Происхождение птиц - Origin of birds

Научный вопрос о том, в какой большой группе животных птицы эволюционировали традиционно назывался 'происхождение птиц'. Настоящее научный консенсус в том, что птицы группа манирапторан теропод динозавры который возник вовремя Мезозойская эра.
Тесная связь между птицами и динозаврами была впервые предложена в девятнадцатом веке после открытия примитивной птицы. Археоптерикс в Германии. Птицы и вымершие динозавры, не являющиеся птицами, имеют много общих черт скелета.[1] Кроме того, были собраны окаменелости более тридцати видов нептичьих динозавров с сохранившимися перьями. Есть даже очень маленькие динозавры, такие как Микрораптор и Анхиорнис, которые давно, фургон перья рук и ног образуют крылья. Юрский базальный слой Avialan Педопенна также показывает эти длинные перья на ногах. Палеонтолог Лоуренс Уитмер в 2009 году пришли к выводу, что этих данных достаточно, чтобы продемонстрировать, что эволюция птиц прошла стадию с четырьмя крыльями.[2] Ископаемые останки также показывают, что у птиц и динозавров есть общие черты, такие как полость, пневматические кости, гастролиты в пищеварительный система, гнездостроение и задумчивое поведение.
Хотя происхождение птиц исторически спорный вопрос в рамках эволюционная биология, лишь некоторые ученые до сих пор оспаривают динозавровое происхождение птиц, предполагая, что они произошли от других типов птиц. архозавр рептилии. В рамках консенсуса, который поддерживает происхождение динозавров, точная последовательность эволюционных событий, которые привели к появлению ранних птиц в пределах теропод-манирапторан, оспаривается. Происхождение полет птицы это отдельный, но связанный вопрос, на который также есть несколько предложенных ответов.
История исследований
Хаксли, Археоптерикс и ранние исследования

Научное исследование происхождения птиц началось вскоре после публикации в 1859 г. Чарльз Дарвин с О происхождении видов.[3] В 1860 году окаменелое перо было обнаружено в Германия с Поздняя юра Зольнхофенский известняк. Кристиан Эрих Герман фон Мейер описал это перо как Литографический археоптерикс в следующем году.[4] Ричард Оуэн описал почти полный скелет в 1863 году, узнав в нем птицу, несмотря на многие черты, напоминающие рептилии включая когтистые передние конечности и длинный костлявый хвост.[5]
Биолог Томас Генри Хаксли, известный как "Бульдог Дарвина" за его упорную поддержку новой теории эволюции посредством естественного отбора, почти сразу же ухватился за Археоптерикс как переходное ископаемое между птицами и рептилиями. Начиная с 1868 г. и следуя более ранним предложениям Карл Гегенбаур,[6] и Эдвард Дринкер Коуп,[7] Хаксли провел подробное сравнение Археоптерикс с различными доисторическими рептилиями и обнаружил, что он больше всего похож на динозавров, таких как Гипсилофодон и Компсогнат.[8][9] Открытие в конце 1870-х годов культового «берлинского образца» Археоптерикс, в комплекте с набором зубов рептилий, предоставили дополнительные доказательства. Как и Коп, Хаксли предложил эволюционную связь между птицами и динозаврами. Хотя Хаксли был противником очень влиятельного Оуэна, его выводы были приняты многими биологами, включая Барон Франц Нопца,[10] в то время как другие, особенно Гарри Сили,[11] утверждал, что сходство было связано с конвергентная эволюция.
Хейльман и гипотеза текодонта
Переломный момент наступил в начале двадцатого века, когда писали Герхард Хайльманн из Дания. Художник по профессии, Хейльманн проявлял научный интерес к птицам, и с 1913 по 1916 год, развивая более ранние работы Отенио Абель,[12] опубликовал результаты своих исследований в нескольких частях, посвященных анатомии, эмбриология, поведение, палеонтология и эволюция птиц.[13] Его работа, первоначально написанная на Датский в качестве Vor Nuvaerende Viden om Fuglenes Afstamning, был составлен, переведен на английский язык и опубликован в 1926 году как Происхождение птиц.

Как и Хаксли, Хейльманн сравнивал Археоптерикс и других птиц к исчерпывающему списку доисторических рептилий, а также пришли к выводу, что динозавры-тероподы любят Компсогнат были самыми похожими. Однако Хейльманн отметил, что у птиц ключицы (ключицы) слились в кость, называемую фуркула («поперечная кость»), и хотя ключицы были известны у более примитивных рептилий, они еще не были обнаружены у динозавров. Поскольку он твердо верил в Закон Долло, который утверждает, что эволюция необратима, Хейльманн не мог согласиться с тем, что ключицы были потеряны у динозавров и повторно эволюционировали у птиц. Поэтому он был вынужден исключить динозавров как предков птиц и приписать все их сходства конвергентная эволюция. Хейльманн заявил, что предки птиц вместо этого будут найдены среди более примитивных "Текодонт «сорт рептилий.[14] Благодаря чрезвычайно тщательному подходу Хейльмана его книга стала классикой в этой области, а его выводы о происхождении птиц, как и по большинству других тем, были приняты почти всеми эволюционными биологами в течение следующих четырех десятилетий.[15]
Ключицы - относительно хрупкие кости, поэтому они могут быть разрушены или, по крайней мере, повреждены до неузнаваемости. Тем не менее, некоторые ископаемые ключицы теропод действительно были раскопаны до того, как Хейльманн написал свою книгу, но они были неправильно идентифицированы.[16]Отсутствие ключиц у динозавров стало ортодоксальной точкой зрения, несмотря на открытие ключиц у примитивных теропод. Сегизавр в 1936 г.[17] Следующее сообщение о ключицах динозавра было в российской статье в 1983 году.[18]
Вопреки тому, во что верил Хейльманн, палеонтологи теперь признают, что ключицы и, в большинстве случаев, фуркулы являются стандартной чертой не только теропод, но и заурисхий динозавры. Вплоть до конца 2007 г. окостеневшие меховые чешуйки (т.е. из кости, а не хрящ ) были обнаружены у всех типов теропод, кроме самых базальных, Эораптор и Herrerasaurus.[19] Оригинальный отчет о фуркуле у примитивных теропод Сегизавр (1936) был подтвержден повторной экспертизой в 2005 году.[20] Сращенные, похожие на фуркулу ключицы также были обнаружены в Массоспондил, раннеюрский период завроподоморф.[21]
Остром, Дейноних и ренессанс динозавров

Тенденция начала разворачиваться против гипотезы «текодонтов» после открытия в 1964 г. нового динозавра-теропод в г. Монтана. В 1969 году этот динозавр был описан и назван Дейноних к Джон Остром из Йельский университет.[22] В следующем году Остром заново описал образец Птеродактиль в нидерландский язык Музей Тейлера как еще один скелет Археоптерикс.[23] Образец состоял в основном из одного крыла, и его описание заставило Острома понять сходство между запястьями Археоптерикс и Дейноних.[24]
В 1972 г. Британский палеонтолог Алик Уокер выдвинули гипотезу, что птицы произошли не от «текодонтов», а от крокодил предки любят Sphenosuchus.[25] Работа Острома как с тероподами, так и с ранними птицами побудила его ответить серией публикаций в середине 1970-х годов, в которых он изложил многие сходства между птицами и тероподными динозаврами, возродив идеи, впервые выдвинутые Хаксли более века назад.[26][27][28] Признание Остромом динозавров предков птиц, наряду с другими новыми идеями о метаболизме динозавров,[29] уровень активности и родительская забота,[30] началось то, что известно как ренессанс динозавров, которая началась в 1970-х годах и продолжается по сей день.
Откровения Острома также совпали с растущим принятием филогенетической систематики (кладистика ), который начался в 1960-х годах работами Вилли Хенниг.[31] Кладистика - это точный метод распределения видов, основанный строго на их эволюционных отношениях, которые рассчитываются путем определения эволюционного древа, предполагающего наименьшее количество изменений в их анатомических характеристиках. В 1980-х годах кладистическая методология была впервые применена к филогении динозавров. Жак Готье и другие, однозначно показывающие, что птицы были производной группой динозавров-теропод.[32] Ранние исследования показали, что тероподы дромеозаврид, такие как Дейноних были особенно тесно связаны с птицами, и этот результат с тех пор подтверждался много раз.[33][34]
Пернатые динозавры в Китае

В начале 1990-х годов в нескольких странах были обнаружены прекрасно сохранившиеся окаменелости птиц. Раннемеловой период геологические образования в северо-восточной китайской провинции Ляонин.[35][36] В 1996 году китайские палеонтологи описали Синозауроптерикс как новый род птиц из Формация Исянь,[37] но это животное было быстро признано более базальным динозавром-теропод, тесно связанным с Компсогнат. Удивительно, но его тело было покрыто длинными нитевидными структурами. Их окрестили праотцами и считали гомологичный с более развитыми перьями птиц,[38] хотя некоторые ученые не согласны с этой оценкой.[39] Китайский и североамериканский ученые описали Каудиптерикс и Протархеоптерикс вскоре после. Судя по скелетным особенностям, эти животные не были птичьими динозаврами, но их останки имели полностью сформированные перья, очень похожие на перья птиц.[40] "Археораптор ", описанный без экспертная оценка в выпуске 1999 г. Национальная география,[41] оказался контрабандным подделкой,[42] но законные останки продолжают выливаться из Исяня, как законно, так и незаконно. Перья, или «праоперья», были обнаружены у самых разных теропод в Исяне,[43][44] и открытия чрезвычайно похожих на птиц нептичьих динозавров,[45] а также нептичьих динозавроподобных примитивных птиц,[46] почти полностью закрыли морфологический разрыв между нептичьими тероподами и птицами.
Гомология цифр
Есть спор между эмбриологи и палеонтологи ли руки теропод динозавры и птицы принципиально разные, в зависимости от фаланговый counts, счет количества фаланг (пальцев) в руке. Это важная и горячо обсуждаемая область исследования, поскольку ее результаты могут поставить под сомнение консенсус в отношении того, что птицы являются (потомками) динозавров.
Эмбриологи и некоторые палеонтологи, которые выступают против связи между птицами и динозаврами, давно пронумеровали пальцы птиц II-III-IV на основе многочисленных исследований развития в яйце.[47] Это основано на том, что в большинстве амниот, первый палец, который образуется в руке с пятью пальцами, - это палец IV, который образует главную ось. Таким образом, эмбриологи определили первичную ось у птиц как палец IV, а уцелевшие - как II-III-IV. Окаменелости продвинутых теропод (Столбняк ) руки, кажется, имеют цифры I-II-III (некоторые роды в пределах Avetheropoda также имеют уменьшенную цифру IV[48]). Если это так, то развитие пальцев у птиц на II-III-IV является показателем против происхождения теропод (динозавров). Однако без онтогенетический (развитие) основа для окончательного определения, какие пальцы какие на руке теропод (поскольку сегодня не наблюдается роста и развития нептичьих теропод), маркировка руки теропод не является абсолютно окончательной.
