Урегулирование Америки - Settlement of the Americas
В поселение в Америке принято считать, что началось, когда Палеолит охотники-собиратели вошел Северная Америка от Северная Азия Мамонт степной через Берингия сухопутный мост, которые образовались между северо-восточными Сибирь и западный Аляска из-за снижения уровень моря вовремя Последний ледниковый максимум.[1]Эти популяции расширились к югу от Ледяной щит Лаурентиды и быстро распространилась по обоим Северная и Южная Америка, 14 000 лет назад.[2][3][4][5] Самые ранние популяции в Северной и Южной Америке, примерно до 10 000 лет назад, известны как Палеоиндейцы.
Население Америки - давний открытый вопрос, и пока археология, Плейстоцен геология, физическая антропология, и Анализ ДНК проливают все больше света на эту тему, но важные вопросы остаются нерешенными.[6] Хотя существует общее мнение о том, что Америка впервые была заселена из Азии, характер миграции, ее время и место (а) происхождения в Евразии народов, мигрировавших в Америку, остаются неясными.[3]
Преобладающие модели миграции описывают разные временные рамки для миграции из Азии из Берингов пролив и последующее расселение основателей по всему континенту.[7] Коренные народы Северной и Южной Америки были связаны с сибирским населением лингвистические факторы, распределение группы крови, И в генетический состав как отражено молекулярный данные, такие как ДНК.[8][9]
«Первая теория Хлодвига» относится к гипотезе 1950-х годов о том, что Хлодвиг культура представляет собой самое раннее присутствие человека в Америке, начавшееся около 13 000 лет назад; свидетельства существования докловисских культур накапливаются с 2000 года, отодвигая возможную дату первого заселения Америки на 33 тысячи лет назад.[10][11][12]
Окружающая среда во время последнего оледенения
Для ознакомления с методами радиоуглеродного датирования, используемыми археологами и геологами, см. радиоуглеродное датирование.
Возникновение и погружение Берингии.
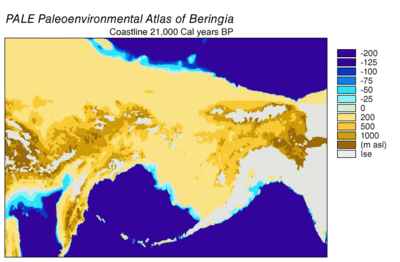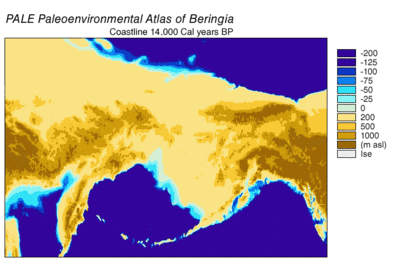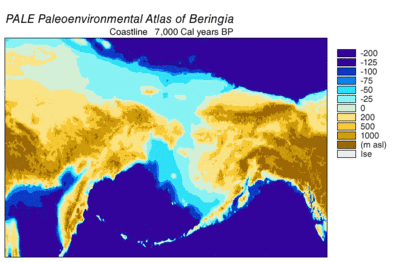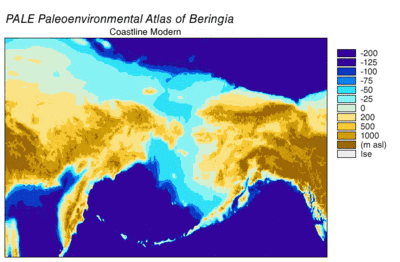
Вовремя Висконсинское оледенение, океанская вода Земли в разной степени с течением времени хранилась в ледник лед. По мере накопления воды в ледниках объем воды в океанах соответственно уменьшался, в результате чего глобальный уровень моря. Изменение уровня моря во времени реконструировано с использованием изотоп кислорода анализ глубоководных кернов, датирование морских террас и отбор проб изотопов кислорода с высоким разрешением из океанских бассейнов и современных ледяных шапок. Капля эвстатический уровень моря примерно на 60–120 метров (от 200 до 390 футов) от современных уровней, начиная примерно с 30 000 лет BP, созданный Берингия, прочный и обширный географический объект, соединяющий Сибирь с Аляской.[13] С подъемом уровня моря после Последний ледниковый максимум (LGM) Берингийский сухопутный мост снова был затоплен. Оценки окончательного повторного затопления Берингийского сухопутного моста основаны исключительно на текущих данных. батиметрия Берингова пролива и эвстатическая кривая уровня моря помещают событие около 11000 лет назад (Рисунок 1). Текущие исследования, восстанавливающие палеогеографию Берингии во время дегляциации, могут изменить эту оценку, а возможное более раннее погружение может еще больше ограничить модели миграции людей в Северную Америку.[13]
Ледники
Наступление последнего ледникового максимума спустя 30 000 лет назад привело к расширению альпийских ледников и континентальных ледяных щитов, которые блокировали пути миграции из Берингии. К 21000 лет назад и, возможно, тысячам лет назад Кордильер и Laurentide ледяные щиты срослись к востоку от скалистые горы, перекрывая потенциальный путь миграции в центр Северной Америки.[14][15][16] Альпийские ледники прибрежных хребтов и Полуостров Аляска изолировал внутреннюю часть Берингии от побережья Тихого океана. Прибрежный альпийские ледники и доли кордильерского льда слились в предгорные ледники которые покрывали большие участки береговой линии на юг до Остров Ванкувер и образовал ледяную ленту через Пролив Хуана де Фука на 15 000 14C лет BP (18,000 кал лет БП).[17][18] Прибрежные альпийские ледники начали отступать около 19000 человек. кал лет BP [19] в то время как кордильерские льды продолжали наступать в Пьюджет-Лоулендс до 14 000 14C лет назад (16800 кал. Лет назад).[18] Даже во время максимальной протяженности прибрежного льда, не покрытого льдом Refugia сохранились на современных островах, которые питали наземных и морских млекопитающих.[16] Когда произошла дегляциация, рефугиумы расширились, пока побережье не стало свободным ото льда к 15 000 кал. Лет назад.[16] Отступление ледников на Аляскинском полуострове обеспечило доступ из Берингии к тихоокеанскому побережью примерно на 17 000 кал. Лет назад.[20] Ледяной барьер между внутренней Аляской и тихоокеанским побережьем разрушился примерно с 13 500 человек. 14C лет (16 200 кал. Лет) л.[17] Свободный ото льда коридор в глубь Северной Америки открылся между 13 000 и 12 000 кал. Лет назад.[14][15][16] Оледенение в Восточной Сибири во время LGM ограничивалось альпийскими и долинными ледниками в горных хребтах и не блокировало доступ между Сибирью и Берингией.[13]
Климат и биологическая среда
Палеоклимат и растительность Восточной Сибири и Аляски во время оледенения Висконсина были выведены из данных изотопов кислорода высокого разрешения и пыльца стратиграфия.[13][21][22] Перед последним ледниковым максимумом климат в Восточной Сибири колебался между условиями, приближающимися к современным условиям, и более холодными периодами. Теплые циклы до LGM в Арктической Сибири были периодом расцвета мегафауны.[13] Запись изотопов кислорода в ледниковой шапке Гренландии предполагает, что эти циклы после примерно 45 тысяч лет назад продолжались от сотен до одной-двух тысяч лет, а более продолжительные холодные периоды начинались примерно с 32 тысяч лет назад.[13] Запись пыльцы с озера Эликчан, к северу от Охотское море, показывает заметный сдвиг от пыльцы деревьев и кустарников к пыльце трав до 26 тыс. 14C лет до н.э., так как травянистая тундра сменила северный лес и кустарниковую степь, переходящую в LGM.[13] Аналогичная запись о замене пыльцы деревьев / кустарников пыльцой трав по мере приближения LGM была обнаружена у реки Колыма в Арктической Сибири.[22] Оставление северных регионов Сибири из-за быстрого похолодания или отступления охотничьих видов с наступлением LGM было предложено для объяснения отсутствия в этом регионе археозитов, относящихся к LGM.[22][23] Запись пыльцы со стороны Аляски показывает сдвиги между травянисто-кустарниковой тундрой и кустарниковой тундрой до LGM, что указывает на менее драматические эпизоды потепления, чем те, которые позволили колонизировать леса на сибирской стороне. Разнообразны, но не обязательно многочисленны, мегафауны присутствовали в тех средах. Во время LGM доминировали травянистые тундры из-за холодных и засушливых условий.[21]
Прибрежная среда во время последнего ледникового максимума была сложной. Пониженный уровень моря и изостатический выступ, уравновешенный депрессией под Кордильерским ледниковым щитом, обнажил континентальный шельф, образуя прибрежную равнину.[24] В то время как большая часть прибрежной равнины была покрыта предгорных ледников, не покрытых льдом Refugia поддерживающие наземные млекопитающие были идентифицированы на Хайда Гвайи, Остров Принца Уэльского, и внешние острова Александровский архипелаг.[21] Теперь затопленная прибрежная равнина имеет потенциал для новых убежищ.[21] Данные по пыльце указывают на то, что тундровая растительность в основном травянисто-кустарниковая, на не покрытых льдом территориях, с некоторыми бореальными лесами в южной части ареала кордильских льдов.[21] Прибрежная морская среда оставалась продуктивной, о чем свидетельствуют окаменелости ластоногие.[24] Высокопроизводительный ламинария леса на каменистых морских отмелях могли быть приманкой для прибрежной миграции.[25][26] Реконструкция южной береговой линии Берингии также предполагает наличие потенциала для высокопродуктивной прибрежной морской среды.[26]
Изменения окружающей среды во время дегляциации
Данные по пыльце указывают на теплый период, достигающий кульминации между 14 и 11 тысячами. 14C лет BP (17–13 000 кал. Лет BP) с последующим охлаждением между 11–10 000 14C лет BP (13–11,5 тыс. Кал. Лет назад).[24] Прибрежные районы быстро теряли ледниковый покров, когда отступали прибрежные альпийские ледники, а затем отступали части кордильерского льда. Отступление ускорилось из-за повышения уровня моря и плавучести ледников. Было подсчитано, что побережье было полностью свободным ото льда между 16 тыс.[24] и 15к[16] кал лет БП. Литораль морские организмы колонизировали береговые линии, когда вода океана заменила талую ледниковую воду. Замещение травяно-кустарниковой тундры хвойными лесами продолжалось на 12,4 тыс. 14C лет назад (15 тыс. Кал. Лет назад) к северу от Хайда-Гвайи. Эвстатический подъем уровня моря вызвал наводнения, которые усиливались по мере того, как скорость росла.[24]
Внутренние ледяные щиты Кордильера и Лаурентиды отступали медленнее, чем прибрежные ледники. Открытие незамерзающего коридора произошло только через 13–12 тыс. Кал. Лет назад.[14][15][16] В ранней среде незамерзающего коридора преобладали ледниковые вымывания и талая вода, озера, покрытые льдом, и периодические наводнения из-за выхода талой воды, перекрытой льдом.[14] Биологическая продуктивность дегляцируемого ландшафта увеличивалась медленно.[16] Наиболее ранняя возможная жизнеспособность незамерзающего коридора в качестве маршрута миграции людей оценивается в 11,5 тыс. Кал. Лет назад.[16]
Березовые леса продвигались по бывшей травяной тундре в Берингии на 14,3 тыс. 14C лет BP (17 тыс. Кал. Лет назад) в ответ на улучшение климата, что указывает на повышение продуктивности ландшафта.[22]
Анализ биомаркеров и микрофоссилий, сохранившихся в отложениях из озера E5 и озера Бериал на севере Аляски, позволяет предположить, что первые люди сожгли берингийские ландшафты еще 34000 лет назад.[27][28] Авторы этих исследований предполагают, что огонь использовался как средство охоты на мегафауну.
