Ras GTPase - Ras GTPase
 | |||||||||
| Идентификаторы | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Символ | Рас | ||||||||
| Pfam | PF00071 | ||||||||
| ИнтерПро | IPR020849 | ||||||||
| PROSITE | PDOC00017 | ||||||||
| SCOP2 | 5п21 / Объем / СУПФАМ | ||||||||
| CDD | cd04138 | ||||||||
| |||||||||
Рас это семейство родственных белков который экспрессируется во всех линиях и органах животных клеток. Все члены семейства белков Ras принадлежат к классу белков, называемых малая ГТФаза, и участвуют в передаче сигналов внутри ячеек (Сотовая связь преобразование сигнала ). Рас - прототипный член Рас надсемейство белков, которые все связаны в трехмерной структуре и регулируют разнообразное поведение клеток.
Когда Ras «включается» входящими сигналами, он впоследствии включает другие белки, которые в конечном итоге включают гены, участвующие в рост клеток, дифференциация и выживание. Мутации в рас гены могут привести к производству постоянно активированных белков Ras. В результате это может вызвать непреднамеренную и сверхактивную передачу сигналов внутри ячейки, даже при отсутствии входящих сигналов.
Поскольку эти сигналы приводят к росту и делению клеток, сверхактивная передача сигналов Ras может в конечном итоге привести к рак.[1] 3 гена Ras у людей (HRas, KRas, и NRas ) являются наиболее распространенными онкогены в человеческом рак; мутации, которые постоянно активируют Ras, обнаруживаются в 20-25% всех опухолей человека и до 90% при некоторых типах рака (например, панкреатический рак ).[2] По этой причине ингибиторы Ras изучаются как средство лечения рака и других заболеваний со сверхэкспрессией Ras.
История
Первые два рас гены HRAS и KRAS, были определены[3] из исследований двух вирусов, вызывающих рак, вируса саркомы Харви и вируса саркомы Кирстен, авторами Эдвард М. Сколник и коллеги из Национального института здоровья (NIH).[4] Эти вирусы были впервые обнаружены у крыс в 1960-х годах Дженнифер Харви.[5] и Вернер Х. Кирстен,[6] соответственно, отсюда и название Рат sаркома.[3] В 1982 году активировал и преобразовал человеческий рас гены были обнаружены в раковых клетках человека Джеффри М. Купер в Гарварде,[7] Мариано Барбакид и Стюарт А. Ааронсон в NIH,[8] Роберт Вайнберг в Массачусетском технологическом институте,[9] и Майкл Виглер в лаборатории Колд-Спринг-Харбор.[10] Третий рас ген был впоследствии обнаружен исследователями в группе Робин Вайс на Институт исследования рака,[11][12] и Майкл Виглер в лаборатории Колд-Спринг-Харбор,[13] названный NRAS, для его первоначальной идентификации в клетках нейробластомы человека.
Три человека рас гены кодируют очень похожие белки, состоящие из цепочек от 188 до 189 аминокислот. Их генные символы HRAS, NRAS и KRAS, последний из которых производит изоформы K-Ras4A и K-Ras4B из альтернативных сращивание.
Структура
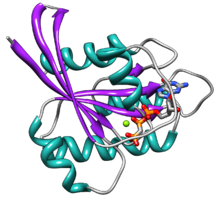
Рас содержит шесть бета-нити и пять альфа спирали.[14]Он состоит из двух доменов: G домена 166 аминокислоты (около 20 кДа), который связывает нуклеотиды гуанозина, и С-концевую нацеливающую область мембраны (CAAX-COOH, также известную как Коробка CAAX ), который модифицирован липидами фарнезилтрансфераза, RCE1, и ICMT.
Домен G содержит пять мотивов G, которые напрямую связываются с GDP / GTP. Мотив G1 или P-петля связывает бета-фосфат GDP и GTP. Мотив G2, также называемый Switch I, содержит треонин35, который связывает концевой фосфат. (γ-фосфат) GTP и двухвалентный ион магния связываются в активном центре. Мотив G3, также называемый Switch II, имеет мотив DXXGQ. D представляет собой аспартат57, который специфичен для связывания гуанина по сравнению с аденином, и Q представляет собой глутамин61, важный остаток, который активирует каталитическую молекулу воды для гидролиза GTP до GDP. Мотив G4 содержит мотив LVGNKxDL и обеспечивает специфическое взаимодействие с гуанином. Мотив G5 содержит консенсусную последовательность SAK. А представляет собой аланин146, который обеспечивает специфичность к гуанину, а не к аденину.