Палеонтологи традиционно идентифицировали пальцы птиц как I-II-III. Они утверждают, что пальцы птиц имеют номера I-II-III, как и у тероподных динозавров, согласно законсервированной формуле фаланг. Количество фалангов у архозавров - 2-3-4-5-3; многие родословные архозавров имеют меньшее количество цифр, но те же фаланговая формула в оставшихся цифрах. Другими словами, палеонтологи утверждают, что архозавры разных родов имеют тенденцию терять одни и те же пальцы, когда происходит потеря пальца, снаружи внутрь. Три цифры дромеозавры, и Археоптерикс имеют ту же фаланговую формулу I-II-III, что и I-II-III пальцы базальный архозавры. Следовательно, потеряны цифры V и IV. Если это правда, то современные птицы также будут обладать I-II-III пальцами.[47] Кроме того, в одной публикации 1999 г. было предложено сдвинуть рамки цифр линии теропод, ведущих к птицам (таким образом, превратив цифру I в цифру II, цифру II в цифру III и т.[49][50]Однако такие сдвиги скелета редки у амниот и - чтобы соответствовать тероподному происхождению птиц - должны были произойти исключительно в передних конечностях линии птица-теропод, а не в задних конечностях (состояние, неизвестное ни у одного животного).[51]Это называется Боковое уменьшение цифр (LDR) против Двустороннее сокращение цифр (BDR) (см. Также Лимузавр[52]
Небольшое меньшинство, известное под аббревиатурой ГРУППА (Птицы - не динозавры)[53] включая орнитологов Алан Федучча и Ларри Мартин, продолжает утверждать, что птицы более тесно связаны с более ранними рептилиями, такими как Longisquama или же Euparkeria, чем динозаврам.[54][55] Эмбриологический исследования птицы биология развития подняли вопросы о гомологии пальцев передних конечностей птиц и динозавров.[56] Однако из-за убедительных доказательств, предоставленных сравнительной анатомией и филогенетикой, а также впечатляющих окаменелостей пернатых динозавров из Китая, идея о том, что птицы являются полученный динозавры, которых отстаивал сначала Хаксли, а затем Нопса и Остром, пользуются почти единодушной поддержкой среди сегодняшних палеонтологов.[15]
Гипотеза термогенных мышц
В публикации 2011 года было предложено выбрать для расширения скелетные мышцы, а не эволюция полета, была движущей силой для появления этой клады.[57][58] Мышцы стали крупнее в перспективе эндотермический ящерицы, согласно этой гипотезе, в ответ на потерю позвоночное животное митохондриальный разобщающий белок, UCP1,[59] который термогенный. В млекопитающие, UCP1 функционирует внутри коричневая жировая ткань защитить новорожденных от переохлаждение. У современных птиц скелетные мышцы выполняют аналогичную функцию и, как предполагается, выполняли это у их предков. С этой точки зрения двуногость и другие птичьи скелетный изменения были побочными эффектами мышц гиперплазия, с дальнейшими эволюционными модификациями передних конечностей, включая адаптацию к полету или плаванию, и рудиментарность, являясь вторичными последствиями двуногости.
Филогения
Археоптерикс исторически считалась первой птицей, или Урвогель. Хотя более новые открытия окаменелостей заполнили пробел между тероподами и Археоптерикс, а также разрыв между Археоптерикс и современные птицы, филогенетические систематики по традиции почти всегда используют Археоптерикс в качестве спецификатора, помогающего определить Aves.[60][61] Реже Авес определялся как группа короны состоящий только из современных птиц.[32] Почти все палеонтологи считают птиц целурозавр теропод динозавры.[15] Внутри целурозаврии несколько кладистический анализы нашли поддержку клады названный Манираптора, состоящий из теризинозавроиды, овирапторозавры, троодонтиды, дромеозавриды, и птицы.[33][34][62] Из них дромеозавриды и троодонтиды обычно объединяются в кладу. Дейнонихозаврия, который является сестринская группа птицам (вместе образуют узел-кладу Eumaniraptora ) внутри ствола-клады Паравес.[33][63]
Другие исследования предложили альтернативные филогении, в которых определенные группы динозавров, обычно считающиеся нептичьими, могли развиться от птичьих предков. Например, анализ 2002 года показал, что овирапторозавры были базальными птицами.[64] Альваресзауриды, известный из Азия и Америка, были по-разному классифицированы как базальный манирапторанс,[33][34][65][66] паравианцы[62] родственный таксон орнитомимозавры,[67] а также специализированные ранние пташки.[68][69] Род Рахонавис, первоначально описанный как ранняя пташка,[70] был идентифицирован как дромеозаврид, не являющийся птичьим, в нескольких исследованиях.[63][71] Было высказано предположение, что сами дромеозавриды и троодонтиды располагаются внутри Aves, а не вне его.[72][73]
Особенности связывания птиц и динозавров
Много анатомический[74] черты характерны для птиц и динозавров-теропод.
Перья
Археоптерикс первый хороший образец «пернатого динозавра» был обнаружен в 1861 году. Первый образец был найден в Зольнхофенский известняк на юге Германии, что lagerstätte, редкий и замечательный геологический образование, известное своими великолепно детализированными окаменелостями. Археоптерикс это переходное ископаемое, с чертами, явно промежуточными между чертами нептичьих динозавров-теропод и птицы. Обнаружен всего через два года после открытия Дарвина Происхождение видов, его открытие спровоцировало зарождающиеся дебаты между сторонниками эволюционная биология и креационизм. Эта ранняя пташка настолько похожа на динозавра, что без четкого отпечатка перьев в окружающей скале хотя бы один образец был ошибочно принят за Компсогнат.[75]

С 1990-х годов появился ряд дополнительных пернатые динозавры были обнаружены, что является еще более убедительным доказательством тесной связи между динозаврами и современными птицами. Первые из них изначально были описаны как простые нитевидные праотцы, о которых сообщалось в родословных динозавров столь же примитивных, как компсогнатиды и тираннозавроиды.[76] Однако вскоре после этого были обнаружены перья, неотличимые от перьев современных птиц, и у нептичьих динозавров.[40]
Незначительное меньшинство исследователей заявили, что простые нитевидные структуры «прото-перья» являются просто результатом разложения коллагеновых волокон под кожей динозавров или в плавниках вдоль их спины, и что виды с несомненными перьями, такими как овирапторозавры и дромеозавры не динозавры, а настоящие птицы, не имеющие отношения к динозаврам.[77] Однако большинство исследований пришли к выводу, что пернатые динозавры на самом деле являются динозаврами и что более простые волокна бесспорных теропод представляют собой простые перья. Некоторые исследователи продемонстрировали наличие цветоносности меланин в структурах - чего можно было бы ожидать от перьев, но не коллагеновых волокон.[78] Другие продемонстрировали, используя исследования разложения современных птиц, что даже развитые перья кажутся нитевидными, когда подвергаются сокрушающим силам, испытываемым во время окаменелости, и что предполагаемые «праоперья», возможно, были более сложными, чем считалось ранее.[79] Детальное изучение «праотцов» Sinosauropteryx prima показали, что отдельные перья состоят из центрального пера (рахис) с более тонким колючки отходящие от него, похожие на перья современных птиц, но более примитивные по структуре.[80]

Скелет
Поскольку перья часто ассоциируются с птицами, пернатых динозавров часто рекламируют как пропущенная ссылка между птицами и динозаврами. Однако многочисленные особенности скелета, также общие для этих двух групп, представляют собой более важную связь для палеонтологи. Более того, становится все более очевидным, что отношения между птицами и динозаврами и эволюция полета - более сложные темы, чем предполагалось ранее. Например, хотя когда-то считалось, что птицы произошли от динозавров в результате одной линейной прогрессии, некоторые ученые, в первую очередь Грегори С. Пол, заключаем, что динозавры, такие как дромеозавры возможно, произошли от птиц, потеряв способность летать, сохранив при этом свои перья, аналогичные современным страус и другие ратиты.
Сравнения скелетов птиц и динозавров, а также кладистический анализ, усиливает аргументы в пользу связи, особенно для ветви теропод, называемой манирапторы. Скелетное сходство включает шею, лобок, запястье (полулунный запястный ), рука и грудной пояс, лопатка, ключица, и Грудина.
Исследование, сравнивающее черепа эмбрионов, молодых и взрослых архозавров, пришло к выводу, что черепа птиц являются производными черепов теропод динозавры прогенез, разновидность педоморфной гетерохрония, что привело к сохранению ювенильных характеристик своих предков.[81]
Легкие

Согласно исследованию, проведенному Патриком М. О'Коннором (Patrick M. O'Connor), у крупных динозавров, питающихся мясом, была сложная система воздушных мешочков, аналогичных тем, которые встречаются у современных птиц. Университет Огайо. У динозавров-теропод (плотоядные животные, которые ходили на двух ногах и имели птицеподобные лапы) гибкие воздушные мешочки из мягких тканей, вероятно, перекачивали воздух через жесткие легкие, как в случае с птицами. «То, что когда-то формально считалось уникальным для птиц, в той или иной форме присутствовало у предков птиц», - сказал О'Коннор.[82][83]
Сердце
Компьютерная томография (КТ) сканирование, проведенное в 2000 году грудной клетки образца орнитопод Тескелозавр обнаружили очевидные остатки сложного четырехкамерного сердца, очень похожего на те, что встречаются у современных млекопитающих и птиц.[84] Эта идея вызывает споры в научном сообществе, ее критикуют за плохую анатомию.[85] или просто принятие желаемого за действительное.[86]
В исследовании, опубликованном в 2011 году, к вопросу о личности объекта использовалось несколько направлений исследования, включая более совершенное компьютерное сканирование, гистология, дифракция рентгеновских лучей, Рентгеновская фотоэлектронная спектроскопия, и сканирующий электрон микроскопия. С помощью этих методов авторы обнаружили, что: внутренняя структура объекта не включает камеры, а состоит из трех несвязанных областей материала с более низкой плотностью и не сопоставима со структурой страус сердце; "стены" состоят из осадочный минералы, которые, как известно, не производятся в биологических системах, такие как гетит, полевой шпат минералы, кварц, и гипс, а также некоторые фрагменты растений; углерод, азот, и фосфор, химические элементы важные для жизни, отсутствовали в их образцах; клеточные структуры сердца отсутствовали. Был один возможный участок с клеточными структурами животных. Авторы обнаружили, что их данные подтверждают идентификацию как конкрецию песка из погребальной среды, а не из сердца, с возможностью сохранения отдельных участков тканей.[87]
Однако вопрос о том, как эта находка отражает скорость метаболизма и внутреннюю анатомию динозавра, является спорным, независимо от личности объекта.[87] Оба современные крокодилы и птицы, самые близкие из ныне живущих родственников динозавров, имеют четырехкамерные сердца (хотя и модифицированные у крокодилов), так что динозавры, вероятно, тоже имели их; структура не обязательно связана со скоростью метаболизма.[88]
Поза во сне
Окаменелости троодонты Мэй и Sinornithoides демонстрируют, что динозавры спали, как некоторые современные птицы, закрыв голову руками.[89] Такое поведение, которое могло помочь согреть голову, также характерно для современных птиц.