Хронология, причины и источники миграции

Археологическое сообщество в целом согласно с тем, что предки Коренные народы Америки исторических записей поступило в Америку в конце Последний ледниковый максимум (LGM), вскоре после 20 000 лет назад, с подтвержденным археологическим присутствием вскоре после 16 000 лет назад.
Остаются неопределенности относительно точной датировки отдельных участков и выводов, сделанных на популяционная генетика исследования современных коренных американцев. Также остается открытым вопрос, представляла ли эта миграция после LGM первое заселение Америки или же это была более ранняя миграция, предшествующая LGM, которая достигла Южной Америки уже 40 000 лет назад.
Хронология
В начале 21 века модели хронологии миграции делятся на два общих подхода.[29][30]
Первый - это теория краткой хронологии, что первая миграция произошла после последнего ледникового максимума, который пошел на убыль примерно 19000 лет назад,[19] а затем последовали последовательные волны иммигрантов.[31]
Вторая теория - это теория длинной хронологии, который предполагает, что первая группа людей вошла в Америку гораздо раньше, возможно, раньше 40 000 лет назад,[32][33][34] за ней последовала гораздо более поздняя вторая волна иммигрантов.[30][35]
В Хлодвиг Первая теория, которая доминировала в размышлениях об антропологии Нового Света на протяжении большей части 20-го века, была поставлена под сомнение надежной датировкой археозитов в Америке до 13000 лет назад в 2000-х годах.[14][15][16][36][37] Сценарий «короткой хронологии» в свете этого относится к заселению Америки вскоре после 19 000 лет назад, в то время как сценарий «длинной хронологии» допускает присутствие до LGM примерно 40 000 лет назад.
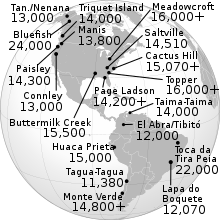
Археозиты в Америке с самыми старыми датами, получившими широкое признание, совместимы с возрастом около 15000 лет. Это включает Комплекс Buttermilk Creek в Техасе,[38] то Meadowcroft Rockshelter сайт в Пенсильвании и Монте-Верде сайт на юге Чили.[37] Археологические свидетельства докловисского народа указывают на Южную Каролину Топпер сайт будучи 16000 лет назад, в то время, когда ледниковый максимум теоретически учитывал более низкие береговые линии.
Часто высказывались предположения, что незамерзающий коридор на территории, которая сейчас Западная Канада, позволил бы миграцию до начала Голоцен, но исследование 2016 года выступило против этого, предполагая, что заселение Северной Америки через такой коридор вряд ли значительно предшествует самым ранним стоянкам Хлодвига. В исследовании делается вывод, что незамерзающий коридор на территории нынешнего Альберта и британская Колумбия «постепенно был захвачен северным лесом, в котором преобладали ели и сосны» и что «люди Хлодвига, вероятно, пришли с юга, а не с севера, возможно, следуя за дикими животными, такими как зубр ".[39][40]Альтернативная гипотеза заселения Америки: прибрежная миграция, что, возможно, было осуществимо вдоль обезледниковой (но теперь затопленной) береговой линии Тихоокеанского Северо-Запада примерно 16000 лет назад.
Доказательства присутствия человека до LGM


Предпоследний ледниковый максимум миграции через Берингию в Америку был предложен для объяснения предполагаемого до-LGM возраста археозитов в Америке, таких как Пещеры Bluefish[33] и Квартиры Old Crow[34] в Территория Юкон, и Приют Медоукрофт Рок в Пенсильвании.[30][35]
В Old Crow Flats были обнаружены кости мамонта, сломанные характерным образом, что указывает на массовую бойню людей. Радиоуглеродные даты на них варьируются от 25 000 до 40 000 лет назад. Также в районе были обнаружены каменные микрочипы, свидетельствующие о производстве инструментов.[42]
Ранее интерпретация следов мясника и геологическая ассоциация костей на участках Bluefish Cave и Old Crow Flats, а также на связанном участке Bonnet Plume ставились под сомнение.[43]
В дополнение к спорным археологическим памятникам, дополнительные доказательства присутствия человека до LGM были обнаружены в записях озерных отложений на севере Аляски. Анализ биомаркеров и микрофоссилий донных отложений из озера E5 и Burial Lake предполагает присутствие человека в восточной Берингии еще 34000 лет назад.[27][28] Эти анализы действительно убедительны, поскольку они подтверждают выводы, сделанные на сайтах Bluefish Cave и Old Crow Flats.
Человеческое присутствие до LGM в Южной Америке частично основывается на хронологии спорных Педра Фурада каменное убежище в Пиауи, Бразилия. Исследование 2003 года датировало свидетельства контролируемого использования огня 40 000 лет назад.[44] Дополнительные доказательства были получены из морфологии Женщина Лузия ископаемое, которое было описано как Австралоид. Эта интерпретация была оспорена в обзоре 2003 года, который пришел к выводу, что рассматриваемые особенности могли также возникнуть в результате генетического дрейфа.[45]
Возраст наиболее ранних положительно идентифицированных артефактов на участке Медоукрофт находится в пределах периода после LGM (13,8–18,5 тыс. кал лет BP ).[36][46]
Камни, описанные как вероятные инструменты, молотковые камни и наковальни, были найдены в Южная Калифорния, на Сайт Cerutti Mastodon, которые связаны с мастодонт скелет, который, по-видимому, был обработан людьми. Скелет мастодонта был датирован радиометрическим анализом торий-230 / уран с использованием моделей диффузионно-адсорбционно-распадного датирования 130,7 ± 9,4 тыс. Лет назад.[47] Человеческие кости не были обнаружены, а заявления об инструментах и обработке костей были описаны как «неправдоподобные».[48]
В Река Яна На участке Рог носорога (справа) заселение человеком восточной Арктической Сибири датируется 27 тыс. 14C лет BP (31,3 тыс. Кал. Лет назад).[49] Эта дата была интерпретирована некоторыми как свидетельство того, что миграция в Берингию была неизбежной, что подтверждает оккупацию Берингии во время LGM.[50][51] Однако дата Яны RHS относится к началу периода охлаждения, который привел к LGM.[13] Но подборка дат археозитов по всей Восточной Сибири предполагает, что период похолодания вызвал отступление людей на юг.[22][23] Каменные свидетельства до LGM в Сибири указывают на оседлый образ жизни, основанный на местных ресурсах, в то время как каменные свидетельства после LGM указывают на более миграционный образ жизни.[23]
Самый старый археозит на аляскинской стороне Берингии датируется 12 тыс. 14C лет BP (14k cal лет BP).[22][52] Возможно, что небольшая популяция основателей вошла в Берингию раньше. Однако археозиты, относящиеся к периоду последнего максимума оледенения на сибирской или аляскинской стороне Берингии, отсутствуют. Анализ биомаркеров и микрофоссилий отложений из озера E5 и Burial Lake на севере Аляски предполагает присутствие человека в восточной Берингии еще 34000 лет назад.[27] Было высказано предположение, что эти осадочные анализы являются единственными возможными извлекаемыми останками людей, живших на Аляске во время последнего ледникового периода.[28]
Оценки геномного возраста
Исследования Американская генетика использовали аналитические методы с высоким разрешением, применяемые к образцам ДНК современных коренных американцев и азиатских популяций, считающихся их исходными популяциями, чтобы реконструировать развитие гаплогруппы ДНК Y-хромосомы человека (yDNA гаплогруппы ) и гаплогруппы митохондриальной ДНК человека (гаплогруппы мтДНК), характерные для индейского населения.[32][50][51] Модели скорости молекулярной эволюции использовались для оценки возраста, в котором линии ДНК коренных американцев отделились от своих родительских линий в Азии, и для определения возраста демографических событий. Одна модель (Tammetal 2007), основанная на гаплотипах мтДНК коренных американцев (рис. 2), предполагает, что миграция в Берингия произошла между 30 тыс. и 25 тыс. кал. лет назад, а миграция в Америку произошла примерно через 10-15 тыс. лет после изоляции небольшого население-основатель.[50] Другая модель (Kitchen et al. 2008) предполагает, что миграция в Берингию произошла приблизительно 36 тыс. Кал. Лет назад, после чего последовали 20 тыс. Лет изоляции в Берингии.[51] Третья модель (Номатто и др. 2009) предполагает, что миграция в Берингию произошла между 40 000 и 30 000 кал. Лет назад, с миграцией до LGM в Америку с последующей изоляцией северной популяции после закрытия незамерзающего коридора.[32] Доказательства примеси австрало-меланезийцев в популяциях Амазонки были обнаружены Скоглундом и Райхом (2016).[53]