Два мотива переключения, G2 и G3, являются основными частями белка, которые перемещаются при активации GTP. Это конформационное изменение за счет двух мотивов переключения - это то, что обеспечивает базовую функциональность в качестве белка переключения молекул. Это связанное с GTP состояние Ras является состоянием «включено», а состояние привязки к GDP - состоянием «выключено».
Рас также связывает ион магния который помогает координировать связывание нуклеотидов.
Функция
Белки Ras функционируют как бинарные молекулярные переключатели, которые контролируют внутриклеточные сигнальные сети. Ras-регулируемый сигнальные пути контролировать такие процессы, как актин цитоскелет целостность, распространение клеток, дифференциация клеток, клеточная адгезия, апоптоз, и миграция клеток. Ras- и Ras-родственные белки часто не регулируются при раке, что приводит к усилению инвазии и метастаз, и уменьшил апоптоз.
Эта секция нуждается в расширении. Вы можете помочь добавляя к этому. (Апрель 2009 г.) |
Рас активирует несколько путей, из которых каскад митоген-активированных протеинов (MAP) киназ был хорошо изучен. Этот каскад передает сигналы вниз по потоку и приводит к транскрипция генов, участвующих в росте и делении клеток.[15] Другой сигнальный путь, активируемый Ras, - это Путь PI3K / AKT / mTOR, который стимулирует синтез белка и рост клеток, а также подавляет апоптоз.
Активация и деактивация
Рас - это гуанозин -нуклеотид -связывающий белок. В частности, это односубъединичный малая ГТФаза, который по структуре связан с Gα подразделение гетеротримерные G-белки (большие GTPases). G-белки действуют как бинарные сигнальные переключатели с состояниями «включено» и «выключено». В "выключенном" состоянии он связан с нуклеотидом. гуанозиндифосфат (ВВП), в то время как в состоянии "включено", Ras обязан гуанозинтрифосфат (GTP), который имеет дополнительный фосфат группа по сравнению с ВВП. Этот дополнительный фосфат удерживает две области переключателя в конфигурации «нагруженная пружина» (в частности, Thr-35 и Gly-60). При отпускании области переключателя расслабляются, что вызывает конформационное изменение в неактивное состояние. Следовательно, активация и дезактивация Ras и других малых G-белков контролируется циклическим переключением между активной GTP-связанной и неактивной GDP-связанной формами.
Процесс обмена связанного нуклеотида облегчается за счет факторы обмена гуаниновых нуклеотидов (ГЭФ) и Белки, активирующие ГТФазу (Пробелы). Согласно своей классификации, Ras обладает внутренним GTPase активность, а это значит, что белок по собственному желанию гидролизовать связанная молекула GTP в GDP. Однако этот процесс слишком медленный для эффективного функционирования, и, следовательно, GAP для Ras, RasGAP, может связываться и стабилизировать каталитический аппарат Ras, поставляя дополнительные каталитические остатки ("аргинин палец ") таким образом, что молекула воды оптимально расположена для нуклеофильный атака на гамма-фосфат ГТФ. Высвобождается неорганический фосфат, и теперь молекула Ras связана с GDP. Поскольку форма, связанная с GDP, «выключена» или «неактивна» для передачи сигналов, белок, активирующий GTPase инактивирует Ras за счет активации его активности GTPase. Таким образом, GAP ускоряют Ras инактивация.
ГЭФ катализируют реакцию «тяни и толкай», которая высвобождает ВВП из Ras. Они вставляются близко к P-образной петле и магнию катион сайт связывания и ингибировать их взаимодействие с гамма-фосфатом анион. Кислотные (отрицательные) остатки в переключателе II «отталкивают» лизин в P-петле от GDP, что «отталкивает» переключатель I от гуанина. Контакты, удерживающие GDP на месте, разрываются, и он попадает в цитоплазму. Поскольку внутриклеточный GTP больше по сравнению с GDP (примерно в 10 раз больше)[15] GTP преимущественно повторно входит в карман связывания нуклеотидов Ras и перезагружает пружину. Таким образом, ГЭФ содействуют Ras активация.[14] Хорошо известные ГЭФ включают Сын Семерых (СОС) и cdc25 которые включают RasGEF домен.
