда Cetartiodactyla после китообразных. Средний вес взрослого человека составляет около 1500 кг (3300 фунтов) и 1300 кг (2900 фунтов) для мужчин и женщин соответственно, при этом крупные самцы достигают более 3200 кг (7100 фунтов). Бегемот и гораздо меньший находящихся под угрозой исчезновения карликовый бегемот (Choeropsis liberiensis) считаются ближайшие сохранившиеся родственники китообразных. Бегемоты относятся к числу видов мегафауны. самый опасный людям.[96]
- Инфраордер Китообразные
- Киты, дельфины и морские свиньи морские млекопитающие. В синий кит (Balaenoptera musculus) самый большой усатый кит и самое большое животное, которое когда-либо существовало, на высоте 30 метров (98 футов)[97] в длину и 170 тонн (190 коротких тонн)[98] или больше по весу. В кашалот (Physeter macrocephalus) самый большой зубастый кит и один из крупнейших хищников в истории позвоночных, а также самый громкий и самое умное животное (с мозг примерно в пять раз массивнее, чем человеческий ). В косатка (Orcinus orca) - самый крупный дельфин.
- порядок Хищник
- Суперзаказ Афротерия
- Инфракласс Метатерия
- порядок Пеликозаврия (традиционный; парафилетический )
- Котилоринх был большим, когтистым, травоядным Идентификатор дела раннего Пермский период Северная Америка, достигая 6 м (20 футов) и 2 тонны.
- порядок Терапсида
- Антеозавр был бить головой, полуводный, плотоядный диноцефал из Средняя пермь Южная Африка. Он достигал 5–6 м (16–20 футов) в длину и весил около 500–600 кг (1100–1300 фунтов).[99]
- Лисовичия был размером со слона (9 тонн) травоядным каннемейериформ дицинодонт позднетриасовой Европы.[100][101]
- Класс Млекопитающие (филогенетически, а клады внутри Терапсиды; см. ниже)
- Clade Sauropsida
- Класс Авес (филогенетически клады внутри Целурозаврия, а таксон в отряде заурисхии; см. ниже)
- порядок Struthioniformes
- В ратиты древняя и разнообразная группа нелетающие птицы которые обнаружены на фрагментах бывшего суперконтинент Гондвана. Самая крупная из ныне живущих птиц, страус (Struthio camelus) был вытеснен вымершими Воромбе из Мадагаскар, самая тяжелая из группы - до (860 кг (1900 фунтов)), а вымершие гигантский моа (Dinornis) из Новая Зеландия, самый высокий, вырастающий до высоты 3,4 м (11 футов). Последние два являются примерами островной гигантизм.
- порядок Gastornithiformes
- Вымершие дроморнитиды Австралии, например Дроморнис приблизились к самым крупным по размеру ратитам. (Из-за небольшого размера континента и изолированности Австралия иногда рассматривается как самый большой остров в мире; таким образом, эти виды можно также считать островными гигантами.)
- порядок Cathartiformes
- Вымершие кондор -любить тераторн Аргентавис Южной Америки имел предполагаемый размах крыльев от 5 до 6 м (от 16 до 20 футов) и массу примерно 70 кг (150 фунтов), что делало его лучшим примером летающей птицы мегафауны.
- порядок Struthioniformes
- Класс Рептилии (традиционный; парафилетический )
- порядок Заурисхия
- Саурисчян динозавры из Юрский период и Меловой включают зауроподы, самые длинные (до 40 м или 130 футов) и самые массивные из известных наземных животных (Аргентинозавр достиг 80–100метрические тонны, или 90–110тонны ), а также тероподы, крупнейшие наземные хищники (Спинозавр вырос до 7–9 тонн; более известный Тиранозавр, до 6,8 т).
- порядок Птерозаврия
- Самый большой аждархид птерозавры, такие как Hatzegopteryx и Quetzalcoatlus, достигали размаха крыльев около 11–12 м (36–39 футов) и веса, вероятно, в диапазоне 70–250 кг (150–550 фунтов). Считается, что первый был главным хищником своего островная экосистема.
- порядок Крокодилы
- Аллигаторы и крокодилы крупные полуводные рептилии, самые крупные из которых, морской крокодил (Crocodylus porosus), может вырасти до веса 1360 кг (3000 фунтов). Далекие предки крокодилов и их родственники, псевдозухи (традиционный круротарзаны ), доминировал в мире в конце Триасовый, до Триас-юрское вымирание позволили динозаврам настигнуть их. Они оставались разнообразными и в более поздний период. Мезозойский, когда крокодиловидные такие как Дейнозух и Саркозух достигли длины 12 м. Так же крупные крокодилы, такие как Mourasuchus и Пуруссавр, присутствовали совсем недавно, когда Миоцен В Южной Америке.
- порядок Squamata
- В то время как самые крупные из сохранившихся ящерица, то Дракон Комодо (Varanus komodoensis), еще один островной гигант, может достигать 3 м (10 футов) в длину, его вымерший австралийский родственник Мегалания могли достигать более чем вдвое большего размера. Эти варан 'морские родственники, мозазавры, мы высшие хищники в позднемеловых морях.
- Самый тяжелый из сохранившихся змея считается зеленая анаконда (Eunectes murinus), в то время сетчатый питон (Питон ретикулатус) длиной до 8,7 м и более считается самой длинной. Вымерший австралиец Плиоцен виды Лиаз, то Гигантский питон Блафф Даунс, достигала 10 м, а Палеоцен Титанобоа Южной Америки достигала длины 12–15 м и предполагаемого веса около 1135 кг (2500 фунтов).
- порядок Testudines
- Самая большая черепаха - это находящихся под угрозой исчезновения морской кожистая черепаха (Dermochelys coriacea), весом до 900 кг (2000 фунтов). Он отличается от других морские черепахи из-за отсутствия костлявый оболочка. Самые массовые земные челониане - это гигантские черепахи из Галапагосские острова (Chelonoidis nigra ) и Атолл Альдабра (Альдабрахелис гигантский ) до 300 кг (660 фунтов). Эти черепахи - самые большие выжившие из множества видов гигантских черепах, которые широко распространены на континентальных территориях.[29][30] и дополнительные острова[29] в плейстоцене.
- порядок Заурисхия
- Класс Авес (филогенетически клады внутри Целурозаврия, а таксон в отряде заурисхии; см. ниже)
- Класс Амфибия (в широком, вероятно, парафилетическом смысле)
- порядок Темноспондили (отношение к сохранившиеся амфибии неясно)
- Пермский темноспондиль Прионозух, самая крупная из известных земноводных, достигала 9 м в длину и была водным хищником, напоминающим крокодилов. После появления настоящих крокодилов темноспондилы, такие как Koolasuchus (5 м в длину) отступили в Антарктический регион к меловому периоду, прежде чем вымерли.
- порядок Темноспондили (отношение к сохранившиеся амфибии неясно)
- Класс Актиноптеригии
- порядок Tetraodontiformes
- Самый крупный из сохранившихся костлявая рыба это океанская солнечная рыба (Мола Мола), средний вес взрослого человека составляет 1000 кг (2200 фунтов). Хотя филогенетически это «костлявая рыба», ее скелет в первую очередь хрящ (что легче, чем кость ). Он имеет дискообразное тело и движется вперед с помощью длинного, тонкого спинной и анальные плавники; он питается в первую очередь медуза. В этих трех отношениях (а также по размеру и привычкам нырять) он напоминает кожистую черепаху.
- порядок Lampriformes
- В гигантская рыба-весло (Regalecus glesne) - самая длинная костистая рыба, достигающая 11 м (36 футов).
- порядок Acipenseriformes
- В находящихся под угрозой исчезновения белуга (Европейский осетр, Huso Huso) весом до 1476 кг (3254 фунта) является самым большим осетр (которые также в основном хрящевые) и считаются крупнейшими проходной рыбы.
- порядок Силурообразные
- В находящихся под угрозой исчезновения Меконгский гигантский сом (Pangasianodon gigas) весом до 293 кг (646 фунтов) часто считается самым большим пресноводная рыба.
- порядок Tetraodontiformes
- Класс Chondrichthyes
- порядок Lamniformes
- Самая крупная из ныне живущих хищных рыб - Большая белая акула (Carcharodon carcharias), достигает веса до 2240 кг (4940 фунтов). Его вымерший родственник C. мегалодон (спорным родом является либо Кархародон или Кархарокл) был больше, чем порядок величины больше по размеру и является самой крупной хищной акулой или рыбой всех времен (и одним из крупнейших хищников в истории позвоночных); он охотился на китов и других морские млекопитающие.
- порядок Orectolobiformes
- Самый крупный из сохранившихся акула, хрящевые рыбы, и рыбы в целом это Китовая акула (Ринкодон тип), который достигает веса более 21,5 тонны (47 000 фунтов). Подобно усатым китам, это питатель фильтра и в первую очередь потребляет планктон.
- порядок Rajiformes
- В скат манта (Манта бирострис) - еще один фильтр-питатель и самый большой луч, дорастает до 2300 кг.
- порядок Lamniformes
- Класс Плакодерми
- порядок Артродира
- Самая большая бронированная рыба, Дунклеостей, возник в конце девона. Длиной до 10 метров (33 футов)[102] и 3,6 тонны (4.0 короткие тонны ) в массе,[103] это было гиперплотоядный хищник вершины которые наняли всасывающее питание.[104][105] Его современник, Titanichthys, очевидно, ранний фильтр-питатель, соперничал с ним по размеру. Артродиры были ликвидированы в результате экологических потрясений Позднее девонское вымирание, просуществовав всего около 50 миллионов лет.
- порядок Артродира
- Класс Головоногие моллюски
- порядок Аммонитиды
- Поздний мел аммонит Парапузозия сеппенраденсис достигал диаметра снаряда более 2 м.
- порядок Teuthida
- Ряд глубоководных существ выставляют бездонный гигантизм. К ним относятся Гигантский кальмар (Architeuthis) и колоссальный кальмар (Mesonychoteuthis hamiltoni); Считается, что оба (хотя их редко можно увидеть) достигают длины 12 м (39 футов) или более. Последний является крупнейшим в мире беспозвоночный, и имеет самый большой глаза любого животного. Оба являются жертвами кашалотов.
- порядок Аммонитиды
- Стебель-группа Членистоногие
- порядок Радиодонта
- Аномалокариды были группой очень ранних безногих морских членистоногих, в которую входили крупнейшие хищники Кембрийский, такие как Аномалокарис. К раннему Ордовик они превратились в гигантские (на то время) фильтраторы, очевидно, в ответ на распространение планктона во время Великое событие биоразнообразия ордовика. Aegirocassis вырос до более 2 м в длину.[106]
- порядок Eurypterida
- Эвриптериды (морские скорпионы) были разнообразной группой водных и, возможно, амфибийных хищников, в которую входили самые массивные хищники. членистоногие чтобы существовать. Они прожили более 200 миллионов лет, но окончательно вымерли в Пермско-триасовое вымирание вместе с трилобиты и большинство других форм жизни, присутствовавших в то время, включая большинство доминирующих наземных терапсидов. Рано Девонский Jaekelopterus достигла расчетной длины 2,5 м (8,2 фута), не считая его хищный хелицеры, и считается, что это был пресноводный вид.
- порядок Радиодонта