Исследование диверсификации гаплогрупп C и D мтДНК из южной Сибири и восточной Азии, соответственно, предполагает, что родительская линия (субгаплогруппа D4h) субгаплогруппы D4h3, линии, обнаруженной среди коренных американцев и ханьцев,[54][55] возникла около 20 тыс. кал. лет назад, что ограничило появление D4h3 после LGM.[56] Возрастные оценки основаны на разнообразии микроспутников Y-хромосомы место происхождения американского Гаплогруппа Q1a3a (Y-ДНК) на уровне от 10 000 до 15 000 кал. лет назад.[57] Большая согласованность моделей скорости молекулярной эволюции ДНК друг с другом и с археологическими данными может быть достигнута за счет использования датированной ископаемой ДНК для калибровки скорости молекулярной эволюции.[54]
Исходные популяции
Среди антропологов существует общее мнение, что исходные популяции для миграции в Америку происходили из района где-то к востоку от Р. Енисей (Дальний Восток России ). Распространенность гаплогрупп A, B, C и D мтДНК среди населения Восточной Азии и коренных американцев была признана давно, наряду с наличием гаплогруппа X.[58] В целом, наибольшая частота четырех гаплогрупп, связанных с коренными американцами, наблюдается в Алтай -Байкал регион южной Сибири.[59] Немного субклады C и D ближе к субкладам коренных американцев встречаются среди монгольского, амурского, японского, корейского и айнского населения.[58][60]
Исследование 2019 года показало, что коренные американцы являются ближайшими родственниками окаменелостей возрастом 10 000 лет, найденных недалеко от Колыма на северо-востоке Сибири.[61]
Геномные модели человека
Развитие геномного анализа с высоким разрешением предоставило возможности для дальнейшего определения субкладов коренных американцев и сужения диапазона азиатских субкладов, которые могут быть родительскими или сестринскими субкладами. Например, широкий географический диапазон гаплогруппы X был интерпретирован как допускающий возможность существования западно-евразийской или даже европейской исходной популяции для коренных американцев, как в Гипотеза солютреана, или предполагая миграцию до Последнего ледникового максимума в Америку.[58] Анализ древнего варианта гаплогруппы X среди аборигенов Алтайского края указывает на общее происхождение с европейским штаммом, а не на происхождение от европейского штамма.[59] Дальнейшее разделение на X субкладов позволило идентифицировать подгаплогруппу X2a, которая считается специфической для коренных американцев.[50][55] С дальнейшим определением субкладов, связанных с индейскими популяциями, требования к выборке азиатских популяций для поиска наиболее близких субкладов становятся более конкретными. Подгаплогруппы D1 и D4h3 были расценены как специфические для коренных американцев на основании их отсутствия среди большой выборки популяций, рассматриваемых как потенциальные потомки исходных популяций, на обширной территории Азии.[50] Среди 3764 образцов Сахалин - ниже Амур регион был представлен 61 Орокс.[50] В другом исследовании субгаплогруппа D1a была идентифицирована среди Ulchis нижнего Приамурья (4 из 87 опрошенных, или 4,6%), а также подгаплогруппа C1a (1 из 87, или 1,1%).[60] Подгаплогруппа C1a считается близкой сестрой подгаплогруппы C1b коренных американцев.[60]
Подгаплогруппа D1a также обнаружена среди древних Jōmon скелеты из Хоккайдо[62] Современный Айны считаются потомками Jōmon.[62] Присутствие субгаплогрупп D1a и C1a в нижнем Приамурье указывает на то, что исходная популяция из этого региона отличается от исходных популяций Алтае-Байкала, где выборка не выявила этих двух конкретных субкладов.[60] Выводы относительно подгаплогруппы D1, указывающие на потенциальные исходные популяции в нижнем течении Амура.[60] и Хоккайдо[62] области отличаются от модели миграции с одним источником.[32][50][51]
Подгаплогруппа D4h3 идентифицирована среди Хань китайский.[54][55] Подгаплогруппа D4h3 из Китая не имеет такого же географического значения, как подгаплотип D1a из Амур-Хоккайдо, поэтому ее значение для исходных моделей является более спекулятивным. Его родительская линия, субгаплотип D4h, как полагают, возникла в Восточной Азии, а не в Сибири, примерно за 20 тыс. Кал. Лет назад.[56] Подгаплогруппа D4h2, родственная клада D4h3, также была обнаружена среди скелетов Дзёмон с Хоккайдо.[63] D4h3 имеет прибрежный след в Северной и Южной Америке.[55]
Контраст между генетическими профилями скелетов Хоккайдо Дзёмон и современных айнов иллюстрирует еще одну неопределенность в исходных моделях, полученных из современных образцов ДНК:[62]
Однако, вероятно, из-за небольшого размера выборки или близкого кровного родства между членами сайта, частота гаплогрупп в скелетах Фунадомари сильно отличалась от любых современных популяций, включая Хоккайдо айнов, которые считались прямым потомком Хоккайдо. Jōmon люди.
Потомки исходных популяций с наиболее близким родством с генетическим профилем с того времени, когда произошла дифференциация, не очевидны. Можно ожидать, что модели исходной популяции станут более надежными по мере того, как будет собрано больше результатов, наследие современных прокси-кандидатов станет более понятным, а ископаемые ДНК в интересующих регионах будут найдены и рассмотрены.
Геномика HTLV-1
Лимфотрофный вирус Т-клеток человека 1 (HTLV-1 ) представляет собой вирус, передающийся при обмене жидкостями организма и от матери к ребенку с грудным молоком. Передача от матери ребенку имитирует наследственный признак, хотя такая передача от материнского носителя составляет менее 100%.[64] Геном вируса HTLV был нанесен на карту, что позволило идентифицировать четыре основных штамма и проанализировать их древность посредством мутаций. Самые высокие географические концентрации штамма HLTV-1 находятся в Африке к югу от Сахары и Японии.[65] В Японии это наибольшая концентрация на Кюсю.[65] Он также присутствует среди потомков африканцев и коренного населения в Карибском регионе и Южной Америке.[65] Редко в Центральной Америке и Северной Америке.[65] Считается, что его распространение в Америке связано с ввозом с работорговлей.[66]
Айны разработали антитела к HTLV-1, что указывает на его эндемичность для айнов и его древность в Японии.[67] Подтип «А» был определен и идентифицирован среди японцев (включая айнов), а также среди карибских и южноамериканских изолятов.[68] Подтип «B» был идентифицирован в Японии и Индии.[68] В 1995 году у коренных американцев в прибрежной части Британской Колумбии были обнаружены подтипы A и B.[69] Сообщалось, что образцы костного мозга от мумии Анд, возрастом около 1500 лет, показали наличие подтипа А.[70] Открытие вызвало разногласия с утверждением, что образец ДНК был недостаточно полным для заключения и что результат отражал современное загрязнение.[71] Однако повторный анализ показал, что последовательности ДНК согласуются с "космополитической кладой" (подтип А), но не определенно от нее.[71] Присутствие подтипов A и B в Северной и Южной Америке наводит на мысль о коренном американском исходном населении, связанном с предками айнов, джомонами.
Физическая антропология
Палеоамериканские скелеты в Америке, такие как Kennewick Man (Штата Вашингтон), Скелет хойи негр (Юкатан), Женщина Лузия и другие черепа из Лагоа Санта сайт (Бразилия), Буль Женщина (Айдахо), Пеньон Женщина III,[72] два черепа со стоянки Тлапакоя (Мехико),[72] и 33 черепа из Нижней Калифорнии[73] проявили черепно-лицевые черты, отличные от большинства современных коренных американцев, что привело физических антропологов к мнению, что некоторые палеоамериканцы имели Австралоид а не сибирского происхождения.[74] Самым основным измеряемым отличительным признаком является долихоцефалия черепа. Некоторые современные изоляты, такие как Перикуэс Нижней Калифорнии и Огнеземельцы Огненной Земли демонстрируют ту же морфологическую черту.[73] Другие антропологи отстаивают альтернативную гипотезу о том, что эволюция первоначального берингийца фенотип дали начало отчетливой морфологии, которая была сходной во всех известных палеоамериканских черепах, с последующей конвергенцией к современному индейскому фенотипу.[75][76] Решение проблемы зависит от идентификации беринговского фенотипа среди палеоамериканских черепов или свидетельства генетической кластеризации среди примеров австралоидного фенотипа.