Баланс между активностью GEF и GAP определяет статус гуаниновых нуклеотидов Ras, тем самым регулируя активность Ras.
В GTP-связанной конформации Ras обладает высоким сродством к многочисленным эффекторы которые позволяют ему выполнять свои функции. Они включают PI3K. Другие небольшие GTPases могут связывать адаптеры, такие как арфаптин или системы вторичного обмена сообщениями, такие как аденилилциклаза. Связывающий домен Ras обнаружен во многих эффекторах и неизменно связывается с одним из переключающих участков, поскольку они изменяют конформацию между активной и неактивной формами. Однако они также могут связываться с остальной поверхностью белка.
Существуют и другие белки, которые могут изменять активность белков семейства Ras. Одним из примеров является GDI (ингибитор диссоциации GDP). Они работают, замедляя обмен GDP на GTP, тем самым продлевая неактивное состояние членов семьи Ras. Могут существовать и другие белки, которые увеличивают этот цикл.
Присоединение к мембране
Рас прикреплен к клеточная мембрана благодаря пренилирование и пальмитоилирование (HRAS и NRAS ) или сочетание пренилирование и многоосновная последовательность, смежная с сайтом пренилирования (KRAS ). C-терминал Коробка CaaX Ras сначала фарнезилируется по остатку Cys в цитозоле, позволяя Ras свободно вставляться в мембрану эндоплазматический ретикулум и другие клеточные мембраны. Затем трипептид (aaX) отщепляется от С-конца специфической пренил-протеин-специфической эндопротеазой, и новый С-конец метилируется с помощью метилтрансфераза. На этом обработка KRas завершена. Динамические электростатические взаимодействия между положительно заряженной основной последовательностью и отрицательными зарядами на внутренней створке плазматическая мембрана объясняют его преимущественную локализацию на поверхности клетки в стационарном состоянии. NRAS и HRAS далее обрабатываются на поверхности аппарат Гольджи от пальмитоилирование одного или двух остатков Cys соответственно, примыкающих к Коробка CaaX. Таким образом, белки устойчиво закрепляются на мембране (липидные рафты) и транспортируются к плазматическая мембрана на пузырьки из секреторный путь. Депальмитоилирование ацилпротеиновые тиоэстеразы в конечном итоге высвобождает белки из мембраны, позволяя им вступить в другой цикл пальмитоилирования и депальмитоилирования.[16] Считается, что этот цикл предотвращает утечку NRAS и HRAS к другим мембранам с течением времени и для поддержания их стационарной локализации вдоль аппарат Гольджи, секреторный путь, плазматическая мембрана и взаимосвязанные эндоцитоз путь.
Члены
Наиболее заметными с клинической точки зрения представителями подсемейства Ras являются: HRAS, KRAS и NRAS, в основном из-за того, что он причастен ко многим типам рака.[17]
Однако есть много других членов этого подсемейства:[18]DIRAS1; DIRAS2; DIRAS3; ERAS; GEM; MRAS; NKIRAS1; NKIRAS2; NRAS; РАЛА; RALB; RAP1A; RAP1B; RAP2A; RAP2B; RAP2C; RASD1; RASD2; RASL10A; RASL10B; RASL11A; RASL11B; RASL12; REM1; REM2; RERG; RERGL; RRAD; RRAS; RRAS2
Рас в раке
Мутации в семье Рас протоонкогены (включающие H-Ras, N-Ras и K-Ras) очень распространены и обнаруживаются в 20-30% всех опухолей человека.[17] Разумно предположить, что фармакологический подход, ограничивающий активность Ras, может представлять собой возможный метод подавления определенных типов рака. Точечные мутации Ras - наиболее частая аномалия протоонкогенов человека.[19]Ингибитор РАС транс-фарнезилтиосалициловая кислота (ФТС, Салирасиб ) проявляет глубокие антионкогенные эффекты во многих линиях раковых клеток.[20][21]
Несоответствующая активация
Было показано, что неправильная активация гена играет ключевую роль в неправильной передаче сигнала, пролиферации и злокачественной трансформации.[15]
Такой эффект могут иметь мутации в ряде различных генов, а также в самом RAS. Онкогены такие как p210BCR-ABL или рецептор роста erbB находятся выше Ras, поэтому, если они конститутивно активированы, их сигналы будут передаваться через Ras.