Галерея
Вымершие

Немного Палеозой морские скорпионы (Евриптер показаны) были крупнее человека.

Дунклеостей был беззубым, длиной 10 м (33 фута) бронированный хищный девонский плакодерма рыбы.

С парусной опорой пеликозавр Диметродон и темноспондил Эриопс из Северной Америки Пермский период.

Leedsichthys, серединаЮрский период питатель фильтра рыба, возможно, достигала размеров 7–16,5 м (23–54 футов).

Макронарский зауроподы; слева, Камаразавр, Брахиозавр, Жирафатитан, Euhelopus.

Тиранозавр был 12,3 м (40 футов) в длину теропод динозавр, хищник вершины из запад Северной Америки.

Азиатский Indricothere носорог Парацератерий был одним из крупнейших наземных млекопитающих,[107] примерно вдвое больше массы куста слона.

В Поздний миоцен тераторн Аргентавис Южной Америки имел размах крыльев 7 м (23 фута).

Реконструированные челюсти C. мегалодон (Балтимор ).

Деинотерий имели загнутые вниз клыки и широко располагались на Афро-Евразия.

Титанис Валлери, единственный ужасная птица известно иметь вторгся Северная Америка, была 2,5 м (8,2 фута) в высоту.

Размером с бегемота Дипротодон Австралии, крупнейший сумчатое животное всех времен вымерли 40 000 лет назад.

Глиптодон, из Южной Америки Плейстоцен, был размер автоматически поясная извилина, родственник броненосцы.

Американские львы превышен сохранившийся львы по размеру и простиралась на большей части Северной Америки до 11000 BP.

Шерстистые мамонты исчез после того, как люди вторглись в их среду обитания в Евразии и Северной Америке.[1]

Орел Хааста, самый большой известный орел, атакующий моа (который включал самая высокая птица известный).


















Живущий

В восточная горилла самый большой и один из самых находящиеся под угрозой исчезновения приматы на планете.

Самый распространенный подвид тигра, Бенгальские тигры находятся находящихся под угрозой исчезновения от браконьерство и разрушение среды обитания.

Полярные медведи, среди крупнейших медведи (в соответствии с Правило Бергмана ), находятся уязвимый к глобальное потепление.