Репродуктивная биология
При откладывании яиц у самок на конечностях вырастают особые кости. Этот медуллярный Кость образует богатый кальцием слой внутри твердой внешней кости и используется в качестве источника кальция для изготовления яичной скорлупы. Наличие костных тканей эндостального происхождения, выстилающих внутренние полости костного мозга частей тиранозавр Рекс задняя конечность экземпляра предполагала, что T. rex использовали аналогичные репродуктивные стратегии и выяснили, что это самка.[90] Дальнейшие исследования обнаружили у тероподов костный мозг. Аллозавр и орнитопод Тенонтозавр. Потому что линейка динозавров, в которую входят Аллозавр и Тиранозавр отклонился от линии, ведущей к Тенонтозавр На очень раннем этапе эволюции динозавров это говорит о том, что динозавры в целом производили мозговую ткань.[91]
Вынашивание и забота о молодых

Несколько Читипати экземпляры были найдены покоящимися над яйцами в гнезде в положении, наиболее напоминающем задумчивый.[92]
Например, многочисленные виды динозавров. Майасаура, были обнаружены в стадах, где смешивались как очень молодые, так и взрослые особи, что свидетельствует о тесном взаимодействии между ними.
Эмбрион динозавра был найден без зубов, что свидетельствует о необходимости некоторой родительской заботы, чтобы накормить молодого динозавра, возможно, взрослый динозавр срыгнул пищу в рот молодого динозавра (видеть альтриальный ). Такое поведение наблюдается у многих видов птиц; родительские птицы срыгивают пищу в рот птенца.
Камни в желудке
И птицы, и динозавры используют желудок камни. Эти камни глотаются животными, чтобы помочь пищеварению и расщеплению пищи и твердых волокон, когда они попадают в желудок. При обнаружении в сочетании с окаменелости, камни в желудке называются гастролиты.[93]Камни желудка также встречаются у некоторых рыб (кефаль, грязь, а гиллару, разновидность форели) и у крокодилов.
Молекулярные доказательства
В нескольких случаях заявлялось об извлечении ДНК и белков из окаменелостей мезозойских динозавров, что позволяет проводить сравнение с птицами. Несколько белки предположительно были обнаружены в окаменелостях динозавров,[94] включая гемоглобин.[95]
В мартовском выпуске 2005 г. Наука, Доктор Мэри Хигби Швейцер и ее команда объявила об открытии гибкого материала, напоминающего настоящие мягкие ткани, внутри 68-миллионного человека. тиранозавр Рекс нога кость образца MOR 1125 из Формация Адского ручья в Монтана. Семь коллаген типы, полученные из костных фрагментов, по сравнению с данными о коллагене от живых птиц (в частности, курица ), предполагают, что более старые теропод и птицы тесно связаны между собой.[96] Мягкие ткани позволили провести молекулярное сравнение сотовый анатомия и секвенирование белков из коллаген ткани, опубликованные в 2007 году, оба из которых указывают на то, что T. rex и птицы более тесно связаны друг с другом, чем с Аллигатор.[97][98] Второе молекулярное исследование убедительно подтвердило связь птиц с динозаврами, хотя не поместило птиц в теропод, как ожидалось. В этом исследовании использовалось восемь дополнительных последовательностей коллагена, извлеченных из бедренной кости "мумифицированного" Брахилофозавр канадский образец MOR 2598, а гадрозавр.[99] Однако эти результаты были очень противоречивыми. Нет другого пептиды мезозойского возраста. В 2008 году было высказано предположение, что предполагаемая мягкая ткань на самом деле является бактериальной микропленкой.[100] В ответ было заявлено, что именно эти микропленки защищают мягкие ткани.[101] Другое возражение заключалось в том, что результаты могли быть вызваны загрязнением.[102] В 2015 году в более контролируемых условиях, защищающих от загрязнения, пептиды все же были идентифицированы.[103] В 2017 году исследование показало, что в кости современного страуса присутствует пептид, идентичный найденному в кости Тиранозавр и Брахилофозавр образцы, подчеркивающие опасность перекрестного заражения.[104]
Об успешном извлечении древней ДНК из окаменелостей динозавров сообщалось в двух разных случаях, но после дальнейшего изучения и экспертная оценка, ни одно из этих сообщений не может быть подтверждено.[105]
Происхождение птичьего полета
Споры о происхождении птичьего полета почти так же стары, как и идея о том, что птицы произошли от динозавры, возникшие вскоре после открытия Археоптерикс в 1862 году. С тех пор в большинстве дискуссий доминировали две теории: беглая («с нуля») теория предполагает, что птицы произошли от маленьких, быстрых хищников, бегавших по земле; Древесная («с деревьев вниз») теория предполагает, что полет с приводом от двигателя произошел от планирования без двигателя древесных (лазающих по деревьям) животных. Более поздняя теория, «бег с помощью крыльев по наклонной» (WAIR), является вариантом бегущей теории и предполагает, что крылья развивали свое аэродинамический функционирует в результате необходимости быстро подниматься по очень крутым склонам, таким как деревья, что поможет маленьким пернатым динозаврам убежать от хищников.
В марте 2018 года ученые сообщили, что Археоптерикс вероятно был способен полет, но способом, существенно отличным от современные птицы.[106][107]
Беглая («с нуля») теория

Беглая теория происхождения полета была впервые предложена Сэмюэл Венделл Уиллистон, и разработан Барон Нопца. Эта гипотеза предполагает, что некоторые быстро бегающие животные с длинными хвостами использовали свои руки, чтобы сохранять равновесие во время бега. Современные версии этой теории во многих деталях отличаются от версии Уиллистона-Нопса, главным образом в результате открытий, сделанных со времен Нопсы.
Нопса предположил, что увеличение площади вытянутых рук могло помочь маленьким бегающим хищникам сохранять равновесие, а чешуя предплечий удлинялась, развивающийся в перья. Перья также могли использоваться для ловли насекомых или другой добычи. Постепенно животные прыгали на большие расстояния, чему способствовали их развивающиеся крылья. Нопса также предложил три стадии эволюции полета. Во-первых, животные развили пассивный полет, при котором развивающиеся структуры крыльев служили своего рода парашют. Во-вторых, они достигли активного полета за счет взмахов крыльев. Он использовал Археоптерикс как пример этого второго этапа. Наконец, птицы получили способность парить.[108]

В настоящее время считается, что перья произошли не из чешуи, поскольку перья состоят из разных белки.[109] Более серьезно, теория Нопса предполагает, что перья эволюционировали как часть эволюции полета, и недавние открытия доказывают, что это предположение неверно.
Перья очень распространены в целурозавры динозавры (включая ранние тираннозавроид Дилонг ).[110] Современное птицы классифицируются как целурозавры почти всеми палеонтологами,[111] хотя и не несколькими орнитологи.[112][113][114] Современная версия гипотезы «с нуля» утверждает, что предки птиц были маленькими, пернатый, наземные хищные динозавры (скорее похожие на Roadrunners в их охотничьем стиле[115]), которые использовали свои передние конечности для равновесия при преследовании добычи, и что передние конечности и перья позже эволюционировали таким образом, что они обеспечивали скольжение, а затем и движение. Наиболее широко предлагаемые оригинальные функции перьев включают теплоизоляцию и конкурентоспособность, как у современных птиц.[116][117]
Все Археоптерикс окаменелости происходят из морских отложений, и было высказано предположение, что крылья могли помогать птицам бегать по воде, как Иисус Христос Ящерица (обыкновенный василиск ).[118]
Самые последние опровержения гипотезы «с нуля» - это попытка опровергнуть современную версию предположения о том, что птицы являются модифицированными целурозаврами динозавров. Самые сильные атаки основаны на эмбриологические анализы Отсюда следует вывод, что крылья птиц образуются из цифр 2, 3 и 4 (что соответствует указательному, среднему и безымянному пальцам человека. Первая из трех пальцев птицы образует Alula, которые они используют, чтобы избежать торможение в полете на малой скорости - например, при посадке). Однако руки целурозавров состоят из 1, 2 и 3 пальцев (большой и первые два пальца у людей).[119] Однако эти эмбриологические анализы немедленно подверглись сомнению на том эмбриологическом основании, что «рука» часто развивается по-разному в клады которые потеряли несколько пальцев в ходе своей эволюции, и что «руки» птиц действительно развиваются из пальцев 1, 2 и 3.[120][121][122] Этот спор сложен и еще не решен - см. "Гомология цифр ".
Бег по наклонной с помощью крыльев
В бег по наклонной с помощью крыльев (WAIR) гипотеза была вызвана наблюдением молодых чукар цыплят, и предлагает, чтобы крылья развили свои аэродинамический функционирует в результате необходимости быстро подниматься по очень крутым склонам, таким как стволы деревьев, например, чтобы спастись от хищников.[123] Это делает ее специализированным типом беглой («с нуля») теории. Обратите внимание, что в этом сценарии птицам нужно прижимная сила чтобы дать ногам больший захват.[124][125] Но ранние пташки, в том числе Археоптерикс, не хватало плечевой механизм с помощью которых крылья современных птиц производят быстрые и мощные взмахи вверх. Поскольку прижимная сила, от которой зависит WAIR, создается восходящими ударами, похоже, что ранние пташки были неспособны к WAIR.[126] Поскольку WAIR - это поведенческая черта без остеологической специализации, филогенетическое расположение гребка полета до расхождения Neornithes, группа, которая включает всех существующих птиц, делает невозможным определение того, является ли WAIR предком птичьего полета или производным от него.[127]
Древесная («с деревьев вниз») теория

Большинство версий древесной гипотезы утверждают, что предки птиц были очень маленькими динозаврами, которые жили на деревьях, прыгая с ветки на ветку. У этого маленького динозавра уже были перья, которые эволюция использовала для создания более длинных и жестких форм, которые использовались в аэродинамике, в конечном итоге создавая крылья. Затем крылья развивались и становились все более совершенными в качестве устройств, позволяющих прыгунам больше контролировать, прыгать с парашютом, планировать и летать ступенчато. Древесная гипотеза также отмечает, что для древесных животных аэродинамика гораздо более энергоэффективна, поскольку такие животные просто падают, чтобы достичь минимальной скорости планирования.[128][129]
Некоторые маленькие динозавры юрского или раннего мелового периода, все с перьями, были интерпретированы как возможно имеющие древесные и / или аэродинамические приспособления. К ним относятся Скансориоптерикс, Эпидексиптерикс, Микрораптор, Педопенна, и Анхиорнис. Анхиорнис особенно важен для этого предмета, поскольку он жил в начале поздней юры, задолго до Археоптерикс.[130]
Анализ пропорций костей пальцев самых примитивных птиц. Археоптерикс и Конфуциусорнис по сравнению с таковыми у живых видов, предполагают, что ранние виды могли жить как на земле, так и на деревьях.[131]
Одно исследование показало, что самые ранние птицы и их непосредственные предки не лазили по деревьям. Это исследование показало, что кривизна когтей на лапах ранних птиц больше похожа на то, что наблюдается у современных наземных птиц, чем у птиц, сидящих на насесте.[132]
Сниженное значение Археоптерикс
Археоптерикс был первым и долгое время единственным известным пернатым Мезозойский животное. В результате обсуждение эволюции птиц и их полета сосредоточилось на Археоптерикс по крайней мере, до середины 1990-х.