Отчет опубликован в Американский журнал физической антропологии в январе 2015 г. был проведен обзор черепно-лицевых вариаций с акцентом на различия между ранними и поздними коренными американцами и их объяснения, основанные либо на морфологии черепа, либо на молекулярной генетике. Аргументы, основанные на молекулярной генетике, в основном, по мнению авторов, допускают единственную миграцию из Азии с вероятной паузой в Беренгии, а также более поздний двунаправленный поток генов. Исследования, посвященные черепно-лицевой морфологии, утверждали, что палеоамериканские останки «были описаны как гораздо более близкие к популяциям африканцев и австрало-меланезийцев, чем к современной серии коренных американцев», что предполагает два проникновения в Америку, ранний из которых произошел до самобытного восточноазиатского развитая морфология (именуемая в документе «Модель двух компонентов». Третья модель, модель «рекуррентного потока генов» [RGF], пытается примирить эти две модели, утверждая, что циркумарктический поток генов после первоначальной миграции может объяснить морфологические В нем дается особая переоценка первоначального отчета о скелете Hoya Negro, который поддерживал модель RGF, авторы не согласились с первоначальным выводом, который предполагал, что форма черепа не соответствует форме современных коренных американцев, утверждая, что «череп падает в субрегион морфопространства, занятый как палеоамериканцами, так и некоторыми современными коренными американцами ».[77]
Стволовые точки
Стержневые острия - это каменная технология, отличная от берингийской и кловисской. Распространены они от прибрежных районов Восточной Азии до тихоокеанского побережья Южной Америки.[25] Возникновение стволовых точек прослеживается до Кореи в период верхнего палеолита.[78] Происхождение и распространение выловленных точек интерпретировалось как культурный маркер, связанный с исходной популяцией из прибрежных районов Восточной Азии.[25]
Маршруты миграции
Внутренний маршрут

Исторически теории о миграции в Америку были сосредоточены на миграции из Берингии через внутренние районы Северной Америки. Обнаружение артефактов, связанных с фауной плейстоцена, недалеко от Кловис, Нью-Мексико в начале 1930-х годов потребовалось продлить сроки заселения Северной Америки до периода, когда ледники еще были обширными. Это привело к гипотезе о маршруте миграции между ледниковыми щитами Лаврентида и Кордильера, чтобы объяснить раннее поселение. На участке Хлодвига использовалась каменная технология, характеризующаяся наконечниками для копий с углублением или канавкой, где наконечник прикреплялся к древку. Каменный комплекс, характеризующийся Clovis Point Впоследствии технология была обнаружена на большей части территории Северной и Южной Америки. Связь сложной технологии Хлодвига с остатками фауны позднего плейстоцена привела к теории о том, что это ознаменовало прибытие охотников на крупную дичь, которые мигрировали из Берингии, а затем рассеялись по всей Америке, иначе известная как теория Хлодвига.
Недавнее радиоуглеродное датирование стоянок Хлодвига дало возраст от 11,1 тыс. До 10,7 тыс. 14C лет назад (13–12,6 тыс. Кал. Лет назад), что несколько позже дат, полученных с помощью более старых методов.[79] Переоценка более ранних радиоуглеродных дат привела к выводу, что не менее 11 из 22 стоянок Хлодвига с радиоуглеродными датами являются «проблематичными» и не должны приниматься во внимание, включая тип сайта в Кловисе, Нью-Мексико. Численное датирование стоянок Хлодвига позволило сравнить даты Хлодвига с датами других археозитов по всей Америке, а также с датами открытия незамерзающего коридора. И то и другое ставит под сомнение теорию Хлодвига Первого. Место Монте-Верде на юге Чили датировано 14,8 тыс. Кал. Лет назад.[37] На участке пещеры Пейсли в восточном Орегоне был обнаружен 14Дата C 12,4 тыс. Лет (14,5 тыс. Кал. Лет) лет назад, на копролите с ДНК человека и 14Даты C составляют 11,3–11 тыс. (13,2–12,9 тыс. Кал. Лет) л.н. на горизонтах, содержащих точки западного ствола.[80] Горизонты артефактов с некловисскими каменными комплексами и докловисским возрастом встречаются в восточной части Северной Америки, хотя максимальные возрасты, как правило, плохо ограничиваются.[36][46]
Геологические данные о сроках создания незамерзающего коридора также ставят под сомнение представление о том, что Хлодвиг и дохловисская оккупация Америки человеком была результатом миграции по этому маршруту после Последний ледниковый максимум. Закрытие коридора перед LGM может приблизиться к 30 тыс. Кал. Лет назад, а оценки отступления льда от коридора находятся в диапазоне от 12 до 13 тыс. Кал. Лет назад.[14][15][16] Жизнеспособность коридора как пути миграции людей была оценена в 11,5 тыс. Кал. Лет назад, что позже, чем период стоянок Хлодвиг и докловис.[16] Датированные археозиты Хлодвига указывают на распространение культуры Хлодвига с юга на север.[14]
Предпоследний ледниковый период Максимальная миграция во внутренние районы была предложена для объяснения докловисского возраста археозитов в Америке.[30][35] хотя места до Хлодвига, такие как Meadowcroft Rock Shelter,[36][46] Монте-Верде,[37] и пещера Пейсли не подтвердили возраст до LGM.
Предложение о дене-енисейской языковой семье
Отношения между Языки на-дене Северной Америки (например, навахо и апачи), а Енисейские языки Сибири была впервые предложена еще в 1923 году и получила дальнейшее развитие другими. Подробное исследование было проведено Эдвард Вайда и опубликовано в 2010 году.[81] Эта теория получила поддержку многих лингвистов. Это также подтвердили археологические и генетические исследования.
В Арктическая традиция малых инструментов Аляски и канадской Арктики, возможно, возникли в Восточной Сибири около 5000 лет назад. Это связано с древним Палеоэскимосский народов Арктики, культура которых сформировалась к 2500 г. до н. э.
Источником традиции Arctic Small Tool, возможно, был Сялах -Белкачи-Ымяхтахская культура толщу Восточной Сибири, датируемую 6500 - 2800 кал.[82]
Внутренний маршрут соответствует распространению языковой группы на-дене.[81] и субгаплогруппа X2a в Америку после самой ранней палеоамериканской миграции.[55]
Тем не менее, некоторые ученые предполагают, что предки западных североамериканцев, говорящих на языках на-дене, совершили прибрежную миграцию на лодке.[83]
Тихоокеанский прибрежный маршрут
«Теория прибрежной миграции» Тихоокеанского региона предполагает, что люди сначала достигли Америки посредством водных путешествий, следуя по береговой линии из северо-восточной Азии в Америку, что было первоначально предложено в 1979 году Кнутом Фладмарком в качестве альтернативы гипотезе о незамерзающем коридоре.[84]
Эта модель поможет объяснить быстрое распространение на прибрежные участки, чрезвычайно удаленные от региона Берингова пролива, включая такие участки, как Монте-Верде на юге Чили и Тайма-Тайма на западе Венесуэла. «Гипотеза морской миграции» - это вариант прибрежной миграции, который предполагает использование лодок. Предлагаемое использование лодок добавляет некоторую гибкость в хронологию прибрежной миграции, поскольку непрерывное свободное ото льда побережье (16-15 тыс. Кал. Лет назад) больше не потребуется, поскольку мигранты поселились бы в прибрежных убежищах во время дегляциации побережья. . Прибрежная популяция источника в Восточной Азии является неотъемлемой частью гипотезы о морской миграции.[25][26]
Статья 2007 года в Журнал островной и прибрежной археологии предложила «гипотезу о шоссе ламинарии», вариант прибрежной миграции, основанный на эксплуатации ламинария леса вдоль большей части Тихоокеанского побережья от Японии до Берингии, Тихоокеанского Северо-Запада и Калифорнии, а также до Андского побережья Южной Америки. После того, как береговые линии Аляски и Британской Колумбии потеряли ледяной покров около 16000 лет назад, эти места обитания водорослей (вместе с эстуариями, мангровыми зарослями и коралловыми рифами) стали бы экологически гомогенным миграционным коридором, полностью на уровне моря и практически беспрепятственным. Анализ ДНК растений и животных показывает, что прибрежный маршрут был возможен.[85][86]
Митохондриальная субгаплогруппа D4h3a, редкий субклад D4h3 Встречающаяся вдоль западного побережья Америки, была идентифицирована как клад, связанная с прибрежной миграцией.[55]Эта гаплогруппа была обнаружена в скелете, названном Анзик-1, найденный в Монтане в тесной связи с несколькими артефактами Хлодвига, датируемыми 12500 лет назад.[87]
Проблемы с оценкой моделей прибрежной миграции
Модели прибрежной миграции предлагают другой взгляд на миграцию в Новый Свет, но они не лишены своих проблем. Одна из таких проблем заключается в том, что уровень мирового океана поднялся более чем на 120 метров (390 футов).[88] с конца последнего ледникового периода, и это затопило древние береговые линии, по которым морские люди следовали бы в Америку. Найти участки, связанные с ранними прибрежными миграциями, чрезвычайно сложно, а систематические раскопки любых участков, обнаруженных в более глубоких водах, сложно и дорого. Стратегии поиска мест наиболее ранних миграций включают выявление потенциальных участков на затопленных палеошорьевых линиях, поиск участков в областях, поднятых либо тектоникой, либо изостатическим отскоком, а также поиск прибрежных участков в районах, которые могли привлекать прибрежных мигрантов.[25][89] С другой стороны, есть свидетельства морских технологий, найденных на холмах Нормандские острова Калифорнии, около 10 000 г. до н. э.[90] Если была ранняя прибрежная миграция до Хлодвига, всегда есть возможность «неудавшейся колонизации». Другая проблема, которая возникает, - это отсутствие убедительных доказательств теории «длинной хронологии». Ни один из сайтов еще не дал последовательной хронологии старше 12 500 радиоуглеродных лет (~ 14 500 календарных лет).[нужна цитата ], но исследования, связанные с возможностью ранних прибрежных миграций, в Южной Америке были ограничены.
Смотрите также
- Ранние миграции людей
- Список первых населенных пунктов
- Генетическая история коренных народов Америки
- Доколумбовые теории трансокеанских контактов
Рекомендации
- ^ Прингл, Хизер (8 марта 2017 г.). «Что происходит, когда археолог бросает вызов общепринятому научному мышлению?». Смитсоновский институт.
- ^ Фаган, Брайан М. и Дуррани, Надя (2016). Предыстория мира: краткое введение. Рутледж. п. 124. ISBN 978-1-317-34244-1.