В ген-супрессор опухоли NF1 кодирует Ras-GAP - его мутацию в нейрофиброматоз будет означать, что Ras с меньшей вероятностью будет деактивирован. Ras также может быть усилен, хотя в опухолях это случается нечасто.
Наконец, онкогены Ras могут активироваться точечными мутациями, так что GTPase реакция больше не может стимулироваться GAP - это увеличивает период полужизни активных мутантов Ras-GTP.[22]
Учредительно активный Рас
Учредительно активный Рас (РасD) - это тот, который содержит мутации, которые предотвращают гидролиз GTP, тем самым блокируя Ras в постоянно включенном состоянии.
Наиболее частые мутации обнаруживаются в остатке G12 в P-петля и каталитический остаток Q61.
- Мутация глицина в валин при остаток 12 делает домен GTPase Ras нечувствительным к инактивации GAP и, таким образом, застревает в состоянии «включено». Ras требует GAP для инактивации, поскольку сам по себе является относительно плохим катализатором, в отличие от других белков, содержащих G-домен, таких как альфа-субъединица гетеротримерных G-белков.
- Остаток 61[23] отвечает за стабилизацию переходного состояния для гидролиза GTP. Поскольку ферментативный катализ обычно достигается за счет снижения энергетического барьера между субстратом и продуктом, мутация Q61 в K (глутамин в лизин) обязательно снижает скорость внутреннего гидролиза Ras GTP до физиологически бессмысленных уровней.
См. Также «доминантно-отрицательные» мутанты, такие как S17N и D119N.
Ras-целевые методы лечения рака
Реовирус было отмечено, что он является потенциальным терапевтическим средством против рака, когда исследования показали, что он хорошо воспроизводится в определенных линиях раковых клеток. Он специфически реплицируется в клетках, которые имеют активированный путь Ras (клеточный сигнальный путь, который участвует в росте и дифференцировке клеток).[24] Реовирус реплицируется и в конечном итоге убивает Ras-активированные опухолевые клетки, и, когда происходит гибель клеток, дочерние вирусные частицы могут инфицировать окружающие раковые клетки. Считается, что этот цикл инфицирования, репликации и гибели клеток повторяется до тех пор, пока все опухолевые клетки, несущие активированный путь Ras, не будут уничтожены.
Другой вирус, лизирующий опухоли, который специфически поражает опухолевые клетки с активированным путем Ras, является вирусом типа II. Вирус простого герпеса (HSV-2) агент, обозначенный FusOn-H2.[25] Активирующие мутации белка Ras и вышестоящих элементов белка Ras могут играть роль в более чем двух третях всех раковых заболеваний человека, включая большинство метастатических заболеваний. Реолизин, состав реовируса и FusOn-H2 в настоящее время проходят клинические испытания или разрабатываются для лечения различных видов рака.[26] Кроме того, лечение, основанное на антимутантной миРНК К-РАН (G12D) под названием siG12D LODER в настоящее время проходит клинические испытания для лечения местно-распространенного рака поджелудочной железы (NCT01188785, NCT01676259).[27]
В моделях мышей с глиобластомой уровни SHP2 были повышены в раковых клетках мозга. Подавление SHP2 в свою очередь ингибировал дефосфорилирование Ras. Это уменьшило размеры опухоли и, как следствие, повысило выживаемость.[28][29]
Другие стратегии пытались манипулировать регуляцией вышеупомянутой локализации Ras. Ингибиторы фарнезилтрансферазы были разработаны, чтобы остановить фарнезилирование Раса и, следовательно, ослабляют его сродство к мембранам.[2] Другие ингибиторы нацелены на пальмитоилирование цикл Ras посредством ингибирования депальмитоилирования посредством ацилпротеиновые тиоэстеразы, что потенциально может привести к дестабилизации цикла Ras.[30]
использованная литература
- ^ Гудселл Д.С. (1999). «Молекулярная перспектива: онкоген ras». Онколог. 4 (3): 263–4. PMID 10394594.