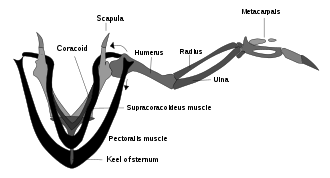
Были споры о том, Археоптерикс действительно мог летать. Оказалось, что Археоптерикс имели структуры мозга и датчики баланса внутреннего уха, которые птицы используют для управления своим полетом.[133] Археоптерикс также имел расположение перьев крыльев, как у современных птиц, и аналогично асимметричные маховые перья на крыльях и хвосте. Но Археоптерикс не хватало плечевой механизм при котором крылья современных птиц производят быстрые и мощные взмахи вверх (см. диаграмму выше надкожного шкива); это может означать, что он и другие ранние птицы были неспособны к взмаху крыльев и могли только парить.[126]
Но открытие с начала 1990-х годов многих пернатые динозавры Значит это Археоптерикс больше не является ключевой фигурой в эволюции птичьего полета. Другие мелкие пернатые целурозавры из Меловой и поздно Юрский период показать возможные предвестники птичьего полета. К ним относятся Рахонавис, наземный бегун с Велоцираптор -поднятый серповидный коготь на втором пальце ноги, который, по мнению некоторых палеонтологов, был лучше приспособлен для полета, чем Археоптерикс,[134] Скансориоптерикс, древесный динозавр, который может поддерживать теорию "с деревьев вниз",[135] и Микрораптор, древесный динозавр, возможно, способный летать, но если это так, то он больше похож на биплан, так как у него были хорошо развитые перья на ногах.[136] Еще в 1915 году некоторые ученые утверждали, что эволюция полета птиц могла происходить через четырехкрылый (или тетраптерикс) сцена.[137][138]
Вторичная нелетаемость у динозавров
| Целурозавры |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Группы, обычно рассматриваемые как птицы, выделены жирным шрифтом.[73]
Гипотеза, приписываемая Грегори Пол и проповедовал в своих книгах Хищные динозавры мира (1988) и Динозавры Воздуха (2002), предполагает, что некоторые группы нелетающих хищных динозавров, особенно деинонихозавры, но, возможно, другие, такие как овирапторозавры, теризинозавры, Альваресзауриды и орнитомимозавры - спускаются от птиц. Пол также предположил, что птица-предок этих групп была более развита в своих летных адаптациях, чем Археоптерикс. Гипотеза означала бы, что Археоптерикс менее тесно связан с современными птицами, чем эти динозавры.[139]
Гипотеза Пола получила дополнительную поддержку, когда Майр и другие. (2005) проанализировали новый, десятый образец Археоптерикс, и пришел к выводу, что Археоптерикс была сестрой рода Deinonychosauria, но более развитая птица Конфуциусорнис был внутри Dromaeosauridae. Этот результат подтверждает гипотезу Пола, предполагающую, что Deinonychosauria и Troodontidae являются частью Aves, собственно птичьего происхождения и, во-вторых, нелетающими.[140] В этот документ, однако, были исключены все другие птицы, и поэтому не проводилась выборка распределения их признаков. Статья подверглась критике со стороны Корфа и Батлера (2006), которые обнаружили, что авторы не могут подтвердить свои выводы статистически. Mayr и другие. согласился, что статистическая поддержка была слабой, но добавила, что она также слабая для альтернативных сценариев.[141]
Текущий кладистический анализ не поддерживают гипотезу Павла о позиции Археоптерикс. Вместо этого они указывают, что Археоптерикс ближе к птицам, внутри клады Avialae, чем дейнонихозавров или овирапторозавров. Однако некоторые окаменелости подтверждают версию этой теории, согласно которой у некоторых нелетающих хищных динозавров могли быть летающие предки. Особенно, Микрораптор, Педопенна, и Анхиорнис все имеют крылатые ноги, имеют много общих черт и лежат близко к основанию клады Паравес. Это говорит о том, что предком паравианцев был четырехкрылый планер, и что более крупные дейнонихозавры вторично утратили способность летать, в то время как аэродинамические способности у птиц увеличивались по мере своего развития.[2] Дейноних может также проявлять частичную подвижность, при этом детеныши способны летать или планировать, а взрослые - нелетающие.[142] В 2018 году исследование показало, что последний общий предок Pennaraptora имел суставные поверхности на пальцах и между плюсневой костью и запястьем, которые были оптимизированы для стабилизации руки в полете. Это рассматривалось как указание на вторичную нелетаемость у тяжелых базальных представителей этой группы.[143]
В Euornithes, самым ранним однозначным примером вторичной нелетаемости является Патагоптерикс.[144]
Смотрите также
![]() Портал динозавров
Портал динозавров
Сноски
- ^ Чиаппе, Луис М. (2009). «Уменьшенные динозавры: эволюционный переход к современным птицам». Эволюция: образование и пропаганда. 2 (2): 248–256. Дои:10.1007 / s12052-009-0133-4.
- ^ а б Витмер, LM (2009). «Палеонтология: пернатые динозавры в клубке». Природа. 461 (7264): 601–2. Bibcode:2009Натура 461..601Вт. Дои:10.1038 / 461601a. PMID 19794481. S2CID 205049989.
- ^ Дарвин, Чарльз Р. (1859). О происхождении видов путем естественного отбора или о сохранении избранных рас в борьбе за жизнь. Лондон: Джон Мюррей. п. 502 стр. ISBN 978-1-4353-9386-8.
- ^ фон Мейер, К.Е. Германн. (1861). "Литографический археоптерикс (Фогель-Федер) и Птеродактиль фон Зольнхофен ". Neues Jahrbuch für Mineralogie, Geologie und Paläontologie (на немецком). 1861: 678–679.
- ^ Оуэн, Ричард. (1863). "На Археоптерикс [sp] фон Мейера, с описанием окаменелостей длиннохвостого вида, из литографического камня Соленхофена [sp] ". Философские труды Лондонского королевского общества. 153: 33–47. Дои:10.1098 / рстл.1863.0003.
- ^ Гегенбаур, К. (1863). "Vergleichend-anatomische Bemerkungen über das Fußskelet der Vögel". Archiv für Anatomie, Physiologie und Wissenschaftliche Medicin. 1863: 450–472.
- ^ Коуп, Э. (1867 г.). «Рассказ о вымерших рептилиях, которые приблизились к птицам». Труды Академии естественных наук Филадельфии. 19: 234–235.связь
- ^ Хаксли, Т. (1868). «О животных, которые занимают промежуточное положение между птицами и рептилиями». Летопись и журнал естественной истории. 4-й. 2: 66–75.
- ^ Хаксли, Томас Х. (1870). «Еще одно доказательство близости динозавров рептилий и птиц». Ежеквартальный журнал Лондонского геологического общества. 26 (1–2): 12–31. Дои:10.1144 / GSL.JGS.1870.026.01-02.08. S2CID 129125487.
- ^ Нопца, Франц. (1907). «Идеи о происхождении полета». Труды Лондонского зоологического общества: 223–238.
- ^ Сили, Гарри Г. (1901). Драконы воздуха: описание вымерших летающих рептилий. Лондон: Methuen & Co., стр. 239pp.
- ^ Абель, О. (1911). "Die Vorfahren der Vögel und ihre Lebensweise". Verhandlungen der Zoologisch-Botanischen Gesellschaft в Вене. 61: 144–191.
- ^ Nieuwland, Ilja J.J. (2004). "Герхард Хейльманн и взгляд художника в науке, 1912-1927 гг." (PDF). Журнал PalArch по палеонтологии позвоночных. 3 (2).
- ^ Хейльманн, Герхард (1926). Происхождение птиц. Лондон: Уизерби. п.208 стр.. ISBN 978-0-486-22784-9.
- ^ а б c Падиан, Кевин. (2004). «Базальные авиалины». В Weishampel, Дэвид Б.; Додсон, Питер; Osmólska, Halszka (ред.). Динозаврия (Второе изд.). Беркли: Калифорнийский университет Press. С. 210–231. ISBN 978-0-520-24209-8.
- ^ Например, в 1923 году, за три года до книги Хейльманса, Рой Чепмен Эндрюс нашел хороший Овираптор ископаемое в Монголия, но Генри Фэрфилд Осборн, которые проанализировали окаменелость в 1924 году, ошибочно идентифицировали фуркулу как межключица; описано в Пол, Г.С. (2002). Динозавры воздуха: эволюция и потеря полета у динозавров и птиц. JHU Press. ISBN 978-0-8018-6763-7.
- ^ Кэмп, Чарльз Л. (1936). «Новый тип маленьких динозавров-теропод из песчаника навахо в Аризоне». Бюллетень Департамента геологических наук Калифорнийского университета. 24: 39–65.
- ^ В Овираптор: Барсболд Р. (1983). «Плотоядные динозавры из мелового периода Монголии». Труды Суместная Советско-Монгольская Палеонтогическая Экспедиция (на русском). 19: 1–117. См. Резюме и изображения на "Желание Целофиз". Архивировано из оригинал 3 февраля 2008 г.
- ^ Липкин, К., Серено, П.С., и Хорнер, Дж. Р. (ноябрь 2007 г.). «Фуркула в зухомиме тенеренсисе и тираннозавре рекс (динозавр: теропода: столбняк)». Журнал палеонтологии. 81 (6): 1523–1527. Дои:10.1666/06-024.1. S2CID 86234363.CS1 maint: несколько имен: список авторов (связь) - полный текст в настоящее время онлайн на "Фуркула в Зухомим Tenerensis и тиранозавр Рекс". 2007-11-11. Архивировано из оригинал на 2011-05-22. Получено 2008-04-17. Здесь перечислено большое количество теропод, у которых были обнаружены меховые раковины, а также описаны таковые из Зухомим Tenerensis и тиранозавр Рекс.