В находящихся под угрозой исчезновения черный носорог длиной до 3,75 м (12,3 фута) под угрозой браконьерством.

Дикие бактрийские верблюды находятся под угрозой исчезновения. Их предки возник в Северной Америке.

В отличие от шерстяных носороги и мамонты, овцебык едва пережил Четвертичное вымирание.[1]

Бегемоты, самый тяжелый и самый водный копытные, находятся киты ' ближайшие живые родственники.

В кашалот, самый большой зубастый кит и зубчатый хищник, имеет самый большой мозг.

В Orca, самый большой дельфин и стая хищников, является очень умный и живет в сложные общества.

В казуар, самая тяжелая неафриканская птица, может бегать со скоростью 50 км / ч через плотную тропический лес.

В морской крокодил самый большой из живущих рептилия и опасный хищник людей.

В Дракон Комодо, островной гигант и самый большой ящерица, имеет зубчатый зубы и ядовитый укус.

В зеленая анаконда, водный констриктор, самый тяжелый змея, весом до 97,5 кг (215 фунтов) или более.

В глубокое погружение океанская солнечная рыба самый большой костлявая рыба, но его скелет в основном хрящевой.

В Нильский окунь, одна из крупнейших пресноводных рыб, также является опасным инвазивные виды.[заметка 3]

В большой белый, самая большая макрохищная рыба, находится под большей угрозой, чем тигр.[110]

Обследование 9 м Гигантский кальмар, бездонный гигант и второй по величине головоногие моллюски.


