- ^ а б Гебель, Тед; Waters, Michael R .; О'Рурк, Деннис Х. (2008). «Расселение современного человека в Америке в позднем плейстоцене» (PDF). Наука. 319 (5869): 1497–1502. Bibcode:2008Sci ... 319.1497G. CiteSeerX 10.1.1.398.9315. Дои:10.1126 / science.1153569. PMID 18339930. S2CID 36149744. Архивировано 02 января 2014 года.. Получено 2010-02-05.CS1 maint: BOT: статус исходного URL-адреса неизвестен (связь)
- ^ Циммер, Карл (3 января 2018 г.). «В костях погребенного ребенка - признаки массовой миграции людей в Америку». Нью-Йорк Таймс. Получено 3 января, 2018.
- ^ Морено-Маяр, СП; Поттер, BA; Виннер, L; и другие. (2018). «Конечный плейстоценовый геном Аляски показывает первую основную популяцию коренных американцев» (PDF). Природа. 553 (7687): 203–207. Bibcode:2018Натура.553..203M. Дои:10.1038 / природа25173. PMID 29323294. S2CID 4454580.
- ^ Вегспак, Николь (2012). "Ранние палеоиндейцы, от колонизации до фолсома". В Тимоти Р. Паукетат (ред.). Оксфордский справочник по археологии Северной Америки. Издательство Оксфордского университета. С. 86–95. ISBN 978-0-19-538011-8.
- ^ Корнфельд, Марсель и Политис, Густаво Г. (2014). «В Америку: первые охотники-собиратели на пустом континенте». В Вики Каммингс; Питер Джордан и Марек Звелебил (ред.). Оксфордский справочник по археологии и антропологии охотников-собирателей. Издательство Оксфордского университета. п. 406. ISBN 978-0-19-102526-6.
- ^ Эш, Патрисия Дж. И Робинсон, Дэвид Дж. (2011). Появление людей: исследование хронологии эволюции. Джон Уайли и сыновья. п. 289. ISBN 978-1-119-96424-7.
- ^ Робертс, Алиса (2010). Невероятное человеческое путешествие. A&C Black. С. 101–103. ISBN 978-1-4088-1091-0.
- ^ Ardelean, Ciprian F .; Бесерра-Вальдивия, Лорена; Педерсен, Миккель Винтер; Швеннингер, Жан-Люк; Oviatt, Charles G .; Масиас-Кинтеро, Хуан I .; Арройо-Кабралес, Хоакин; Сикора, Мартин; Окампо-Диас, Ям Зул Э .; Рубио-Сиснерос, Игорь I .; Уотлинг, Дженнифер Дж .; Де Медейрос, Ванда Б .; Де Оливейра, Пауло Э .; Барба-Пингарон, Луис; Ортис-Бутрон, Агустин; Бланкас-Васкес, Хорхе; Ривера-Гонсалес, Иран; Солис-Росалес, Корина; Родригес-Сеха, Мария; Gandy, Devlin A .; Наварро-Гутьеррес, Замара; де ла Роса-Диас, Хесус Дж .; Уэрта-Арельяно, Владимир; Маррокин-Фернандес, Марко Б .; Мартинес-Риохас, Л. Мартин; Лопес-Хименес, Алехандро; Хайэм, Томас; Виллерслев, Эске (2020). «Свидетельства человеческой оккупации Мексики в районе последнего ледникового максимума». Природа. 584 (7819): 87–92. Дои:10.1038 / s41586-020-2509-0. PMID 32699412. S2CID 220697089.
- ^ Бесерра-Вальдивия, Лорена; Хайэм, Томас (2020). «Время и эффект первых прибытий людей в Северную Америку». Природа. 584 (7819): 93–97. Дои:10.1038 / s41586-020-2491-6. PMID 32699413. S2CID 220715918.
- ^ Грун, Рут (22 июля 2020 г.). «Появляются доказательства того, что заселение Америки началось более 20 000 лет назад». Природа. 584 (7819): 47–48. Дои:10.1038 / d41586-020-02137-3. PMID 32699366. S2CID 220717778. Получено 24 июля 2020.
- ^ а б c d е ж грамм час Бригам-Гретта, Джули; Ложкин, Анатолий В .; Андерсон, Патриция М. и Глушкова, Ольга Ю. (2004). «Палеоэкологические условия Западной Берингии до последнего ледникового максимума». В D.B. Мэдсен (ред.). Вход в Америку: Северо-Восточная Азия и Берингия до последнего ледникового максимума. Университет Юты Пресс. ISBN 978-0-87480-786-8.
- ^ а б c d е ж грамм Джексон, Лайонел Э. младший и Уилсон, Майкл С. (февраль 2004 г.). "Новый взгляд на незамерзающий коридор". Geotimes. Американский геологический институт.
- ^ а б c d е Джексон, Л. Jr .; Phillips, F.M .; Шимамура К. и Литтл Э. К. (1997). «Космогенный 36Cl-датирование поезда Foothills Erratics, Альберта, Канада ". Геология. 25 (3): 195–198. Bibcode:1997Гео .... 25..195J. Дои:10.1130 / 0091-7613 (1997) 025 <0195: ccdotf> 2.3.co; 2.
- ^ а б c d е ж грамм час я j k Мандрик, Кэрол А.С .; Йозенханс, Хайнер; Fedje, Дэрил В. и Мэтьюз, Рольф В. (январь 2001 г.). «Позднечетвертичные палеосреды северо-запада Северной Америки: последствия для внутренних и прибрежных маршрутов миграции». Четвертичные научные обзоры. 20 (1): 301–314. Bibcode:2001QSRv ... 20..301M. Дои:10.1016 / s0277-3791 (00) 00115-3.
- ^ а б Дайк, A.S .; Мур А. и Робертсон Л. (2003). Дегляциация Северной Америки (Отчет). Откройте файл 1574. Геологическая служба Канады. Дои:10.4095/214399.
- ^ а б Бут, Дерек Б .; Троост, Кэти Гетц; Клэг, Джон Дж. И Уайтт, Ричард Б. (2003). "Ледяной покров Кордильеров". Четвертичный период в США. Развитие четвертичных наук. 1. С. 17–43. Дои:10.1016 / S1571-0866 (03) 01002-9. ISBN 978-0-4445-1470-7.
- ^ а б Blaise, B .; Clague, J.J. И Мэтьюз Р.В. (1990). «Время максимального позднего Висконсинского оледенения, западное побережье Канады». Четвертичное исследование. 34 (3): 282–295. Bibcode:1990QuRes..34..282B. Дои:10.1016 / 0033-5894 (90) 90041-я.
- ^ Мисарти, Николь; Финни, Брюс П .; Джордан, Джеймс У .; и другие. (10 августа 2012 г.). «Раннее отступление ледникового комплекса полуострова Аляска и последствия для прибрежных миграций первых американцев». Четвертичные научные обзоры. 48: 1–6. Bibcode:2012QSRv ... 48 .... 1M. Дои:10.1016 / j.quascirev.2012.05.014.
- ^ а б c d е Clague, Джон Дж .; Мэтьюз, Рольф В. и Агер, Томас А. (2004). «Окрестности северо-запада Северной Америки до последнего ледникового максимума». В D.B. Мэдсен (ред.). Вход в Америку: Северо-Восточная Азия и Берингия до последнего ледникового максимума. Университет Юты Пресс. ISBN 978-0-87480-786-8.
- ^ а б c d е ж Васильев, Сергей А .; Кузьмин, Ярослав В .; Орлова, Любовь А., Дементьев, Вячеслав Н. (2002). «Хронология палеолита Сибири на основе радиоуглерода и ее актуальность для заселения Нового Света». Радиоуглерод. 44 (2): 503–530. Дои:10.1017 / s0033822200031878.
- ^ а б c Граф, Келли Э. (2009). «Современная человеческая колонизация мамонтовой степи: взгляд из южно-центральной Сибири» (PDF). В лагерях Марты; Парт Чаухан (ред.). Справочник переходов палеолита. Springer. С. 479–501. Дои:10.1007/978-0-387-76487-0_32. ISBN 978-0-387-76478-8.
- ^ а б c d е Fedje, Дэрил В .; Маки, Квентин; Диксон, Э. Джеймс и Хитон, Тимоти Х. (2004). «Окружающая среда позднего Висконсина и археологические раскопки вдоль северного северо-западного побережья». В D.B. Мэдсен (ред.). Вход в Америку: Северо-Восточная Азия и Берингия до последнего ледникового максимума. Университет Юты Пресс. ISBN 978-0-87480-786-8.
- ^ а б c d е Эрландсон, Джон М. и Брейдж, Тодд Дж. (2011). «Из Азии в Америку на лодке? Палеогеография, палеоэкология и опорные точки северо-западной части Тихого океана». Четвертичный международный. 239 (1–2): 28–37. Bibcode:2011QuInt 239 ... 28E. Дои:10.1016 / j.quaint.2011.02.030.
- ^ а б c Эрландсон, Джон М .; Грэм, Майкл Х .; Bourque, Брюс Дж .; и другие. (2007). «Гипотеза автомагистрали ламинарии: морская экология, теория прибрежной миграции и заселение Америки». Журнал островной и прибрежной археологии. 2 (2): 161–174. Дои:10.1080/15564890701628612. S2CID 140188874.
- ^ а б c Vachula, R.S .; Huang, Y .; Russell, J.M .; и другие. (20 мая 2020 г.). «Осадочные биомаркеры подтверждают влияние человека на экосистемы северной Берингии во время последнего ледникового периода». Борей. 49 (3): 514–525. Дои:10.1111 / бор.12449.
- ^ а б c Vachula, R.S .; Huang, Y .; Longo, W. M .; и другие. (13 декабря 2018 г.). «Свидетельства людей ледникового периода в восточной Берингии предполагают раннюю миграцию в Северную Америку». Четвертичные научные обзоры. 205: 35–44. Дои:10.1016 / j.quascirev.2018.12.003.
- ^ Белый, Филип М. (2006). Хронология американских индейцев: хронология американской мозаики. Гринвуд. п. 1. ISBN 978-0-313-33820-5.