- ^ Каррано, М.Р., Хатчинсон, Дж. Р., Сэмпсон, С. (Декабрь 2005 г.). "Новая информация о Segisaurus halli, маленький динозавр-теропод из ранней юры Аризоны " (PDF). Журнал палеонтологии позвоночных. 25 (4): 835–849. Дои:10.1671 / 0272-4634 (2005) 025 [0835: NIOSHA] 2.0.CO; 2. ISSN 0272-4634. Архивировано из оригинал (PDF) на 2006-05-12.CS1 maint: несколько имен: список авторов (связь)
- ^ Йейтс, Адам М .; и Васконселос, Сесилио К. (2005). "Фуркуловидные ключицы у прозауроподных динозавров Массоспондил". Журнал палеонтологии позвоночных. 25 (2): 466–468. Дои:10.1671 / 0272-4634 (2005) 025 [0466: FCITPD] 2.0.CO; 2. ISSN 0272-4634.
- ^ Остром, Джон Х. (1969). «Остеология Deinonychus antirrhopus, необычный теропод из нижнего мела Монтаны ». Бюллетень музея естественной истории Пибоди. 30: 1–165.
- ^ Остром, Джон Х. (1970). "Археоптерикс: Уведомление о «новом» экземпляре. Наука. 170 (3957): 537–8. Bibcode:1970Sci ... 170..537O. Дои:10.1126 / science.170.3957.537. PMID 17799709. S2CID 46682559.
- ^ Чемберс, Пол (2002). Кости раздора: скандалы с археоптериксами. Лондон: John Murray Ltd., стр. 183–184. ISBN 978-0-7195-6054-5.
- ^ Уокер, Алик Д. (1972). «Новый взгляд на происхождение птиц и крокодилов». Природа. 237 (5353): 257–263. Bibcode:1972 год. 237..257 Вт. Дои:10.1038 / 237257a0. S2CID 4174690.
- ^ Остром, Джон Х. (1973). «Родословная птиц». Природа. 242 (5393): 136. Bibcode:1973НФС..242..136О. Дои:10.1038 / 242136a0. S2CID 29873831.
- ^ Остром, Джон Х. (1975). Происхождение птиц. Ежегодный обзор наук о Земле и планетах. 3. С. 55–77. Bibcode:1975 АРЕПС ... 3 ... 55О. Дои:10.1146 / annurev.ea.03.050175.000415. ISBN 978-0-912532-57-8.
- ^ Остром, Джон Х. (1976). "Археоптерикс и происхождение птиц ». Биологический журнал Линнеевского общества. 8 (2): 91–182. Дои:10.1111 / j.1095-8312.1976.tb00244.x.
- ^ Баккер, Роберт Т. (1972). «Анатомические и экологические свидетельства эндотермии у динозавров». Природа. 238 (5359): 81–85. Bibcode:1972 г.Натура 238 ... 81Б. Дои:10.1038 / 238081a0. S2CID 4176132.
- ^ Хорнер, Джон Р.; Макела, Роберт (1979). «Гнездо молоди свидетельствует о семейной структуре динозавров». Природа. 282 (5736): 296–298. Bibcode:1979Натура.282..296H. Дои:10.1038 / 282296a0. S2CID 4370793.
- ^ Хенниг, Э. Вилли (1966). Филогенетическая систематика. перевод Дэвиса, Д. Дуайта; & Зангерл, Райнер. Урбана: Университет Иллинойса Press. ISBN 978-0-252-06814-0.
- ^ а б Готье, Жак. (1986). «Заурисхиевая монофилия и происхождение птиц». В Padian, Кевин (ред.). Происхождение птиц и эволюция полета. Мемуары Калифорнийской академии наук 8. С. 1–55.
- ^ а б c d Сентер, Фил (2007). «Новый взгляд на филогению целурозаврии (Dinosauria: Theropoda)». Журнал систематической палеонтологии. 5 (4): 429–463. Дои:10.1017 / S1477201907002143. S2CID 83726237.
- ^ а б c Тернер, Алан Х .; Хван, Санни; И Норелл, Марк А. (2007). «Маленький теропод из Оёша, ранний мел, Байхангор, Монголия». Американский музей Novitates. 3557 (1): 1–27. Дои:10.1206 / 0003-0082 (2007) 3557 [1: ASDTFS] 2.0.CO; 2. HDL:2246/5845. ISSN 0003-0082.CS1 maint: несколько имен: список авторов (связь)
- ^ Серено, Пол К.; Рао Ченган (1992). «Ранняя эволюция полета птиц и насестов: новые данные из нижнего мела Китая». Наука. 255 (5046): 845–8. Bibcode:1992Sci ... 255..845S. Дои:10.1126 / science.255.5046.845. PMID 17756432. S2CID 41106677.
- ^ Хоу Лянь-Хай, Лянь-Хай; Чжоу Чжунхэ; Мартин, Ларри Д.; & Федучча, Алан (1995). «Птица с клювом из юрского периода Китая». Природа. 377 (6550): 616–618. Bibcode:1995Натура.377..616H. Дои:10.1038 / 377616a0. S2CID 4357707.CS1 maint: несколько имен: список авторов (связь)
- ^ Цзи Цян; Цзи Шу-ань (1996). «Об открытии самой ранней окаменелости птиц в Китае и происхождении птиц» (PDF). Китайская геология. 233: 30–33.
- ^ Чэнь Пей-цзи, Пей-цзи; Дун Чжимин; И Чжэнь Шуо-нань. (1998). «Исключительно сохранившийся динозавр теропод из формации Исянь в Китае». Природа. 391 (6663): 147–152. Bibcode:1998Натура.391..147C. Дои:10.1038/34356. S2CID 4430927.CS1 maint: несколько имен: список авторов (связь)
- ^ Lingham-Soliar, Theagarten; Федучча, Алан; И Ван Сяолинь. (2007). «Новый китайский образец показывает, что« праоперья »в раннемеловом тероподном динозавре Sinosauropteryx представляют собой деградированные коллагеновые волокна». Труды Королевского общества B. 274 (1620): 1823–9. Дои:10.1098 / rspb.2007.0352. ЧВК 2270928. PMID 17521978.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Цзи Цян, Филип Дж .; Карри, Филип Дж.; Норелл, Марк А .; & Цзи Шу-ань. (1998). «Два пернатых динозавра из северо-восточного Китая» (PDF). Природа. 393 (6687): 753–761. Bibcode:1998Натура.393..753Q. Дои:10.1038/31635. S2CID 205001388.CS1 maint: несколько имен: список авторов (связь)
- ^ Слоан, Кристофер П. (1999). "Перья для T. rex?". Национальная география. 196 (5): 98–107. Архивировано из оригинал на 2012-10-11.
- ^ Монастерский, Ричард (2000). «Все смешалось из-за птиц и динозавров». Новости науки. 157 (3): 38. Дои:10.2307/4012298. JSTOR 4012298. Архивировано из оригинал на 2008-10-03. Получено 2008-07-04.
- ^ Сюй Син, Син; Тан Чжи-лу; И Ван Сяолинь. (1999). «Динозавр теризинозаврид с покровными структурами из Китая». Природа. 399 (6734): 350–354. Bibcode:1999Натура.399..350X. Дои:10.1038/20670. S2CID 204993327.CS1 maint: несколько имен: список авторов (связь)
- ^ Сюй Син, X; Норелл, Марк А .; Куанг Сюэвэнь; Ван Сяолинь; Чжао Ци; И Цзя Чэнкай. (2004). «Базальные тиранозавроиды из Китая и свидетельства праотцов у тираннозавроидов». Природа. 431 (7009): 680–4. Bibcode:2004Натура 431..680X. Дои:10.1038 / природа02855. PMID 15470426. S2CID 4381777.CS1 maint: несколько имен: список авторов (связь)
- ^ Сюй Син, X; Чжоу Чжунхэ; Ван Сяолинь; Куанг Сюэвэнь; Чжан Фучэн; И Ду Сянкэ (2003). «Четырехкрылые динозавры из Китая». Природа. 421 (6921): 335–40. Bibcode:2003Натура.421..335X. Дои:10.1038 / природа01342. PMID 12540892. S2CID 1160118.CS1 maint: несколько имен: список авторов (связь)
- ^ Чжоу, Чжунхэ; Чжан Фучэн (2002). «Длиннохвостая птица, питающаяся семенами, из раннего мелового периода Китая». Природа. 418 (6896): 405–9. Bibcode:2002Натурал.418..405Z. Дои:10.1038 / природа00930. PMID 12140555. S2CID 4423299.
- ^ а б Чаттерджи, Санкар (17 апреля 1998 г.). «Подсчет пальцев птиц и динозавров». Наука. 280 (5362): 355a – 355. Bibcode:1998Sci ... 280..355C. Дои:10.1126 / science.280.5362.355a.
- ^ Университет Мэриленда домашняя страница отдела геологии, "Theropoda I" на Avetheropoda, 14 июля 2006 г.
- ^ Wagner, G.P .; Готтье, Дж. А. (1999). «1,2,3 = 2,3,4: решение проблемы гомологии цифр на руке птицы». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 96 (9): 5111–6. Bibcode:1999PNAS ... 96,5111 Вт. Дои:10.1073 / пнас.96.9.5111. ЧВК 21825. PMID 10220427.
- ^ Научные блоги: Лимузавр это превосходно В архиве 2009-06-22 на Wayback Machine.
- ^ Биология развития 8e Online. Глава 16: Птицы произошли от динозавров? В архиве 2008-01-18 на Wayback Machine
- ^ Варгас А.О., Вагнер Г.П. и Готье Я. 2009. Лимузавр и идентификация пальца птицы. Доступно в Nature Precedings [1]
- ^ Havstad, Joyce C .; Смит, Н. Адам (2019). «Ископаемые с перьями и философия науки». Систематическая биология. 68 (5): 840–851. Дои:10.1093 / sysbio / syz010. ISSN 1076-836X. ЧВК 6701454. PMID 30753719.
- ^ Мартин, Ларри Д. (2004). «Базальное архозавровое происхождение птиц». Acta Zoologica Sinica. 50 (6): 978–990.
- ^ Федучча, Алан; Lingham-Soliar, Theagarten; И Хинклифф, Дж. Ричард. (2005). «Существуют ли пернатые динозавры? Проверка гипотезы на неонтологических и палеонтологических данных». Журнал морфологии. 266 (2): 125–66. Дои:10.1002 / jmor.10382. PMID 16217748. S2CID 15079072.CS1 maint: несколько имен: список авторов (связь)
- ^ Берк, Энн С.; Федучча, Алан (1997). «Модели развития и идентификация гомологий в птичьей руке». Наука. 278 (5338): 666–668. Bibcode:1997Sci ... 278..666B. Дои:10.1126 / science.278.5338.666.