{kind=link}
{kind=link}
Смотрите также
- Австралийская мегафауна
- Правило Бергмана
- Харизматичная мегафауна
- Правило Копа
- Глубоководный гигантизм
- Фауна
- Островная карликовость
- Островной гигантизм
- Крупнейшие организмы
- Крупнейшие доисторические организмы
- Список самых тяжелых наземных млекопитающих
- Список крупнейших млекопитающих
- Список мегафауны, обнаруженной в наше время
- Мегафауна (мифология)
- Мегафауна волк
- Мегафлора
- Megaherb
- Плейстоценовые вымирания в Новом Свете
- Мегафауна плейстоцена
- Событие четвертичного вымирания
Заметки
- ^ Размер неавианских динозавров не ограничивался аналогичным образом, потому что у них было другое соотношение между массой тела и размером яйца, чем у птиц. 400 кг Эпиорнис яйца были больше, чем почти у всех динозавров.[26][27]
- ^ Анализ показывает, что в этом случае более или менее одновременно вымерли 35 родов североамериканских млекопитающих.[42]
- ^ Перспектива заставляет рыбу казаться больше по сравнению с человеком, стоящим за ней (еще один пример мегафауны), чем она есть на самом деле.
использованная литература
- ^ а б c d Стюарт, А. Дж. (Ноябрь 1991 г.). «Вымирание млекопитающих в позднем плейстоцене Северной Евразии и Северной Америки». Биологические обзоры. 66 (4): 453–562. Дои:10.1111 / j.1469-185X.1991.tb01149.x. PMID 1801948. S2CID 41295526.
- ^ Мартин, П. С. (1984). «Доисторическое излишество: глобальная модель». В Martin, P. S .; Кляйн, Р. Г. (ред.). Четвертичное вымирание: доисторическая революция. Университет Аризоны Press. С. 354–403. ISBN 978-0-8165-1100-6. OCLC 258362030.
- ^ Мартин, П.С.; Стедман, Д. В. (1999-06-30). «Доисторические вымирания на островах и континентах». В MacPhee, R. D. E (ed.). Вымирание в ближайшее время: причины, контексты и последствия. Успехи палеобиологии позвоночных. 2. Нью-Йорк: Kluwer / Plenum. С. 17–56. ISBN 978-0-306-46092-0. OCLC 41368299. Получено 2011-08-23. см. страницу 17
- ^ Ричард А. Фарина, Серджио Ф. Бискайно, Джерри Де Юлиис (2013). "Великий американский биотический обмен". Мегафауна: гигантские звери плейстоцена Южной Америки. Издательство Индианского университета, Блумингтон, Индиана. п. 150. ISBN 978-0-253-00230-3.CS1 maint: использует параметр авторов (ссылка на сайт)
- ^ Бернхард А. Хубер, Брэдли Дж. Синклер, Карл-Хайнц Лампе (2005). «Исторические детерминанты видов млекопитающих в Африке». Африканское биоразнообразие: молекулы, организмы, экосистемы. Springer. п. 294. ISBN 978-0387243153.CS1 maint: использует параметр авторов (ссылка на сайт)
- ^ Животные ледникового периода. Государственный музей Иллинойса
- ^ https://www.britannica.com/science/K-selected-species. Британника. Проверено 2 апреля 2017.
- ^ Барноски, А. Д. (2004-10-01). «Оценка причин позднего плейстоценового вымирания на континентах». Наука. 306 (5693): 70–75. Bibcode:2004Наука ... 306 ... 70Б. CiteSeerX 10.1.1.574.332. Дои:10.1126 / science.1101476. PMID 15459379. S2CID 36156087.
- ^ Brook, B.W .; Джонсон, К. Н. (2006). «Селективная охота на молодь как причина незаметного массового истребления австралийской мегафауны плейстоцена». Алчеринга: Австралазийский журнал палеонтологии. 30 (sup1): 39–48. Дои:10.1080/03115510609506854. S2CID 84205755.
- ^ а б c d е ж г Evans, A.R .; Jones, D .; Boyer, A. G .; Brown, J. H .; Коста, Д. П .; Эрнест, С. К. М .; Фитцджеральд, Э. М. Г .; Фортелиус, М .; Gittleman, J. L .; Гамильтон, М. Дж .; Harding, L.E .; Lintulaakso, K .; Lyons, S.K .; Okie, J.G .; Saarinen, J. J .; Сибли, Р. М .; Smith, F.A .; Stephens, P. R .; Theodor, J.M .; Ухен, М. Д. (30 января 2012 г.). «Максимальная скорость эволюции млекопитающих». PNAS. 109 (11): 4187–4190. Bibcode:2012PNAS..109.4187E. Дои:10.1073 / pnas.1120774109. ЧВК 3306709. PMID 22308461.
- ^ а б c d е ж г Smith, F.A .; Boyer, A. G .; Brown, J. H .; Costa, D. P .; Даян, Т .; Эрнест, С. К. М .; Evans, A.R .; Fortelius, M .; Gittleman, J. L .; Гамильтон, М. Дж .; Harding, L.E .; Lintulaakso, K .; Lyons, S.K .; McCain, C .; Okie, J.G .; Saarinen, J. J .; Сибли, Р. М .; Stephens, P. R .; Theodor, J .; Ухен, М. Д. (26 ноября 2010 г.). «Эволюция максимального размера тела наземных млекопитающих». Наука. 330 (6008): 1216–1219. Bibcode:2010Научный ... 330.1216S. CiteSeerX 10.1.1.383.8581. Дои:10.1126 / science.1194830. PMID 21109666. S2CID 17272200.
- ^ Клаусс, М .; Frey, R .; Кифер, В .; Lechner-Doll, M .; Loehlein, W .; Polster, C .; Roessner, G.E .; Штрайх, В. Дж. (24 апреля 2003 г.). «Максимально достижимый размер тела травоядных млекопитающих: морфофизиологические ограничения на переднюю кишку и адаптация ферментеров задней кишки» (PDF). Oecologia. 136 (1): 14–27. Bibcode:2003 Oecol.136 ... 14C. Дои:10.1007 / s00442-003-1254-z. PMID 12712314. S2CID 206989975.
- ^ а б Соркин Б. (2008-04-10). «Биомеханическое ограничение массы тела у наземных хищников млекопитающих». Lethaia. 41 (4): 333–347. Дои:10.1111 / j.1502-3931.2007.00091.x.
- ^ Carbone, C .; Учитель, А; Роклифф, Дж. М. (16 января 2007 г.). "Цена хищничества". PLOS Биология. 5 (2, e22): 363–368. Дои:10.1371 / journal.pbio.0050022. ЧВК 1769424. PMID 17227145.
- ^ а б c Ashton, K. G .; Tracy, M.C .; де Кейруш, А. (октябрь 2000 г.). «Действительно ли правило Бергмана для млекопитающих?». Американский натуралист. 156 (4): 390–415. Дои:10.1086/303400. JSTOR 10.1086/303400. PMID 29592141.
- ^ Уэбб, Дж. (19 февраля 2015 г.). «Эволюция» отдает предпочтение более крупных морских существ'". Новости BBC. BBC. Получено 2015-02-22.
- ^ Heim, N.A .; Ноуп, М. Л .; Schaal, E.K .; Wang, S.C .; Пейн, Дж. Л. (20 февраля 2015 г.). «Правило Коупа в эволюции морских животных». Наука. 347 (6224): 867–870. Bibcode:2015Научный ... 347..867H. Дои:10.1126 / science.1260065. PMID 25700517.