- ^ а б c d Уэллс, Спенсер и Рид, Марк (2002). Путешествие человека - генетическая одиссея. Случайный дом. С. 138–140. ISBN 978-0-8129-7146-0.
- ^ Ловгрен, Стефан (13 марта 2008 г.). «Согласно исследованию, Америка была заселена 15 000 лет назад». Национальная география.
- ^ а б c d Бонатто, Сандро Л. и Сальцано, Франциско М. (1997). «Единичная и ранняя миграция населения Америки, подтвержденная данными о последовательности митохондриальной ДНК». Труды Национальной академии наук. 94 (5): 1866–1871. Bibcode:1997PNAS ... 94.1866B. Дои:10.1073 / пнас.94.5.1866. ЧВК 20009. PMID 9050871.
- ^ а б Cinq-Mars, J. (1979). "Пещера Блюфиш 1: Пещерное месторождение Восточного Берингии позднего плейстоцена в Северном Юконе". Канадский журнал археологии (3): 1–32. JSTOR 41102194.
- ^ а б Бонничсен, Робсон (1978). «Критические аргументы в пользу артефактов плейстоцена из бассейна Old Crow, Юкон: предварительное заявление». В Алан Л. Брайан (ред.). Ранний человек в Америке с точки зрения тихоокеанского региона. Случайные статьи № 1. Эдмонтон: Международный отдел антропологии археологических исследований, Университет Альберты. С. 102–118. ISBN 9780888649997.
- ^ а б c Оппенгеймер, Стивен. «Путешествие человечества». Фонд Брэдшоу.
- ^ а б c d Goodyear, Альберт С. (2005). «Свидетельства древних поселений Хлодвига в восточной части Соединенных Штатов». В Робсоне Бонничсене; и другие. (ред.). Палеоамериканское происхождение: за пределами Хлодвига. Население Америки. Центр изучения первых американцев, Техасский университет A&M. С. 103–112. ISBN 978-1-60344-812-3.
- ^ а б c d Диллехей, Томас (2000). Американское поселение: новая предыстория. Нью-Йорк: Основные книги. ISBN 978-0-465-07669-7.
- ^ Каплан, Сара (24 октября 2018 г.). «Самые старые наконечники копий континента дают новые подсказки о первых американцах». Вашингтон Пост.
- ^ Pedersen, Mikkel W .; Рутер, Энтони; Швегер, Чарльз; и другие. (10 августа 2016 г.). «Постледниковая жизнеспособность и колонизация в незамерзающем коридоре Северной Америки». Природа. 537 (7618): 45–49. Bibcode:2016Натура.537 ... 45П. Дои:10.1038 / природа19085. PMID 27509852. S2CID 4450936.
- ^ Чанг, Эмили (10 августа 2016 г.). «Популярная теория о том, как люди заселили Северную Америку, не может быть верной, как показывают исследования: свободный ото льда коридор через Альберту, Британская Колумбия, не может использоваться людьми до прибытия людей Хлодвига». CBC Новости. Получено 10 августа, 2016.
- ^ Рисунок 4 из Андрей, Кухня (2008). "Трехэтапная модель колонизации населения Америки". PLOS ONE. 3 (2): e1596. Bibcode:2008PLoSO ... 3,1596 тыс.. Дои:10.1371 / journal.pone.0001596. ЧВК 2223069. PMID 18270583.
- ^ Морлан, Ричард Э. (4 марта 2015 г.). "Старый Вороний Бассейн". Канадская энциклопедия. Historica Canada.
- ^ Брайант, Вон М., младший (1998). "Pre-Clovis". У Гая Гиббона; и другие. (ред.). Археология доисторической коренных народов Америки: энциклопедия. Справочная библиотека гуманитарных наук Гарленд. 1537. С. 682–683. ISBN 978-0-8153-0725-9.
- ^ Santos, G.M; Bird, M.I; Parenti, F .; и другие. (2003). «Пересмотренная хронология самого низкого уровня заселения скального убежища Педра Фурада, Пиауи, Бразилия: плейстоценовое заселение Америки». Четвертичные научные обзоры. 22 (21–22): 2303–2310. Bibcode:2003QSRv ... 22.2303S. Дои:10.1016 / S0277-3791 (03) 00205-1.
- ^ van Vark, G.N .; Куйзенга Д. и Уильямс Ф.Л. (Июнь 2003 г.). «Кенневик и Лузия: уроки европейского верхнего палеолита». Американский журнал физической антропологии. 121 (2): 181–184, обсуждение 185–188. Дои:10.1002 / ajpa.10176. PMID 12740961.
• Фидель, Стюарт Дж. (2004). «Кенневикские глупости:« Новые »теории о заселении Америки». Журнал антропологических исследований. 60 (1): 75–110. Дои:10.1086 / jar.60.1.3631009. JSTOR 3631009. S2CID 163722475.
• González-José, R .; Bortolini, M.C .; Сантос, Ф. И Бонатто, С. (Октябрь 2008 г.). «Население Америки: изменение формы черепа в континентальном масштабе и его интерпретация с междисциплинарной точки зрения». Американский журнал физической антропологии. 137 (2): 175–187. Дои:10.1002 / ajpa.20854. PMID 18481303. S2CID 32748672. - ^ а б c Adovasio, J.M; Донахью, Дж. И Штукенрат, Р. (1990). "Расдиокарбонная хронология Meadowcroft Rockshelter 1975–1990". Американская древность. 55 (2): 348–354. Дои:10.2307/281652. JSTOR 281652.
• Херст, К. Крис (23 октября 2017 г.). "Что означает кал АД?". Thoughtco.com. Получено 30 октября, 2018. - ^ Холен, Стивен Р .; Deméré, Thomas A .; Фишер, Дэниел С .; и другие. (2017). «Археологические раскопки возрастом 130 000 лет в южной Калифорнии, США». Природа. 544 (7651): 479–483. Bibcode:2017Натура.544..479H. Дои:10.1038 / природа22065. PMID 28447646.
- ^ Ринкон, Пол (26 апреля 2017 г.). «Первые американцы заявляют, что разжигают споры». Новости BBC. Получено 30 апреля 2017.
Майкл Р. Уотерс прокомментировал: «Чтобы продемонстрировать такое раннее заселение Америки, необходимо наличие явных каменных артефактов. Однозначных каменных орудий, связанных с костями, нет ... это место, вероятно, просто интересное палеонтологическое место». Крис Стрингер сказал, что «экстраординарные утверждения требуют экстраординарных доказательств - каждый аспект требует самой тщательной проверки», добавив, что «высокие и сконцентрированные силы, должно быть, потребовались, чтобы сломать самые толстые кости мастодонта, а среда осаждения с низкой энергией, по-видимому, не предоставляет очевидной альтернативы людям, использующим с костями найдены тяжелые булыжники.
- ^ Питулько, В.В .; Никольский, П.А .; Гиря Э.Ю .; и другие. (2 января 2004 г.). «Яна РИТ: человек в Арктике до последнего ледникового максимума». Наука. 303 (5654): 52–56. Bibcode:2004Наука ... 303 ... 52С. Дои:10.1126 / science.1085219. ISSN 0036-8075. PMID 14704419. S2CID 206507352.
- ^ а б c d е ж грамм Тамм, Эрика; Кивисилд, Тоомас; Рейдла, Маэре; и другие. (2007). "Берингийский застой и распространение основателей коренных американцев". PLOS ONE. 2 (9): e829. Bibcode:2007PLoSO ... 2..829T. Дои:10.1371 / journal.pone.0000829. ЧВК 1952074. PMID 17786201.
- ^ а б c d Кухня, Андрей; Миямото, Михал М. и Маллиган, Конни Дж. (2008). "Трехэтапная модель колонизации населения Америки". PLOS ONE. 3 (2): e1596. Bibcode:2008PLoSO ... 3,1596 тыс.. Дои:10.1371 / journal.pone.0001596. ЧВК 2223069. PMID 18270583.
- ^ Гебель, Тед и Бувит, Ян (2011). От Енисея до Юкона: интерпретация изменчивости каменных комплексов в позднем плейстоцене / раннем голоцене Берингии. Центр изучения первых американцев, издательство Texas A&M University Press. п. 5. ISBN 978-1-60344-384-5.
- ^ Скоглунд, Понтус и Райх, Дэвид (декабрь 2016 г.). «Геномный взгляд на население Америки» (PDF). Текущие мнения в области генетики и развития. 41: 27–35. Дои:10.1016 / j.gde.2016.06.016. ЧВК 5161672. PMID 27507099.
Недавно мы провели строгую проверку нулевой гипотезы единой популяции - выходцев из Центральной и Южной Америки, используя полногеномные данные различных коренных американцев. Мы обнаружили статистически четкий сигнал, связывающий коренных американцев в районе Амазонки в Бразилии с современными австрало-меланезийцами и жителями Андаманских островов («австралийцами»). В частности, мы обнаружили, что австралийцы имеют значительно больше генетических вариантов с некоторыми популяциями Амазонки, в том числе говорящими на языках тупи, чем с другими коренными американцами. Мы назвали эту предполагаемую древнюю индейскую родословную «Население Y» в честь Ипикуэра, что означает «предок» в языковой семье тупи.
- ^ а б c Кемп, Брайан М .; Malhi, Ripan S .; Макдонаф, Джон; и другие. (2007). "Генетический анализ останков скелетов раннего голоцена с Аляски и его значение для заселения Америки" (PDF). Американский журнал физической антропологии. 132 (4): 605–621. CiteSeerX 10.1.1.576.7832. Дои:10.1002 / ajpa.20543. PMID 17243155.
- ^ а б c d е ж Perego, Ugo A .; Ахилли, Алессандро; Ангерхофер, Норманн; и другие. (2009). «Отличительные палеоиндийские миграционные пути из Берингии, отмеченные двумя редкими гаплогруппами мтДНК». Текущая биология. 19 (1): 1–8. Дои:10.1016 / j.cub.2008.11.058. PMID 19135370. S2CID 9729731.