- ^ Ньюман С.А. (2011). «Термогенез, гиперплазия мышц и происхождение птиц». BioEssays. 33 (9): 653–656. Дои:10.1002 / bies.201100061. PMID 21695679. S2CID 42012034.
- ^ Ньюман С.А., Мезенцева Н.В., Бадяев А.В. (2013). «Потеря генов, термогенез и происхождение птиц». Летопись Нью-Йоркской академии наук. 1289 (1): 36–47. Bibcode:2013НЯСА1289 ... 36Н. Дои:10.1111 / няс.12090. PMID 23550607.
- ^ Мезенцева Н.В., Кумаратилаке Ю.С., Ньюман С.А. (2008). «Путь дифференцировки коричневых адипоцитов у птиц: эволюционный путь не пройден». BMC Биология. 6 (1): 17. Дои:10.1186/1741-7007-6-17. ЧВК 2375860. PMID 18426587.
- ^ Серено, Пол К. (1997). «Происхождение и эволюция динозавров». Ежегодный обзор наук о Земле и планетах. 25 (1): 435–489. Bibcode:1997AREPS..25..435S. Дои:10.1146 / annurev.earth.25.1.435.
- ^ Чиаппе, Луис М. (1997). «Авес». В Карри, Филип Дж.; Падиан, Кевин (ред.). Энциклопедия динозавров. Сан-Диего: Academic Press. С. 45–50. ISBN 978-0-12-226810-6.
- ^ а б Хольц, Томас Р.; Осмольска, Гальска. (2004). «Заурисхия». В Weishampel, Дэвид Б.; Додсон, Питер; Osmólska, Halszka (ред.). Динозаврия (Второе изд.). Беркли: Калифорнийский университет Press. С. 21–24. ISBN 978-0-520-24209-8.
- ^ а б Тернер, Алан Х .; Пол, Диего; Кларк, Джулия А .; Эриксон, Грегори М .; И Норелл, Марк А. (2007). «Базальный дромеозаврид и эволюция размера, предшествующая полету птиц». Наука. 317 (5843): 1378–81. Bibcode:2007Научный ... 317.1378Т. Дои:10.1126 / science.1144066. PMID 17823350.CS1 maint: несколько имен: список авторов (связь)
- ^ Осмольска, Гальска; Марьянская, Тереза; & Wolsan, Mieczysław. (2002). «Статус Авиалана для Овирапторозаврии». Acta Palaeontologica Polonica. 47 (1): 97–116.CS1 maint: несколько имен: список авторов (связь)
- ^ Martinelli, Agustín G .; Вера, Эсекьель I. (2007). "Achillesaurus manazzonei, новый теропод альварессаурид (Dinosauria) из позднемеловой формации Бахо-де-ла-Карпа, провинция Рио-Негро, Аргентина " (PDF). Zootaxa. 1582: 1–17. Дои:10.11646 / zootaxa.1582.1.1.
- ^ Новас, Фернандо Э.; Пол, Диего. (2002). «Отношения с Альваресзавридом пересмотрены». In Chiappe, Luis M .; Витмер, Лоуренс М. (ред.). Птицы мезозоя: над головами динозавров. Беркли: Калифорнийский университет Press. С. 121–125. ISBN 978-0-520-20094-4.
- ^ Серено, Пол К. (1999). «Эволюция динозавров». Наука. 284 (5423): 2137–47. Дои:10.1126 / science.284.5423.2137. PMID 10381873.
- ^ Перле, Алтангерель; Норелл, Марк А .; Chiappe, Луис М .; И Кларк, Джеймс М. (1993). «Нелетающая птица из мелового периода Монголии». Природа. 362 (6421): 623–626. Bibcode:1993Натура.362..623A. Дои:10.1038 / 362623a0. S2CID 4252852.CS1 maint: несколько имен: список авторов (связь)
- ^ Chiappe, Луис М .; Норелл, Марк А .; И Кларк, Джеймс М. (2002). "Короткорукие Alvarezsauridae мелового периода: Мононикус и его родственники ». In Chiappe, Luis M .; Witmer, Lawrence M. (eds.). Птицы мезозоя: над головами динозавров. Беркли: Калифорнийский университет Press. С. 87–119. ISBN 978-0-520-20094-4.CS1 maint: несколько имен: список авторов (связь)
- ^ Форстер, Кэтрин А .; Сэмпсон, Скотт Д.; Chiappe, Луис М .; И Краузе, Дэвид В. (1998). «Тероподное происхождение птиц: новые свидетельства из позднего мела Мадагаскара». Наука. 279 (5358): 1915–9. Bibcode:1998Научный ... 279.1915F. Дои:10.1126 / science.279.5358.1915. PMID 9506938.CS1 maint: несколько имен: список авторов (связь)
- ^ Маковицкий, Питер Дж .; Апестегия, Себастьян; И Аньолин, Федерико Л. (2005). «Самый ранний теропод-дромеозаврид из Южной Америки». Природа. 437 (7061): 1007–11. Bibcode:2005Натура 437.1007М. Дои:10.1038 / природа03996. PMID 16222297. S2CID 27078534.CS1 maint: несколько имен: список авторов (связь)
- ^ Пол, Грегори С. (2002). Динозавры воздуха: эволюция и потеря полета у динозавров и птиц. Балтимор: Издательство Университета Джона Хопкинса. п.472 стр.. ISBN 978-0-8018-6763-7.
- ^ а б Майр, Джеральд; Поль, Буркхард; И Петерс, Д. Стефан (2005). "Хорошо сохранившийся Археоптерикс экземпляр с чертами теропод ». Наука. 310 (5753): 1483–6. Bibcode:2005Научный ... 310.1483M. Дои:10.1126 / наука.1120331. PMID 16322455. S2CID 28611454.CS1 maint: несколько имен: список авторов (связь)
- ^ Чаттерджи, Иммур; Л. Иммур (9 сентября 2005 г.). "Динозавры из фильмов о Парке Юрского периода". Geolor.com. Архивировано из оригинал 12 мая 2008 г.. Получено 23 июня, 2007.
- ^ Велнхофер, П. (1988). "Ein neuer Exemplar von Археоптерикс". Археоптерикс. 6: 1–30.
- ^ Сюй X, Норелл М.А., Куанг X, Ван X, Чжао Ц., Цзя Ц. (октябрь 2004 г.). «Базальные тиранозавроиды из Китая и свидетельства праотцов у тираннозавроидов». Природа. 431 (7009): 680–4. Bibcode:2004Натура 431..680X. Дои:10.1038 / природа02855. PMID 15470426. S2CID 4381777.
- ^ Федучча, А. (2012). Загадка пернатых драконов: Скрытые птицы Китая. Издательство Йельского университета, ISBN 0-300-16435-1, ISBN 978-0-300-16435-0
- ^ Zhang, F .; Kearns, S.L .; Орр, П.Дж .; Benton, M.J .; Чжоу, З .; Johnson, D .; Сюй, X .; и Ван, X. (2010). «Окаменелые меланосомы и цвет меловых динозавров и птиц» (PDF). Природа. 463 (7284): 1075–1078. Bibcode:2010 Натур.463.1075Z. Дои:10.1038 / природа08740. PMID 20107440. S2CID 205219587.CS1 maint: несколько имен: список авторов (связь)
- ^ Фот, С. (2012). «Об идентификации структур перьев у представителей линии стебля птиц: данные по окаменелостям и актуопалеонтологии». Paläontologische Zeitschrift. 86: 91–102. Дои:10.1007 / s12542-011-0111-3. S2CID 86362907.
- ^ Currie, P.J .; Чен, П.-ж. (2001). "Анатомия Sinosauropteryx prima из Ляонина, Северо-Восточный Китай ". Канадский журнал наук о Земле. 38 (1): 705–727. Bibcode:2001CaJES..38.1705C. Дои:10.1139 / cjes-38-12-1705.
- ^ Bhullar, B.A. S .; Маруган-Лобон, Дж. С .; Racimo, F .; Бевер, Г. С .; Rowe, T. B .; Норелл, М. А .; Абжанов, А. (2012). «У птиц есть педоморфные черепа динозавров». Природа. 487 (7406): 223–226. Bibcode:2012Натура.487..223Б. Дои:10.1038 / природа11146. PMID 22722850. S2CID 4370675.
- ^ О'Коннор, П.М.; Claessens, L.P.A.M. (2005). «Базовая конструкция легких птиц и проточная вентиляция у динозавров-теропод, не являющихся птицами». Природа. 436 (7048): 253–6. Bibcode:2005Натура.436..253O. Дои:10.1038 / природа03716. PMID 16015329. S2CID 4390587.
- ^ Пол С. Серено, Рикардо Н. Мартинес, Джеффри А. Уилсон, Дэвид Дж. Варриккио, Оскар А. Алкобер, Ханс К. Э. Ларссон (2008). Кемп, Том (ред.). «Доказательства наличия птичьих внутригрудных воздушных мешков у нового хищного динозавра из Аргентины». PLOS ONE. 3 (9): e3303. Bibcode:2008PLoSO ... 3.3303S. Дои:10.1371 / journal.pone.0003303. ЧВК 2553519. PMID 18825273.CS1 maint: несколько имен: список авторов (связь)
- ^ Фишер, П. Э .; Рассел, Д. А .; Stoskopf, M. K .; Barrick, R.E .; Молоток, М .; Кузьмиц, А.А. (2000). «Сердечно-сосудистые доказательства средней или высокой скорости метаболизма у орнитисхийских динозавров». Наука. 288 (5465): 503–5. Bibcode:2000Sci ... 288..503F. Дои:10.1126 / science.288.5465.503. PMID 10775107.
- ^ Hillenius, W. J .; Рубен, Дж. А. (2004). «Эволюция эндотермии у наземных позвоночных: кто? Когда? Почему?». Физиологическая и биохимическая зоология. 77 (6): 1019–42. Дои:10.1086/425185. PMID 15674773.
- ^ Rowe, T .; McBride, E. F .; Sereno, P.C .; Рассел, Д. А .; Фишер, П. Э .; Barrick, R.E .; Стоскопф, М. К. (2001). «Динозавр с каменным сердцем». Наука. 291 (5505): 783. Дои:10.1126 / science.291.5505.783a. PMID 11157158.