- ^ Thewissen, J.G.M .; Баджпай, С.(1 января 2001 г.). «Происхождение китов как пример макроэволюции». Бионаука. 51 (12): 1037–1049. Дои:10.1641 / 0006-3568 (2001) 051 [1037: WOAAPC] 2.0.CO; 2. ISSN 0006-3568.
- ^ а б Goldbogen, J. A .; Кейд, Д. Э .; Вишневска, Д. М .; Potvin, J .; Segre, P. S .; Savoca, M. S .; Hazen, E.L .; Чапанский, М. Ф .; Kahane-Rapport, S. R .; DeRuiter, S.L .; Gero, S .; Tønnesen, P .; Gough, W. T .; Hanson, M. B .; Holt, M. M .; Jensen, F. H .; Саймон, М .; Stimpert, A.K .; Arranz, P .; Johnston, D.W .; Новачек, Д. П .; Парки, С. Э .; Visser, F .; Friedlaender, A. S .; Tyack, P.L .; Madsen, P.T .; Пиенсон, Н. Д. (2019). «Почему киты большие, но не больше: физиологические факторы и экологические ограничения в эпоху океанских гигантов». Наука. 366 (6471): 1367–1372. Bibcode:2019Научный ... 366.1367G. Дои:10.1126 / science.aax9044. HDL:10023/19285. PMID 31831666. S2CID 209339266.
- ^ а б c Mitchell, K.J .; Llamas, B .; Soubrier, J .; Rawlence, N.J .; Уорти, Т. Х .; Wood, J .; Lee, M. S. Y .; Купер, А. (23.05.2014). «Древняя ДНК показывает, что птицы-слоны и киви являются родственными таксонами, и проясняет эволюцию птиц-ратитов» (PDF). Наука. 344 (6186): 898–900. Bibcode:2014Наука ... 344..898М. Дои:10.1126 / science.1251981. HDL:2328/35953. PMID 24855267. S2CID 206555952.
- ^ Buffetaut, E .; Ангст, Д. (ноябрь 2014 г.). «Стратиграфическое распространение крупных нелетающих птиц в палеогене Европы и его палеобиологические и палеогеографические последствия». Обзоры наук о Земле. 138: 394–408. Bibcode:2014ESRv..138..394B. Дои:10.1016 / j.earscirev.2014.07.001.
- ^ Филлипс М.Дж., Гибб Г.К., Кримп Е.А., Пенни Д. (январь 2010 г.). «Тинамус и моа собираются вместе: анализ последовательности митохондриального генома показывает независимые потери полета среди крысолов». Систематическая биология. 59 (1): 90–107. Дои:10.1093 / sysbio / syp079. PMID 20525622.
- ^ Бейкер, А. Дж .; Haddrath, O .; McPherson, J.D .; Клотье, А. (2014). «Геномная поддержка клады Моа-Тинамо и адаптивной морфологической конвергенции у нелетающих крыс». Молекулярная биология и эволюция. 31 (7): 1686–1696. Дои:10.1093 / молбев / мсу153. PMID 24825849.
- ^ Мюррей, Питер Ф .; Викерс-Рич, Патрисия (2004). Великолепные Михирунг: Колоссальные нелетающие птицы австралийских времен снов. Издательство Индианского университета. С. 51, 314. ISBN 978-0-253-34282-9. Получено 7 января 2012.
- ^ Там же (2004). п. 212. ISBN 978-0253342829.
- ^ Кеннет Карпентер (1999). Яйца, гнезда и детеныши динозавров: взгляд на воспроизводство динозавров. Издательство Индианского университета. п.100. ISBN 978-0-253-33497-8. OCLC 42009424. Получено 6 мая 2013.
- ^ Джексон, Ф. Д .; Varricchio, D. J .; Джексон, Р. А .; Vila, B .; Чиаппе, Л. М. (2008). "Сравнение проводимости водяного пара в яйце титанозавра из верхнего мела Аргентины и Megaloolithus siruguei яйцо из Испании ». Палеобиология. 34 (2): 229–246. Дои:10.1666 / 0094-8373 (2008) 034 [0229: COWVCI] 2.0.CO; 2. ISSN 0094-8373.
- ^ Там же (2004). п. 277. ISBN 978-0253342829.
- ^ а б c Hansen, D. M .; Donlan, C.J .; Griffiths, C.J .; Кэмпбелл, К. Дж. (Апрель 2010 г.). «Экологическая история и скрытый потенциал сохранения: большие и гигантские черепахи как модель для замены таксонов» (PDF). Экография. 33 (2): 272–284. Дои:10.1111 / j.1600-0587.2010.06305.x. Архивировано из оригинал (PDF) 24 июля 2011 г.. Получено 2011-02-26.
- ^ а б Cione, A. L .; Tonni, E. P .; Сойбелзон, Л. (2003). "Сломанный зигзаг: вымирание крупных млекопитающих и черепах в позднем кайнозое в Южной Америке" (PDF). Rev. Mus. Argentino Cienc. Nat., N.s. 5 (1): 1–19. Дои:10.22179 / REVMACN.5.26. ISSN 1514-5158. Архивировано из оригинал (PDF) 6 июля 2011 г.. Получено 2011-02-06.
- ^ Алрой, Дж. (1999), «Рассмотрение вымирания мегафауны в конце плейстоцена в Северной Америке в контексте: крупномасштабный анализ пространственных закономерностей, темпов вымирания и распределения размеров», в Макфи, Р. Д. Э. (ред.), Вымирание в ближайшее время: причины, контексты и последствия, Достижения в палеобиологии позвоночных, 2, Нью-Йорк: Пленум, стр. 105–143, Дои:10.1007/978-1-4757-5202-1_6, ISBN 978-1-4757-5202-1, OCLC 41368299
- ^ Корлетт, Р. Т. (2006). «Вымирание мегафауны в тропической Азии» (PDF). Тропинет. 17 (3): 1–3. Получено 2010-10-04.
- ^ Эдмидс, Баз. «Мегафауна - первые жертвы антропогенного вымирания». megafauna.com. (опубликованная в Интернете книга с предисловием Пол С. Мартин ). Архивировано из оригинал на 2014-12-25. Получено 2020-02-13.
- ^ а б Мартин, П.С. (2005). "Глава 6. Смертельная синкопа". Сумерки мамонтов: вымирание ледникового периода и возрождение Америки. Калифорнийский университет Press. С. 118–128. ISBN 978-0-520-23141-2. OCLC 58055404. Получено 2014-11-11.
- ^ а б Burney, D.A .; Фланнери, Т.Ф. (Июль 2005 г.). «Пятьдесят тысячелетий катастрофических исчезновений после контакта с людьми» (PDF). Тенденции в экологии и эволюции. 20 (7): 395–401. Дои:10.1016 / j.tree.2005.04.022. PMID 16701402. Архивировано из оригинал (PDF) на 2010-06-10. Получено 2014-11-11.
- ^ Roberts, R.G .; Фланнери, Т.Ф.; Ayliffe, L.K .; Yoshida, H .; Olley, J.M .; Prideaux, G.J .; Laslett, G.M .; Baynes, A .; Smith, M. A .; Jones, R .; Смит, Б. Л. (2001-06-08). "Новые века последней австралийской мегафауны: исчезновение в масштабах всего континента около 46 000 лет назад" (PDF). Наука. 292 (5523): 1888–1892. Bibcode:2001Научный ... 292.1888R. Дои:10.1126 / science.1060264. PMID 11397939. S2CID 45643228. Получено 2011-08-26.
- ^ Каллавей, Э. (21 сентября 2016 г.). «Человеческие останки найдены в пещере хоббитов». Природа. Дои:10.1038 / природа.2016.20656. S2CID 89272546.
- ^ Даймонд, Джаред (13.08.2008). «Палеонтология: последний гигантский кенгуру». Природа. 454 (7206): 835–836. Bibcode:2008Натура.454..835D. Дои:10.1038 / 454835a. PMID 18704074. S2CID 36583693.