- ^ а б Деренко, Мирослава; Малярчук, Борис; Гжибовски, Томаш; и другие. (21 декабря 2010 г.). «Происхождение и расселение гаплогрупп C и D митохондриальной ДНК в Северной Азии».. PLOS ONE. 5 (12): e15214. Bibcode:2010PLoSO ... 515214D. Дои:10.1371 / journal.pone.0015214. ЧВК 3006427. PMID 21203537.
- ^ Бортолини, Мария-Катира; Salzano, Francisco M .; Thomas, Mark G .; и другие. (2003). «Свидетельства Y-хромосомы для различных древних демографических историй в Северной и Южной Америке» (PDF). Американский журнал генетики человека. 73 (3): 524–539. Дои:10.1086/377588. ЧВК 1180678. PMID 12900798.
- ^ а б c Шурр, Теодор Г. (май 2000 г.). «Митохондриальная ДНК и заселение Нового Света» (PDF). Американский ученый. 88 (3): 246. Bibcode:2000AmSci..88..246S. Дои:10.1511/2000.3.246.
- ^ а б Захаров, И.А .; Деренко, М.В .; Малярчук Б.А .; и другие. (12 января 2006 г.). «Вариации митохондриальной ДНК в аборигенных популяциях Алтае-Байкальского региона: значение для генетической истории Северной Азии и Америки». Летопись Нью-Йоркской академии наук. 1011 (1): 21–35. Bibcode:2004НЯСА1011 ... 21З. Дои:10.1196 / Анналы.1293.003. PMID 15126280. S2CID 37139929.
- ^ а б c d е Стариковская, Елена Б .; Сукерник, Рем И .; Дербенева, Ольга А .; и другие. (Январь 2005 г.). «Разнообразие митохондриальной ДНК в коренных популяциях южной части Сибири и происхождение гаплогрупп коренных американцев». Анналы генетики человека. 69 (Pt 1): 67–89. Дои:10.1046 / j.1529-8817.2003.00127.x. ЧВК 3905771. PMID 15638829.
- ^ Сикора, Мартин; Питулько, Владимир В .; Sousa, Vitor C .; и другие. (2019). «История населения Северо-Восточной Сибири с плейстоцена» (PDF). Природа. 570 (7760): 182–188. Bibcode:2019Натура.570..182С. Дои:10.1038 / s41586-019-1279-z. PMID 31168093. S2CID 174809069.
- ^ а б c d Адачи, Нобору; Шинода, Кен-ичи; Умецу, Кадзуо и Мацумура, Хирофуми (март 2009 г.). «Анализ митохондриальной ДНК скелетов Дзёмон с участка Фунадомари, Хоккайдо, и его значение для происхождения коренных американцев». Американский журнал физической антропологии. 138 (3): 255–265. Дои:10.1002 / ajpa.20923. PMID 18951391.
- ^ Адачи, Нобору; Шинода, Кен-ичи; Умецу, Кадзуо; и другие. (Ноябрь 2011 г.). «Анализ митохондриальной ДНК скелетов Хоккайдо Дзёмон: остатки архаичных материнских линий на юго-западном краю бывшей Берингии». Американский журнал физической антропологии. 146 (3): 346–360. Дои:10.1002 / ajpa.21561. PMID 21953438.
- ^ Ли, Хун-Чуань; Биггар, Роберт Дж .; Майли, Венделл Дж .; и другие. (2004). «Провирусная нагрузка в грудном молоке и риск передачи от матери ребенку Т-лимфотропного вируса человека типа I». Журнал инфекционных болезней. 190 (7): 1275–1278. Дои:10.1086/423941. PMID 15346338.
- ^ а б c d Verdonck, K .; González, E .; Van Dooren, S .; и другие. (Апрель 2007 г.). «Человеческий Т-лимфотропный вирус 1: последние сведения о древней инфекции». Ланцетные инфекционные болезни. 7 (4): 266–281. Дои:10.1016 / S1473-3099 (07) 70081-6. PMID 17376384.
- ^ Gessain, A .; Галло, Р. И Франкини, Г. (апрель 1992 г.). «Низкая степень генетического дрейфа Т-клеточного лейкоза / вируса лимфомы человека I типа in vivo как средство мониторинга передачи и перемещения вирусов в древних человеческих популяциях». Журнал вирусологии. 66 (4): 2288–2295. Дои:10.1128 / JVI.66.4.2288-2295.1992. ЧВК 289023. PMID 1548762.
- ^ Исида, Такафуми; Ямамото, Кохтаро; Омото, Кейчи; и другие. (Сентябрь 1985 г.). «Распространенность ретровируса человека у коренных японцев: свидетельства возможного древнего происхождения». Журнал инфекции. 11 (2): 153–157. Дои:10.1016 / s0163-4453 (85) 92099-7. PMID 2997332.
- ^ а б Миура, Т .; Фукунага, Т .; Игараси, Т .; и другие. (Февраль 1994). «Филогенетические подтипы Т-лимфотропного вируса человека типа I и их связь с антропологическим фоном». Труды Национальной академии наук Соединенных Штатов Америки. 91 (3): 1124–1127. Bibcode:1994ПНАС ... 91.1124М. Дои:10.1073 / пнас.91.3.1124. ЧВК 521466. PMID 8302841.
- ^ Picard, F.J .; Coulthart, M.B .; Oger, J .; и другие. (Ноябрь 1995 г.). «Человеческий Т-лимфотропный вирус типа 1 у коренных жителей побережья Британской Колумбии: филогенетическое родство и возможное происхождение». Журнал вирусологии. 69 (11): 7248–56. Дои:10.1128 / JVI.69.11.7248-7256.1995. ЧВК 189647. PMID 7474147.
- ^ Ли, Хун-Чуань; Фудзиёси, Тосинобу; Лу, Хонг; и другие. (Декабрь 1999 г.). «Присутствие ДНК провируса лимфотропного вируса типа I древних Т-клеток человека в мумии Анд». Природа Медицина. 5 (12): 1428–1432. Дои:10.1038/71006. PMID 10581088. S2CID 12893136.
- ^ а б Coulthart, Michael B .; Посада, Давид; Крэндалл, Кейт А. И Декабанд, Грегори А. (март 2006 г.). «О филогенетическом размещении последовательностей вируса Т-клеточного лейкоза типа 1, ассоциированных с андской мумией». Инфекция, генетика и эволюция. 6 (2): 91–96. Дои:10.1016 / j.meegid.2005.02.001. ЧВК 1983367. PMID 16503510.
- ^ а б Гонсалеза, Сильвия; Худдарт, Дэвид; Исраде-Алькантара, Изабель; и другие. (30 марта 2015 г.). «Палеоиндийские памятники в бассейне Мексики: данные стратиграфии, тефрохронологии и датирования» (PDF). Четвертичный международный. 363: 4–19. Bibcode:2015QuInt.363 .... 4G. Дои:10.1016 / j.quaint.2014.03.015.
- ^ а б Гонсалес-Хосе, Роландо; Гонсалес-Мартин, Антонио; Эрнандес, Микель; и другие. (4 сентября 2003 г.). «Краниометрические доказательства выживания палеоамериканцев в Нижней Калифорнии». Природа. 425 (6953): 62–65. Bibcode:2003 Натур. 425 ... 62G. Дои:10.1038 / природа01816. PMID 12955139. S2CID 4423359.
- ^ Диллехей, Томас Д. (4 сентября 2003 г.). «По следам первых американцев». Природа. 425 (6953): 23–24. Дои:10.1038 / 425023a. PMID 12955120. S2CID 4421265.
- ^ Фидель, Стюарт Дж. (Весна 2004 г.). «Безумие Кенневика:« новые »теории о заселении Америки». Журнал антропологических исследований. 60 (1): 75–110. Дои:10.1086 / jar.60.1.3631009. JSTOR 3631009. S2CID 163722475.
- ^ Болтовни, Джеймс К .; Kennett, Douglas J .; Асмером, Йеман; и другие. (16 мая 2014 г.). "Скелет человека позднего плейстоцена и связь мтДНК палеоамериканцев и современных коренных американцев" (PDF). Наука. 344 (6185): 750–754. Bibcode:2014Наука ... 344..750C. Дои:10.1126 / science.1252619. PMID 24833392. S2CID 206556297. Архивировано из оригинал (PDF) на 2015-07-13.
- ^ де Азведо, Соледад; Бортолини, Мария Ч .; Bonatto, Sandro L .; и другие. (Январь 2015 г.). «Древние останки и первое заселение Америки: переоценка негритянского черепа Хойо». Американский журнал физической антропологии. 148 (3): 514–521. Дои:10.1002 / ajpa.22801. PMID 26174009.
• Азеведо, Соледад де; Кинто-Санчес, Мирша; Пашетта, Каролина и Гонсалес-Хосе, Роландо (28 февраля 2017 г.). ""Первое человеческое поселение в Новом Свете «Более пристальный взгляд на черепно-лицевую изменчивость и эволюцию групп коренных американцев раннего и позднего голоцена». Четвертичный международный. 431 (часть B): 152–167. Дои:10.1016 / j.quaint.2015.11.012. - ^ Сеонг, Чунтэк (декабрь 2008 г.). «Острие с косой, микролезвия и поздняя палеолитическая охота в Корее». Античность. 82 (318): 871–883. Дои:10.1017 / с0003598x00097647.
- ^ Уотерс, Майкл Р. и Стаффорд, Томас У. (23 февраля 2007 г.). «Новое определение эпохи Хлодвига: последствия для населения Америки». Наука. 315 (5815): 1122–1126. Bibcode:2007Научный ... 315.1122W. Дои:10.1126 / science.1137166. PMID 17322060. S2CID 23205379.