- ^ а б Cleland, Timothy P .; Stoskopf, Майкл К .; и Швейцер, Мэри Х. (2011). «Гистологическое, химическое и морфологическое исследование« сердца »небольшого позднемелового периода. Тескелозавр". Naturwissenschaften. 98 (3): 203–211. Bibcode:2011NW ..... 98..203C. Дои:10.1007 / s00114-010-0760-1. PMID 21279321. S2CID 2408562.CS1 maint: несколько имен: список авторов (связь)
- ^ Чинсами, Анусуя; и Хиллениус, Виллем Дж. (2004). «Физиология неавианских динозавров». Динозаврия, 2-й. 643–659.
- ^ Сюй, X .; Норелл, М.А. (2004). «Новый динозавр троодонтид из Китая со спящей позой, напоминающей птицу». Природа. 431 (7010): 838–41. Bibcode:2004Натура.431..838X. Дои:10.1038 / природа02898. PMID 15483610. S2CID 4362745. См. Комментарий к статье
- ^ Schweitzer, M.H .; Wittmeyer, J.L .; и Хорнер, Дж. Р. (2005). "Половая репродуктивная ткань у крысят и тиранозавр Рекс". Наука. 308 (5727): 1456–60. Bibcode:2005Научный ... 308.1456С. Дои:10.1126 / наука.1112158. PMID 15933198. S2CID 30264554.CS1 maint: несколько имен: список авторов (связь)
- ^ Ли, Эндрю Х .; Вернинг, Сара (2008). «Половая зрелость у растущих динозавров не соответствует моделям роста рептилий». Труды Национальной академии наук. 105 (2): 582–7. Bibcode:2008ПНАС..105..582Л. Дои:10.1073 / pnas.0708903105. ЧВК 2206579. PMID 18195356.
- ^ Норелл, М.А., Кларк, Дж. М., Дашзевег, Д., Барсболд, Т., Чиаппе, Л. М., Дэвидсон, А. Р., МакКенна, М. К. и Новачек, М. Дж. (Ноябрь 1994 г.). «Эмбрион динозавра теропод и сходство яиц динозавров Пылающих Скал». Наука. 266 (5186): 779–82. Bibcode:1994Наука ... 266..779N. Дои:10.1126 / science.266.5186.779. PMID 17730398. S2CID 22333224.CS1 maint: несколько имен: список авторов (связь)
- ^ Крылья О (2007). «Обзор функции гастролита с последствиями для ископаемых позвоночных и пересмотренная классификация». Palaeontologica Polonica. 52 (1): 1–16.
- ^ Embery, G; Милнер, AC; Waddington, RJ; Холл, ЖК; Лэнгли, MS; Милан, AM (2003). «Идентификация белкового материала в кости динозавра игуанодона». Соединительная ткань исследования. 44 (Дополнение 1): 41–6. Дои:10.1080/03008200390152070. PMID 12952172. S2CID 2249126.
- ^ Schweitzer, MH; Маршалл, М; Каррон, К; Боле, Д.С.; Буссе, Южная Каролина; Арнольд, EV; Барнард, Д.; Хорнер, младший; Старки, младший (июнь 1997 г.). «Соединения гема в губчатой кости динозавра». Труды Национальной академии наук Соединенных Штатов Америки. 94 (12): 6291–6. Bibcode:1997PNAS ... 94.6291S. Дои:10.1073 / пнас.94.12.6291. ЧВК 21042. PMID 9177210.
- ^ Schweitzer, MH; Wittmeyer, JL; Хорнер, младший; Топорский, JK (2005). "Сосуды мягких тканей и сохранение клеток у тираннозавра". Наука. 307 (5717): 1952–5. Bibcode:2005Научный ... 307.1952С. Дои:10.1126 / science.1108397. PMID 15790853. S2CID 30456613. Также охватывает Параграф "Биология репродукции" в разделе "Пернатые динозавры и птицы".
- ^ Асара, JM; Швейцер М.Х., Фреймарк Л.М., Филлипс М., Cantley LC (2007). «Последовательности белков мастодонта и тираннозавра Rex, выявленные с помощью масс-спектрометрии». Наука. 316 (5822): 280–5. Bibcode:2007Наука ... 316..280A. Дои:10.1126 / science.1137614. PMID 17431180. S2CID 85299309.CS1 maint: несколько имен: список авторов (связь)
- ^ Орган, CL; Schweitzer, MH; Чжэн, Вт; Freimark, LM; Cantley, LC; Асара, Дж. М. (2008). «Молекулярная филогенетика мастодонта и тиранозавр Рекс". Наука. 320 (5875): 499. Дои:10.1126 / science.1154284. PMID 18436782. S2CID 24971064.
- ^ Schweitzer, M. H .; Zheng W., Organ CL, Avci R., Suo Z., Freimark LM, Lebleu VS, Duncan MB, Vander Heiden MG, Neveu JM, Lane WS, Cottrell JS, Horner JR, Cantley LC, Kalluri R. & Asara JM ( 2009 г.). «Биомолекулярная характеристика и белковые последовательности кампанского Hadrosaur B. canadensis». Наука. 324 (5927): 626–31. Bibcode:2009Sci ... 324..626S. Дои:10.1126 / science.1165069. PMID 19407199. S2CID 5358680.CS1 maint: несколько имен: список авторов (связь)
- ^ Kaye, TG; Гоглер, G; Савлович, З (2008). «Мягкие ткани динозавров интерпретируются как бактериальные биопленки». PLOS ONE. 3 (7): e2808. Bibcode:2008PLoSO ... 3,2808 тыс.. Дои:10.1371 / journal.pone.0002808. ЧВК 2483347. PMID 18665236.
- ^ Петерсон, Дж. Э .; Lenczewski, ME; Шерер, Р.П. (2010). «Влияние микробных биопленок на сохранение первичных мягких тканей у ископаемых и современных архозавров». PLOS ONE. 5 (10): e13334. Bibcode:2010PLoSO ... 513334P. Дои:10.1371 / journal.pone.0013334. ЧВК 2953520. PMID 20967227.
- ^ Берн, М; Финни, BS; Гольдберг, Д. (2009). "Повторный анализ тиранозавр Рекс масс-спектры ». Журнал протеомных исследований. 8 (9): 4328–4332. Дои:10.1021 / pr900349r. ЧВК 2738754. PMID 19603827.
- ^ Cleland, TP; и другие. (2015). «Масс-спектрометрия и определение характеристик кровеносных сосудов на основе антител Брахилофозавр канадский". Журнал протеомных исследований. 14 (12): 5252–5262. Дои:10.1021 / acs.jproteome.5b00675. ЧВК 4768904. PMID 26595531.
- ^ Бакли, Майкл; Уорвуд, Стейси; ван Донген, Барт; Китченер, Эндрю С .; Мэннинг, Филлип Л. (2017). «Химера ископаемого белка; трудности в различении пептидных последовательностей динозавров от современного перекрестного заражения». Труды Королевского общества B. 284 (1855): 20170544. Дои:10.1098 / rspb.2017.0544. ЧВК 5454271. PMID 28566488.
- ^ Wang, HL; Ян, З.Ы .; Джин, Д.Й. (1997). «Повторный анализ опубликованной последовательности ДНК, амплифицированной из окаменелостей яиц динозавров мелового периода». Молекулярная биология и эволюция. 14 (5): 589–91. Дои:10.1093 / oxfordjournals.molbev.a025796. PMID 9159936.
- ^ Voeten, Dennis F.A.E .; и другие. (13 марта 2018 г.). «Геометрия костей крыла свидетельствует об активном полете археоптерикса». Nature Communications. 9 (923): 923. Bibcode:2018НатКо ... 9..923В. Дои:10.1038 / s41467-018-03296-8. ЧВК 5849612. PMID 29535376.
- ^ Гуарино, Бен (13 марта 2018 г.). «Этот пернатый динозавр, наверное, летал, но не как любая птица, которую вы знаете». Вашингтон Пост. Получено 13 марта 2018.
- ^ Террес, Джон К. (1980). Энциклопедия североамериканских птиц от Общества Одюбона. Нью-Йорк, штат Нью-Йорк: Кнопф. стр.398–401. ISBN 978-0-394-46651-4.
- ^ Полинг, Дж. (1996). «Перья, щитки и происхождение птиц». dinosauria.com. Архивировано из оригинал на 2008-04-02. Получено 2008-03-21.
- ^ Прум, Р. и Браш, А.Х. (2002). «Эволюционное происхождение и разнообразие перьев» (PDF). Ежеквартальный обзор биологии. 77 (3): 261–95. Дои:10.1086/341993. PMID 12365352. Архивировано из оригинал (PDF) на 2003-10-15. Получено 2018-09-17.
- ^ Майр Г., Поль Б., Петерс Д. (2005). "Хорошо сохранившийся Археоптерикс экземпляр с чертами теропод ». Наука. 310 (5753): 1483–6. Bibcode:2005Научный ... 310.1483M. Дои:10.1126 / наука.1120331. PMID 16322455. S2CID 28611454.
- ^ Федучча, А. (1999). Происхождение и эволюция птиц. Издательство Йельского университета. ISBN 978-0-300-07861-9.
- ^ Федучча, А. (февраль 1995 г.). «Взрывная эволюция третичных птиц и млекопитающих». Наука. 267 (5198): 637–8. Bibcode:1995Научный ... 267..637F. Дои:10.1126 / science.267.5198.637. PMID 17745839. S2CID 42829066. Архивировано из оригинал на 2008-03-20.
- ^ Федучча, А. (1993).[уточнить ]
- ^ Меловой были найдены следы птицы с похожим образом жизни - Локли, М.Г., Ли, Р., Харрис, Д.Д., Мацукава, М., и Лю, М. (август 2007 г.). «Самые ранние зигодактильные птичьи лапы: свидетельства из следов раннего мелового дорожного бегуна». Naturwissenschaften. 94 (8): 657–65. Bibcode:2007NW ..... 94..657L. Дои:10.1007 / s00114-007-0239-x. PMID 17387416. S2CID 15821251.CS1 maint: несколько имен: список авторов (связь)
- ^ Бюргерс, П. и Л. М. Чиаппе (1999). «Крыло археоптерикса как основной генератор тяги». Природа. 399 (6731): 60–62. Bibcode:1999Натура.399 ... 60Б. Дои:10.1038/19967. S2CID 4430686.
- ^ Коуэн, Р. История Жизни. Blackwell Science. ISBN 978-0-7266-0287-0.
- ^ Виделер, Дж. Дж. 2005: Птичий полет. Оксфордский университет. Press, Оксфорд.
- ^ Берк, A.C. и Feduccia, A. (1997). «Модели развития и идентификация гомологий в птичьей руке». Наука. 278 (5338): 666–668. Bibcode:1997Sci ... 278..666B. Дои:10.1126 / science.278.5338.666. Обобщено на «Исследования эмбрионов показывают, что динозавры не могли стать популярными среди современных птиц». ScienceDaily. Октябрь 1997 г.