- ^ Turney, C. S. M .; Фланнери, Т.Ф.; Roberts, R.G .; Reid, C .; Fifield, L.K .; Higham, T. F. G .; Jacobs, Z .; Kemp, N .; Colhoun, E.A .; Калин, Р. М .; Огл, Н. (21 августа 2008 г.). «Мегафауна, сохранившаяся поздно, в Тасмании, Австралия, предполагает участие человека в их исчезновении». PNAS. 105 (34): 12150–12153. Bibcode:2008ПНАС..10512150Т. Дои:10.1073 / pnas.0801360105. ЧВК 2527880. PMID 18719103.
- ^ Roberts, R .; Джейкобс, З. (октябрь 2008 г.). "Затерянные гиганты Тасмании" (PDF). Австралазийская наука. 29 (9): 14–17. Архивировано из оригинал (PDF) на 2011-09-27. Получено 2011-08-26.
- ^ Norton, C.J .; Кондо, Й .; Оно, А .; Zhang, Y .; Диаб, М. К. (23 мая 2009 г.). «Природа вымирания мегафауны во время перехода MIS 3–2 в Японии». Четвертичный международный. 211 (1–2): 113–122. Bibcode:2010QuInt.211..113N. Дои:10.1016 / j.quaint.2009.05.002.
- ^ Faith, J. T .; Суровелл, Т. А. (2008-12-08). «Синхронное вымирание плейстоценовых млекопитающих Северной Америки». Труды Национальной академии наук. 106 (49): 20641–20645. Bibcode:2009PNAS..10620641F. Дои:10.1073 / pnas.0908153106. ЧВК 2791611. PMID 19934040.
- ^ Хейнс, Гэри (2009). «Введение в том». В Хейнсе, Гэри (ред.). Вымирание мегафауны в Америке в конце плейстоцена. Палеобиология и палеоантропология позвоночных. Springer. С. 1–20. Дои:10.1007/978-1-4020-8793-6_1. ISBN 978-1-4020-8792-9.
- ^ Фидель, Стюарт (2009). «Внезапные смерти: хронология вымирания мегафауны в конце плейстоцена». В Хейнсе, Гэри (ред.). Вымирание мегафауны в Америке в конце плейстоцена. Палеобиология и палеоантропология позвоночных. Springer. С. 21–37. Дои:10.1007/978-1-4020-8793-6_2. ISBN 978-1-4020-8792-9.
- ^ Симмонс, А. Х. (1999). Вымирание фауны в островном обществе: охотники на карликовых бегемотов на Кипре. Междисциплинарные вклады в археологию. Kluwer Academic / Plenum Publishers. п. 382. Дои:10.1007 / b109876. ISBN 978-0-306-46088-3. OCLC 41712246.
- ^ Simmons, A.H .; Мандель, Р. Д. (декабрь 2007 г.). «Не такой новый свет: ответ Аммерману и Ноллеру». Мировая археология. 39 (4): 475–482. Дои:10.1080/00438240701676169. JSTOR 40026143. S2CID 161791746.
- ^ Стедман, Д. В.; Мартин, П.С.; MacPhee, R.D.E .; Jull, A. J. T .; McDonald, H.G .; Woods, C.A .; Iturralde-Vinent, M .; Ходгинс, Г. В. Л. (16 августа 2005 г.). «Асинхронное вымирание позднечетвертичных ленивцев на континентах и островах». Proc. Natl. Акад. Sci. Соединенные Штаты Америки. 102 (33): 11763–11768. Bibcode:2005ПНАС..10211763С. Дои:10.1073 / pnas.0502777102. ЧВК 1187974. PMID 16085711.
- ^ Cooke, S. B .; Dávalos, L.M .; Мычайлив, А. М .; Turvey, S.T .; Упхэм, Н. С. (2017). «Антропогенное вымирание преобладает в голоцене западно-индийских млекопитающих». Ежегодный обзор экологии, эволюции и систематики. 48 (1): 301–327. Дои:10.1146 / annurev-ecolsys-110316-022754.
- ^ Андерсон, А .; Sand, C .; Petchey, F .; Уорти, Т. Х. (2010). «Исчезновение фауны и обитание людей в Новой Каледонии: первые результаты и последствия новых исследований в пещерах Пиндай». Журнал тихоокеанской археологии. 1 (1): 89–109. HDL:10289/5404.
- ^ White, A. W .; Уорти, Т. Х .; Hawkins, S .; Бедфорд, S .; Сприггс, М. (16 августа 2010 г.). «Рогатые черепахи-мейоланиды-мегафауна дожили до первых поселений людей в Вануату, юго-западная часть Тихого океана». Proc. Natl. Акад. Sci. Соединенные Штаты Америки. 107 (35): 15512–15516. Bibcode:2010PNAS..10715512W. Дои:10.1073 / pnas.1005780107. ЧВК 2932593. PMID 20713711.
- ^ Burney, D.A .; Burney, L.P .; Годфри, Л. Р .; Jungers, W. L .; Goodman, S.M .; Wright, H.T .; Джулл. А. Дж. Т. (июль 2004 г.). «Хронология позднего доисторического Мадагаскара». Журнал эволюции человека. 47 (1–2): 25–63. Дои:10.1016 / j.jhevol.2004.05.005. PMID 15288523.
- ^ Holdaway, R. N .; Джакомб, К. (2000-03-24). «Быстрое вымирание моа (Aves: Dinornithiformes): модель, испытание и последствия». Наука. 287 (5461): 2250–2254. Bibcode:2000Sci ... 287.2250H. Дои:10.1126 / science.287.5461.2250. PMID 10731144.
- ^ Яну, А. (апрель 2005 г.). «Открытие изолированных костей дронта (Raphus cucullatus (L.), Aves, Columbiformes) из пещерных убежищ Маврикия подчеркивает человеческое хищничество с комментарием о статусе семейства Raphidae Wetmore, 1930 ». Анналы палеонтологии. 91 (2): 167–180. Дои:10.1016 / j.annpal.2004.12.002.
- ^ Андерсон, П. К. (июль 1995 г.). «Конкуренция, хищничество, эволюция и исчезновение белопольной морской коровы», Hydrodamalis gigas". Наука о морских млекопитающих. 11 (3): 391–394. Дои:10.1111 / j.1748-7692.1995.tb00294.x. Архивировано из оригинал на 2011-05-11. Получено 2011-08-30.
- ^ Купер, А .; Turney, C .; Hughen, K. A .; Brook, B.W .; McDonald, H.G .; Брэдшоу, К. Дж. А. (23 июля 2015 г.). «Резкие потепления стимулировали круговорот мегафауны позднего плейстоцена в Голарктике». Наука. 349 (6248): 602–6. Bibcode:2015Научный ... 349..602C. Дои:10.1126 / science.aac4315. PMID 26250679. S2CID 31686497.
- ^ Müller, U.C .; Pross, J .; Tzedakis, P.C .; Gamble, C .; Kotthoff, U .; Schmiedl, G .; Wulf, S .; Кристанис, К. (февраль 2011 г.). «Роль климата в распространении современного человека в Европу». Четвертичные научные обзоры. 30 (3–4): 273–279. Bibcode:2011QSRv ... 30..273M. Дои:10.1016 / j.quascirev.2010.11.016.
- ^ Бьелло, Д. (22 марта 2012 г.). "Большое убийство, а не большой холод, покончено с гигантскими кенгуру". Новости журнала Scientific American. Получено 2012-03-25.
- ^ МакГлоун, М. (23 марта 2012 г.). «Это сделали охотники». Наука. 335 (6075): 1452–1453. Bibcode:2012Наука ... 335.1452M. Дои:10.1126 / science.1220176. PMID 22442471. S2CID 36914192.
- ^ а б c Правило, S .; Brook, B.W .; Haberle, S.G .; Turney, C. S. M .; Кершоу, А. П. (23 марта 2012 г.). «Последствия вымирания мегафауны: трансформация экосистемы в плейстоценовой Австралии». Наука. 335 (6075): 1483–1486. Bibcode:2012Научный ... 335.1483R. Дои:10.1126 / наука.1214261. PMID 22442481. S2CID 26675232.