- ^ Jenkins, Dennis L .; Дэвис, Лорен Дж .; Стаффорд, Томас В., младший; и другие. (13 июля 2012 г.). "Западные стволовые точки снарядов эпохи Хлодвига и человеческие копролиты в пещерах Пейсли". Наука. 337 (6091): 223–228. Bibcode:2012Sci ... 337..223J. Дои:10.1126 / science.1218443. PMID 22798611. S2CID 40706795.
- ^ а б Вайда, Эдвард Дж. (18 апреля 2017 г.). "Дене-Енисейский". Оксфордские библиографии онлайн. Дои:10.1093 / OBO / 9780199772810-0064.
- ^ Флегонтов, Павел; Алтынышик, Н. Эзги; Чангмай, Пия; и другие. (13 октября 2017 г.). «Генетическое наследие палеоэскимосов в Северной Америке». bioRxiv. Дои:10.1101/203018. S2CID 90288469.
• Флегонтов, Павел; Алтынышик, Н. Эзги; Чангмай, Пия; и другие. (5 июня 2019 г.). «Палео-эскимосское генетическое происхождение и население Чукотки и Северной Америки» (PDF). Природа. 570 (7760): 236–240. Bibcode:2019Натура.570..236F. Дои:10.1038 / с41586-019-1251-у. ISSN 0028-0836. ЧВК 6942545. PMID 31168094. - ^ Хандверк, Брайан (12 февраля 2010 г.). "Лицо древнего человека, нарисованного из ДНК волос; Геном рисует картину человека из вымершей культуры Гренландии". Новости National Geographic.
- ^ Фладмарк, Кнут Р. (январь 1979 г.). «Маршруты: альтернативные коридоры миграции для древнего человека в Северной Америке». Американская древность. 44 (1): 55–69. Дои:10.2307/279189. JSTOR 279189.
- ^ Каллавей, Юэн (11 августа 2016 г.). «ДНК растений и животных позволяет предположить, что первые американцы выбрали прибрежный путь». Природа. 536 (7615): 138. Bibcode:2016Натура. 536..138C. Дои:10.1038 / 536138a. PMID 27510205.
- ^ Лето, Томас (10 августа 2016 г.). «Люди, возможно, пошли другим путем в Америку, чем думали, что арктический проход не принесет достаточно еды для первых путешествий американцев». Новости науки.
- ^ Расмуссен, Мортен; Анзик, Сара Л .; Waters, Michael R .; и другие. (Февраль 2014). "Геном человека позднего плейстоцена из могильника Кловис в западной Монтане". Природа. 506 (7487): 225–229. Bibcode:2014Натура.506..225R. Дои:10.1038 / природа13025. ЧВК 4878442. PMID 24522598.
- ^ Горниц, Вивиан (январь 2007 г.). «Повышение уровня моря после таяния льда и сегодня». Институт космических исследований Годдарда. НАСА. Получено 23 апреля 2015.
- ^ Хетерингтон, Рене; Барри, Дж. Вон; Маклауд, Роджер и Уилсон, Майкл (февраль 2004 г.). «В поисках затерянной земли». Geotimes.
- ^ Университет Орегона (4 марта 2011 г.). «Калифорнийские острова отказываются от свидетельств раннего мореплавания: многочисленные артефакты, обнаруженные на стоянках позднего плейстоцена на Нормандских островах». ScienceDaily.
Библиография
- Брэдли, Брюс и Стэнфорд, Деннис Дж. (2004). «Североатлантический коридор кромки льда: возможный путь палеолита в Новый Свет». Мировая археология. 36 (4): 459–478. CiteSeerX 10.1.1.694.6801. Дои:10.1080/0043824042000303656. S2CID 161534521.
- Брэдли, Брюс и Стэнфорд, Деннис Дж. (2006). «Связь солютрейцев и Хлодвига: ответ Штраусу, Мельцеру и Гебелю». Мировая археология. 38 (4): 704–714. Дои:10.1080/00438240601022001. JSTOR 40024066. S2CID 162205534.
- Стэнфорд, Деннис Дж .; Брэдли, Брюс (2012). Первые американцы до Хлодвига: происхождение культуры Хлодвига в Америке. Калифорнийский университет Press. ISBN 978-0-520-22783-5.
- Стэнфорд, Деннис Дж. И Брэдли, Брюс А. (2013). По ту сторону Атлантического льда: происхождение культуры Хлодвига в Америке. Калифорнийский университет Press. ISBN 978-0-520-27578-2.
- Диксон, Э. Джеймс (1993). Поиски истоков первых американцев. Университет Нью-Мексико. ISBN 978-0-8263-1406-2.
- Диксон, Э. Джеймс (1999). Кости, лодки и бизоны: археология и первая колонизация западной части Северной Америки. Пресса Университета Нью-Мексико. ISBN 978-0-8263-2138-1.
- Эрландсон, Джон М. (2013). Ранние охотники-собиратели побережья Калифорнии. Springer Science & Business Media. ISBN 978-1-4757-5042-3.
- Эрландсон, Джон М. (2001). «Археология водных адаптаций: парадигмы нового тысячелетия». Журнал археологических исследований. 9 (4): 287–350. Дои:10.1023 / а: 1013062712695. S2CID 11120840.
- Эрландсон, Джон М. (2002). «Анатомически современные люди, морские путешествия и плейстоценовая колонизация Америки». В Нине Г. Яблонски (ред.). Первые американцы: плейстоценовая колонизация Нового Света. Калифорнийская академия наук. С. 59–92. ISBN 978-0-940228-50-4.
- Эрландсон, Джон. М .; Graham, M. H .; Bourque, Брюс Дж .; и другие. (30 октября 2007 г.). "Гипотеза водорослей: морская экология, теория прибрежной миграции и заселение Америки". Журнал островной и прибрежной археологии. 2 (2): 161–174. Дои:10.1080/15564890701628612. S2CID 140188874.
- Эшлеман, Джейсон А .; Малхи, Рипан С. и Гленн Смит, Дэвид (2003). «Исследования митохондриальной ДНК коренных американцев: концепции и заблуждения о предыстории населения Америки». Эволюционная антропология. 12 (1): 7–18. Дои:10.1002 / evan.10048. S2CID 17049337.
- Fedje, Дэрил В. и Кристенсен, Тина (октябрь 1999 г.). "Моделирование палеошорелей и определение прибрежных участков раннего голоцена в Хайда-Гвайи". Американская древность. 64 (4): 635–652. Дои:10.2307/2694209. JSTOR 2694209.CS1 maint: дата и год (связь)
- Гринман, Э. Ф. (февраль 1963 г.). «Верхний палеолит и Новый Свет». Современная антропология. 4 (1): 41–66. Дои:10.1086/200337. JSTOR 2739818. S2CID 144250630.
- Привет, Джоди (25 мая 2005 г.). "О количестве основателей Нового Света: генетический портрет населения Америки". PLOS Биология. 3 (6): e193. Дои:10.1371 / journal.pbio.0030193. ЧВК 1131883. PMID 15898833.
- Яблонски, Нина Г. (2002). Первые американцы: плейстоценовая колонизация Нового Света. Калифорнийская академия наук. ISBN 978-0-940228-50-4.
- Джонс, Питер Н. (2005). Уважение к предкам: культурная принадлежность американских индейцев на американском Западе. Институт Бауу. ISBN 978-0-9721349-2-7.
- Коротаев Андрей; Березкин, Юрий Е .; Боринская, Светлана А .; Давлетшин, Альберт I .; Халтурина, Дарья А. (2017). «Какие гены и мифы принесли в Новый Свет разные волны заселения Америки?». У Леонида Е. Гринина; Андрей В. Коротаев; Юрий Евгеньевич Березкин (ред.). История и математика: экономика, демография, культура и космические цивилизации. С. 9–77. ISBN 978-5-7057-5247-8.
- Лаубер, Патрисия (2003). Кто пришел первым: новые ключи к доисторическим американцам. Детские книги National Geographic Soc. ISBN 978-0-7922-8228-0.
- Мэтсон, Р. Г. и Коупленд, Гэри (2016). Предыстория Северо-Западного побережья. Тейлор и Фрэнсис. ISBN 978-1-315-41739-4.
- Мельцер, Дэвид Дж. (2009). Первые народы в новом мире: колонизация Америки ледникового периода. Калифорнийский университет Press. ISBN 978-0-520-94315-5.
- Сноу, Дин Р. (1996). «Первые американцы и дифференциация культур охотников-собирателей». В Брюсе Г. Триггере; Уилкомб Э. Уошберн (ред.). Кембриджская история коренных народов Америки: Северная Америка. Том 1: Часть 1. Издательство Кембриджского университета. С. 125–199. ISBN 978-0-521-57392-4.
- Уэллс, Спенсер (2002). Путешествие человека: генетическая одиссея. Издательство Принстонского университета. ISBN 0-691-11532-X.
внешняя ссылка
- Палеоиндийская база данных – Университет Теннесси, факультет антропологии.
- «Первые американцы: как и когда были заселены Америки?», земной шар, Январь 2016
- «Когда люди пришли в Америку?» - Смитсоновский журнал Февраль 2013
- Палеоиндийский период – Министерство внутренних дел США, Служба национальных парков
- Шепард Креч III, Палеоиндийцы и отмирание великого плейстоцена – Американская академия искусств и наук, Национальный гуманитарный центр, 2008.
- Путешествие человека: генетическая одиссея (фильм) на YouTube - к Спенсер Уэллс – PBS и National Geographic Channel, 2003 г. - 120 минут, UPC / EAN: 841887001267
| История |  | |
|---|---|---|
| Урегулирование | ||
| Общества | ||
| Связанный |
| |
| Списки | ||
| Хронология |
| |
| ||
| Предыстория |  | ||||
|---|---|---|---|---|---|
| Мифология / Религия | |||||
| Европейский колонизация | |||||
| Современные группы по стране |
| ||||
| похожие темы | |||||
| |||||