- ^ Чаттерджи, С. (апрель 1998 г.). «Подсчет пальцев птиц и динозавров». Наука. 280 (5362): 355a – 355. Bibcode:1998Sci ... 280..355C. Дои:10.1126 / science.280.5362.355a.
- ^ Варгас, А.О., Фэллон, Дж. Ф. (октябрь 2004 г.). «У птиц есть крылья динозавра: молекулярные доказательства». Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития. 304B (1): 86–90. Дои:10.1002 / jez.b.21023. PMID 15515040.CS1 maint: несколько имен: список авторов (связь)
- ^ Пенниси, Э. (январь 2005 г.). «Крылья птиц действительно похожи на руки динозавров» (PDF). Наука. 307 (5707): 194b. Дои:10.1126 / science.307.5707.194b. PMID 15653478. S2CID 82490156. Архивировано из оригинал (PDF) 27 июля 2011 г.
- ^ Есть видеоклип, в котором очень молодая цыпочка делает это в «Бег с опорой на крыло и эволюция полета».
- ^ Циферблат, К. (2003). «Бег на наклонной поверхности с крыльями и эволюция полета». Наука. 299 (5605): 402–4. Bibcode:2003Наука ... 299..402D. Дои:10.1126 / science.1078237. PMID 12532020. S2CID 40712093. Обобщено в Морелль, Ребекка (24 января 2008 г.). «Раскрыты секреты птичьего полета» (Интернет). Ученые считают, что они могут быть на шаг ближе к разгадке тайны того, как первые птицы поднялись в воздух.. Новости BBC. Получено 2008-01-25.
- ^ Связка, M.W & Dial, K.P. (2003). «Механика бега по наклонной плоскости с крыльями (WAIR)» (PDF). Журнал экспериментальной биологии. 206 (Pt 24): 4553–64. Дои:10.1242 / jeb.00673. PMID 14610039. S2CID 6323207.
- ^ а б Сентер, П. (2006). «Ориентация лопатки у теропод и базальных птиц и происхождение машущего полета». Acta Palaeontologica Polonica. 51 (2): 305–313.
- ^ Dececchi, T. Alexander; Ларссон, Ханс К. Э. (2011). «Оценка древесных адаптаций антецедентов птиц: проверка экологических условий происхождения птичьего полетного инсульта». PLOS ONE. 6 (8): e22292. Bibcode:2011PLoSO ... 622292D. Дои:10.1371 / journal.pone.0022292. ЧВК 3153453. PMID 21857918.
- ^ Чаттерджи, Санкар, Темплин, Р.Дж. (2004) «Пернатые целурозавры из Китая: новый взгляд на древесное происхождение полета птиц», стр. 251-281. В «Пернатых драконах: исследования перехода от динозавров к птицам» (П. Дж. Карри, Э. Б. Коппельхус, М. А. Шугар и Дж. Л. Райт (ред.). Издательство Индианского университета, Блумингтон.
- ^ Тарситано, Самуэль Ф .; Рассел, Энтони П .; Хорн, Фрэнсис; Пламмер, Кристофер; Миллерчип, Карен (2000). «Об эволюции перьев с точки зрения аэродинамики и конструкции». Американский зоолог. 40 (4): 676–686. Дои:10.1093 / icb / 40.4.676.
- ^ Hu, D .; Hou, L .; Чжан, Л. и Сюй, X. (2009). "ПредварительноАрхеоптерикс Troodontid теропод из Китая с длинными перьями на плюсне ". Природа. 461 (7264): 640–3. Bibcode:2009Натура 461..640H. Дои:10.1038 / природа08322. PMID 19794491. S2CID 205218015.
- ^ Хопсон, Джеймс А. "Экоморфология пропорций фаланг тероподов птиц и нонавий: последствия для древесного и наземного происхождения птичьего полета" (2001) с новых перспектив происхождения и ранней эволюции птиц: материалы Международного симпозиума в честь Иоанна Х. Остром. Дж. Готье и Л. Ф. Галл, ред. Нью-Хейвен: Пибоди Мус. Nat. Hist., Yale Univ. ISBN 0-912532-57-2© 2001 Музей естественной истории Пибоди, Йельский университет. Все права защищены.
- ^ Глен, К. И Беннетт, М. (Ноябрь 2007 г.). «Режимы кормления мезозойских птиц и нептичьих теропод». Текущая биология. 17 (21): R911–2. Дои:10.1016 / j.cub.2007.09.026. PMID 17983564. S2CID 535424. Архивировано из оригинал на 2012-12-08.
- ^ Алонсо, П.Д., Милнер, А.К., Кетчем, Р.А., Коксон, М.Дж., Роу, Т. (Август 2004 г.). «Птичья природа мозга и внутреннего уха археоптерикса». Природа. 430 (7000): 666–9. Bibcode:2004Натура 430..666А. Дои:10.1038 / природа02706. PMID 15295597. S2CID 4391019.CS1 maint: несколько имен: список авторов (связь)
- ^ Чиаппе, Л.М. (02.02.2007). Прославленные динозавры: происхождение и ранняя эволюция птиц. Сидней: UNSW Press. ISBN 978-0-471-24723-4.
- ^ Чжан, Ф., Чжоу, З., Сюй, X. и Ван, X. (2002). «Молодой теропод целурозавра из Китая указывает на древесные привычки». Naturwissenschaften. 89 (9): 394–8. Bibcode:2002NW ..... 89..394Z. Дои:10.1007 / s00114-002-0353-8. PMID 12435090. S2CID 556221.CS1 maint: несколько имен: список авторов (связь)
- ^ Чаттерджи, S; Templin, RJ (2007). "Форма крыла биплана в плане и летные характеристики пернатого динозавра Microraptor gui" (PDF). Труды Национальной академии наук. 104 (5): 1576–80. Bibcode:2007PNAS..104.1576C. Дои:10.1073 / pnas.0609975104. ЧВК 1780066. PMID 17242354.
- ^ Биби, К. У. А. (1915). «Этап тетраптерикса в потомстве птиц». Zoologica. 2: 38–52.
- ^ Лукас, Фредерик А. (Frederic Augustus); Американский музей естественной истории (13 июня 2018 г.). Животные прошлого: описание некоторых существ древнего мира. Нью-Йорк: [амер. Mus. Пресса] - через Интернет-архив.
- ^ Пол, Г.С. (2002). «Динозавры воздуха: эволюция и потеря полета у динозавров и птиц». Балтимор: Издательство Университета Джона Хопкинса. стр. 257
- ^ Майр, Г. (2006). "Ответ на комментарий к Хорошо сохранившийся образец археоптерикса с чертами теропод". Наука. 313 (5791): 1238c. Bibcode:2006Научный ... 313.1238M. Дои:10.1126 / наука.1130964.
- ^ Corfe, I.J .; Батлер, Р.Дж. (2006). "Комментировать Хорошо сохранившийся образец археоптерикса с чертами теропод". Наука. 313 (5791): 1238b. Bibcode:2006Научный ... 313.1238C. Дои:10.1126 / наука.1130800. PMID 16946054. S2CID 19859320.
- ^ Парсонс, Уильям Л .; Парсонс, Кристен М. (2015). «Морфологические вариации в онтогенезе Deinonychus antirrhopus (Theropoda, Dromaeosauridae)». PLOS ONE. 10 (4): e0121476. Bibcode:2015PLoSO..1021476P. Дои:10.1371 / journal.pone.0121476. ЧВК 4398413. PMID 25875499.
- ^ Hutson, Joel D .; Хатсон, Кельда Н. (2018). «Сохранение адаптированного к полету комплекса птичьих суставов пальцев у страуса помогает определить, когда крылья начали развиваться у динозавров». Страус: журнал африканской орнитологии. 89 (2): 173–186. Дои:10.2989/00306525.2017.1422566. S2CID 91081348.
- ^ Чиаппе, Луис (01.01.2002), Остеология нелетающего Patagopteryx deferrariisi из позднего мела Патагонии (Аргентина), стр. 281–316, ISBN 978-0-520-20094-4, получено 2020-07-27
Рекомендации
- Барсболд, Ринчен (1983): О птичьих чертах в строительстве хищных динозавров. [«Птичьи» особенности в морфологии хищных динозавров]. Труды Объединенной Советско-Монгольской палеонтологической экспедиции 24: 96-103. [Оригинал статьи на русском языке.] Перевод В. Роберта Уэлша, копия предоставлена Кеннетом Карпентером и преобразована Мэтью Каррано. Полный текст PDF
- Боренштейн, Сет (31 июля 2014 г.). «Исследование прослеживает эволюцию динозавров до первых птиц». AP Новости. Получено 3 августа, 2014.
- Боствик, Кимберли S (2003). «Происхождение и эволюция птиц: данные накапливаются, ученые объединяют, но« споры »все еще бушуют». Кладистика. 19 (4): 369–371. Дои:10.1016 / S0748-3007 (03) 00069-0.
- Дингус, Лоуэлл и Роу, Тимоти (1997): Ошибочное вымирание: эволюция динозавров и происхождение птиц. В. Х. Фриман и компания, Нью-Йорк. ISBN 0-7167-2944-X
- Динозаврия онлайн (1995): Связь археоптерикса с современными птицами. Проверено 30 сентября 2006.
- Динозаврия онлайн (1996): Синапоморфии динозавров, обнаруженные в Археоптерикс. Проверено 30 сентября 2006.
- Хейльманн, Г. (1926): Происхождение птиц. Уизерби, Лондон. ISBN 0-486-22784-7 (1972 г., Dover переиздание)
- Майр, Джеральд; Pohl, B .; Питерс, Д. С. (2005). "Хорошо сохранившийся Археоптерикс Образец с чертами теропод ». Наука. 310 (5753): 1483–1486. Bibcode:2005Научный ... 310.1483M. Дои:10.1126 / наука.1120331. PMID 16322455. S2CID 28611454.
- Олсон, Сторрс Л. (1985): Летопись окаменелостей птиц. В: Фарнер, Д.С.; Кинг, Дж. Р. и Паркс, Кеннет К. (ред.): Биология птиц 8: 79-238. Academic Press, Нью-Йорк.
внешняя ссылка
- «Динозавры среди нас» повторяют эволюционный путь от динозавров к птицам, NY Times, 28 марта 2016 г.
- DinoBuzz Популярное обсуждение гипотезы динозавров-птиц.
- Археоптерикс - Часто задаваемые вопросы из группы новостей Usenet talk.origins.
- Динозавры среди нас Статья и видео Американский музей естественной истории выставка эволюции динозавров, ведущих к птицам
| Викискладе есть медиафайлы по теме Окаменелости авеса. |
| Плавники |  | |
|---|---|---|
| Конечности | ||
| Крылья | ||
| Эволюция | ||
| Связанный | ||