- ^ а б Johnson, C.N .; Alroy, J .; Beeton, N.J .; Bird, M. I .; Brook, B.W .; Купер, А .; Gillespie, R .; Herrando-Pérez, S .; Jacobs, Z .; Miller, G.H .; Prideaux, G.J .; Roberts, R.G .; Rodríguez-Rey, M .; Saltré, F .; Turney, C. S. M .; Брэдшоу, К. Дж. А. (10 февраля 2016 г.). «Что стало причиной исчезновения плейстоценовой мегафауны Сахула?». Труды Королевского общества B: биологические науки. 283 (1824): 20152399. Дои:10.1098 / rspb.2015.2399. ЧВК 4760161. PMID 26865301.
- ^ Miller, G.H .; Magee, J. W .; Johnson, B.J .; Fogel, M. L .; Spooner, N.A .; McCulloch, M. T .; Эйлифф, Л. К. (1999-01-08). "Плейстоценовое вымирание Genyornis newtoni: Влияние человека на австралийскую мегафауну ». Наука. 283 (5399): 205–208. Дои:10.1126 / science.283.5399.205. PMID 9880249.
- ^ Miller, G .; Magee, J .; Smith, M .; Spooner, N .; Baynes, A .; Lehman, S .; Fogel, M .; Johnston, H .; Уильямс, Д .; Clark, P .; Флориан, С .; Holst, R .; ДеФогель, С. (29 января 2016 г.). «Человеческие хищники способствовали исчезновению австралийской мегафауны птицы Genyornis newtoni ∼47 тыс. Лет назад». Nature Communications. 7: 10496. Bibcode:2016НатКо ... 710496M. Дои:10.1038 / ncomms10496. ЧВК 4740177. PMID 26823193.
- ^ Джонсон, К. (20 ноября 2009 г.). «Упадок и падение мегафауны». Наука. 326 (5956): 1072–1073. Bibcode:2009Sci ... 326.1072J. Дои:10.1126 / science.1182770. PMID 19965418. S2CID 206523763.
- ^ Gill, J. L .; Уильямс, Дж. В .; Джексон, С. Т .; Lininger, K. B .; Робинсон, Г.С. (20 ноября 2009 г.). «Плейстоценовый мегафаунальный коллапс, новые сообщества растений и усиленные пожарные режимы в Северной Америке» (PDF). Наука. 326 (5956): 1100–1103. Bibcode:2009Научный ... 326.1100G. Дои:10.1126 / science.1179504. PMID 19965426. S2CID 206522597.
- ^ Федал, Стюарт (2009). «Внезапные смерти: хронология вымирания мегафауны в конце плейстоцена». В Хейнсе, Гэри (ред.). Вымирание мегафауны в Америке в конце плейстоцена. Палеобиология и палеоантропология позвоночных. Springer. С. 21–37. Дои:10.1007/978-1-4020-8793-6_2. ISBN 978-1-4020-8792-9.
- ^ Мартин, П.С. (2005). «Глава 4. Земляные ленивцы дома». Сумерки мамонтов: вымирание ледникового периода и возрождение Америки. Калифорнийский университет Press. С. 78–99. ISBN 978-0-520-23141-2. OCLC 58055404. Получено 2014-11-11.
- ^ Милман, Оливер (6 февраля 2019 г.). «Убийство крупных видов подталкивает их к исчезновению, как показывают исследования». Хранитель. Получено 13 февраля, 2019.
- ^ Ripple, W. J .; и другие. (2019). «Неужели мы до исчезновения поедаем мегафауну мира?». Письма о сохранении: e12627. Дои:10.1111 / conl.12627.
- ^ Алрой, Дж. (12 августа 2008 г.). «Динамика возникновения и исчезновения в морской летописи окаменелостей». PNAS. 105 Дополнение 1 (Дополнение_1): 11536–11542. Bibcode:2008PNAS..10511536A. Дои:10.1073 / pnas.0802597105. ЧВК 2556405. PMID 18695240.
- ^ Wolf, A .; Doughty, C.E .; Малхи, Ю. (2013). "Боковая диффузия питательных веществ млекопитающими травоядными животными в наземных экосистемах". PLoS ONE. 8 (8): e71352. Bibcode:2013PLoSO ... 871352W. Дои:10.1371 / journal.pone.0071352. ЧВК 3739793. PMID 23951141.
- ^ Маршалл, М. (11 августа 2013 г.). «Экосистемы все еще чувствуют боль от вымирания древних людей». Новый ученый. Получено 2013-08-12.
- ^ а б Doughty, C.E .; Wolf, A .; Малхи, Ю. (2013-08-11). «Наследие вымирания мегафауны плейстоцена на доступность питательных веществ в Амазонии». Природа Геонауки. 6 (9): 761–764. Bibcode:2013НатГе ... 6..761Д. Дои:10.1038 / ngeo1895.
- ^ Roman, J .; Маккарти, Дж. Дж. (2010). «Китовый насос: морские млекопитающие повышают первичную продуктивность в прибрежном бассейне». PLOS ONE. 5 (10): e13255. Bibcode:2010PLoSO ... 513255R. Дои:10.1371 / journal.pone.0013255. ЧВК 2952594. PMID 20949007.
- ^ а б c Уилкинсон, Д. М .; Nisbet, E.G .; Ракстон, Г. Д. (2012-05-08). «Мог ли метан, производимый динозаврами-завроподами, способствовать потеплению мезозойского климата?». Текущая биология. 22 (9): R292 – R293. Дои:10.1016 / j.cub.2012.03.042. PMID 22575462. Получено 2012-05-08.
- ^ "Газы динозавров" согрели Землю'". BBC Nature News. 2012-05-07. Получено 2012-05-08.
- ^ а б c Smith, F.A .; Elliot, S.M .; Лайонс, С. К. (23 мая 2010 г.). «Выбросы метана от исчезнувшей мегафауны». Природа Геонауки. 3 (6): 374–375. Bibcode:2010NatGe ... 3..374S. Дои:10.1038 / ngeo877.
- ^ Kelliher, F.M .; Кларк, Х. (15 марта 2010 г.). «Выбросы метана от бизонов - историческая оценка стада Великих равнин Северной Америки». Сельскохозяйственная и лесная метеорология. 150 (3): 473–577. Bibcode:2010AgFM..150..473K. Дои:10.1016 / j.agrformet.2009.11.019.
- ^ Helgen et. все, Кристофер М. (2006). «Экологическое и эволюционное значение размеров гигантских вымерших кенгуру» (PDF). Австралийский зоологический журнал. 54 (4): 293–301. Дои:10.1071 / ZO05077 - через si.edu.
- ^ Ларраменди, А. (2016). «Высота плеч, масса тела и форма хоботков» (PDF). Acta Palaeontologica Polonica. 61 (3): 537–574. Дои:10.4202 / app.00136.2014. S2CID 2092950. Получено 2018-03-22.
- ^ а б Фаринья, Ричард А .; Бискайно, Серджио Ф .; Де Юлис, Джерри (22 мая 2013 г.). Мегафауна: гигантские звери плейстоцена Южной Америки. Издательство Индианского университета. ISBN 978-0-253-00719-3. OCLC 779244424.
- ^ Zhang, Y .; Харрисон, Т. (2017). "Гигантопитек черный: повторное посещение гигантской обезьяны из плейстоцена Азии ". Американский журнал физической антропологии. 162 (S63): 153–177. Дои:10.1002 / ajpa.23150. PMID 28105715.
- ^ Ruff, C. B .; Trinkaus, E .; Холлидей, Т. В. (1997-05-08). «Масса тела и энцефализация в плейстоцене Homo». Природа. 387 (6629): 173–176. Bibcode:1997Натура.387..173р. Дои:10.1038 / 387173a0. PMID 9144286. S2CID 4320413.
- ^ Grine, F.E .; Jumgers, W. L .; Тобиас, П. В .; Пирсон, О. М. (июнь 1995 г.). "Ископаемое Гомо бедренная кость из Берга Аукаса, север Намибии ". Американский журнал физической антропологии. 97 (2): 151–185. Дои:10.1002 / ajpa.1330970207. PMID 7653506.
- ^ Смит, Крис; Бургер, Ли (ноябрь 2007 г.). «Наша история: окаменелости человеческих предков». Голые ученые. Получено 2011-02-19.
- ^ Каппельман, Джон (1997-05-08). «Они могут быть гигантами». Природа. 387 (6629): 126–127. Bibcode:1997Натура.387..126K. Дои:10.1038 / 387126a0. PMID 9144276. S2CID 4328242.
- ^ de Barros Ferraz, K.M.P.M .; Bonach, K .; Вердад, Л. М. (2005). «Связь между массой тела и длиной тела капибар (Hydrochoerus hydrochaeris)". Биота Neotropica. 5 (1): 197–200. Дои:10.1590 / S1676-06032005000100020.
- ^ Китченер, A.C., Брайтенмозер-Вюрстен, К., Эйзирик, Э., Джентри, А., Верделин, Л., Уилтинг, А., Ямагути, Н. (2017). «Пересмотренная таксономия кошачьих: окончательный отчет рабочей группы по классификации кошек Группы специалистов МСОП по кошачьим» (PDF). Новости кошек (Специальный выпуск 11).CS1 maint: использует параметр авторов (ссылка на сайт)
- ^ Брейкфилд, Том (1993). Большие кошки: Королевство мощи. Voyageur Press. п.44. ISBN 978-0-89658-329-0.
- ^ Новелл, Кристин; Джексон, Питер (1996). Дикие кошки: обзор состояния и план действий по сохранению (PDF). Гланд, Швейцария: Группа специалистов по кошкам МСОП / SSC. п. 56. ISBN 978-2-8317-0045-8.
- ^ Китченер А. и Ямагути Н. (2009). «Что такое тигр? Биогеография, морфология и таксономия». В Tilson, R .; Нихус, П. Дж. (Ред.). Тигры мира: наука, политика и сохранение Panthera tigris. Академическая пресса. С. 53–84. ISBN 978-0-08-094751-8.CS1 maint: использует параметр авторов (ссылка на сайт)
- ^ Слагт, Дж. К., Микель, Д. Г., Николаев, И. Г., Гудрич, Дж. М., Смирнов, Э. Н., Трейлор-Хольцер, К., Кристи, С., Арьянова, Т., Смит, Дж. Л. Д. и Карант, К. У. (2005). «Глава 6. Кто царь зверей? Исторические и современные данные о массе тела диких и содержащихся в неволе амурских тигров в сравнении с другими подвидами» (PDF). В Д. Г. Микелле; Э. Н. Смирнов; Дж. М. Гудрич (ред.). Тигры в Сихотэ-Алинском заповеднике: экология и охрана (по-русски). Владивосток, Россия: PSP. С. 25–35.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ http://www.ligerworld.com/samson-the-biggest-tiger.html
- ^ ДеМастер, Д.П .; Стирлинг, И. (8 мая 1981 г.). "Ursus maritimus". Виды млекопитающих (145): 1–7. Дои:10.2307/3504138. JSTOR 3503828.
- ^ Пасичняк-Артс, М. (23 апреля 1993 г.). "Ursus arctos". Виды млекопитающих (439): 1–10. Дои:10.2307/3503828. JSTOR 3504138.
- ^ Soibelzon, L.H .; Шуберт, Б. В. (январь 2011 г.). "Самый большой известный медведь, Arctotherium angustidens, из пампейского региона раннего плейстоцена в Аргентине: с обсуждением размера и диетических тенденций медведей ». Журнал палеонтологии. 85 (1): 69–75. Дои:10.1666/10-037.1. S2CID 129585554. Получено 2011-06-01.
- ^ Свифт, Э. М. (1997-11-17). «Какой у них большой рот: путешественники по Африке, столкнувшиеся с бегемотами, возможно, не доживут до того, чтобы рассказать эту историю». Sports Illustrated Vault. Time Inc. Получено 2011-11-16.
- ^ Перейти ↑ J. Calambokidis и G. Steiger (1998). Синие киты. Voyageur Press. ISBN 0-89658-338-4.
- ^ ^ «Записи животных». Смитсоновский национальный зоологический парк. Проверено 29 мая 2007.
- ^ Антеозавр В архиве 2016-03-14 в Wayback Machine. Palaeos.org (22 апреля 2013 г.)
- ^ Сулей, Т .; Niedźwiedzki, Г. (2019). "Поздний триасовый синапсид размером со слона с прямостоячими конечностями". Наука. 363 (6422): 78–80. Bibcode:2019Научный ... 363 ... 78С. Дои:10.1126 / science.aal4853. PMID 30467179.
- ^ Санкт-Флер, Николас (4 января 2019 г.). «Родственник размером со слона млекопитающих, пасущихся вместе с динозаврами». Нью-Йорк Таймс. Получено 6 января 2019.
- ^ Палмер, Д. (1 июля 2002 г.). Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла. Новые линейные книги. ISBN 978-1-57717-293-2. OCLC 183092423. Получено 2013-06-10.
- ^ Рыба-монстр сокрушила противника самым сильным укусом. Сидней Морнинг Геральд. 30 ноября 2006 г.
- ^ Андерсон, П. С. Л.; Вестнит, М. В. (28 ноября 2006 г.). «Механика кормления и моделирование силы прикуса черепа Dunkleosteus terrelli, древний высший хищник ". Письма о биологии. 3 (1): 77–80. Дои:10.1098 / rsbl.2006.0569. ISSN 1744-9561. ЧВК 2373817. PMID 17443970.
- ^ Андерсон, П.С.Л. (2010-05-04). «Использование моделей сцепления для изучения кинематического разнообразия черепа и функциональной конвергенции в артродермных плакодермах». Журнал морфологии. 271 (8): 990–1005. Дои:10.1002 / jmor.10850. ISSN 0362-2525. PMID 20623651. S2CID 46604512.
- ^ Van Roy, P .; Daley, A.C .; Бриггс, Д. Э. Г. (11 марта 2015 г.). «Гомология туловища и конечностей аномалокарид, выявленная гигантским фильтром с парными створками». Природа. 522 (7554): 77–80. Bibcode:2015Натура.522 ... 77В. Дои:10.1038 / природа14256. PMID 25762145. S2CID 205242881.
- ^ Цубамото, Т. (2012). «Оценка массы тела астрагала у млекопитающих». Acta Palaeontologica Polonica: 259–265. Дои:10.4202 / app.2011.0067. S2CID 54686160.
- ^ Палмер, Д., изд. (1999). Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла. Лондон: Издания Маршалла. п. 248. ISBN 978-1-84028-152-1.
- ^ Мояно, С.Р .; Джаннини, Н. (10.10.2018). «Черепные признаки, связанные с постнатальным развитием хоботка у тапируса (Perissodactyla: Tapiridae) и сравнения с другими современными и ископаемыми копытными млекопитающими». Zoologischer Anzeiger. 277 (7554): 143–147. Дои:10.1016 / j.jcz.2018.08.005. ISSN 0044-5231.
- ^ Образец, Ян (19 февраля 2010 г.). «Большая белая акула находится в большей опасности, чем тигр, - утверждает ученый». Хранитель. Получено 14 августа 2013.