- ^ а б Вниз J (январь 2003 г.). «Нацеливание на сигнальные пути RAS в терапии рака». Обзоры природы. Рак. 3 (1): 11–22. Дои:10.1038 / nrc969. PMID 12509763.
- ^ а б Malumbres M, Barbacid M (июнь 2003 г.). «Онкогены РАН: первые 30 лет». Обзоры природы. Рак. 3 (6): 459–65. Дои:10.1038 / nrc1097. PMID 12778136.
- ^ Чанг Э. Х., Гонда М. А., Эллис Р. В., Сколник Е. М., Лоуи Д. Р. (август 1982 г.). «Геном человека содержит четыре гена, гомологичных трансформирующим генам вирусов саркомы мышей Харви и Кирстен». Труды Национальной академии наук Соединенных Штатов Америки. 79 (16): 4848–52. Дои:10.1073 / пнас.79.16.4848. ЧВК 346782. PMID 6289320.
- ^ Харви Дж. Дж. (Декабрь 1964 г.). «Неопознанный вирус, который вызывает быстрое образование опухолей у мышей». Природа. 204 (4963): 1104–5. Дои:10.1038 / 2041104b0. PMID 14243400.
- ^ Кирстен У.Х., Шауф В., Маккой Дж. (1970). «Свойства вируса мышиной саркомы». Bibliotheca Haematologica. Международный симпозиум по сравнительным исследованиям лейкемии. 36 (36): 246–9. Дои:10.1159/000391714. ISBN 978-3-8055-1160-5. PMID 5538357.
- ^ Купер GM (август 1982 г.). «Клеточные трансформирующие гены». Наука. 217 (4562): 801–6. Дои:10.1126 / science.6285471. PMID 6285471.
- ^ Сантос Э., Троник С. Р., Ааронсон С. А., Пульчиани С., Барбакид М. (июль 1982 г.). «Онкоген карциномы мочевого пузыря человека T24 представляет собой активированную форму нормального человеческого гомолога трансформирующих генов BALB- и Harvey-MSV». Природа. 298 (5872): 343–7. Дои:10.1038 / 298343a0. PMID 6283384.
- ^ Парада Л.Ф., Табин С.Дж., Ши К., Вайнберг Р.А. (июнь 1982 г.). «Онкоген карциномы мочевого пузыря человека EJ является гомологом гена ras вируса саркомы Харви». Природа. 297 (5866): 474–8. Дои:10.1038 / 297474a0. PMID 6283357.
- ^ Тапаровски Е., Суард Ю., Фазано О., Шимицу К., Гольдфарб М., Виглер М. (декабрь 1982 г.). «Активация гена, трансформирующего карциному мочевого пузыря Т24, связана с одной заменой аминокислоты». Природа. 300 (5894): 762–5. Дои:10.1038 / 300762a0. PMID 7177195.
- ^ Marshall CJ, зал A, Weiss RA (сентябрь 1982 г.). «Трансформирующий ген, присутствующий в линиях клеток саркомы человека». Природа. 299 (5879): 171–3. Дои:10.1038 / 299171a0. PMID 6287287.
- ^ Зал A, Маршалл С.Дж., Спурр Н.К., Вайс Р.А. (1983). «Идентификация трансформирующего гена в двух линиях клеток саркомы человека как нового члена семейства генов ras, расположенного на хромосоме 1». Природа. 303 (5916): 396–400. Дои:10.1038 / 303396a0. PMID 6304521.
- ^ Симидзу К., Гольдфарб М., Перучо М., Виглер М. (январь 1983 г.). «Выделение и предварительная характеристика трансформирующего гена клеточной линии нейробластомы человека». Труды Национальной академии наук Соединенных Штатов Америки. 80 (2): 383–7. Дои:10.1073 / pnas.80.2.383. ЧВК 393381. PMID 6300838.
- ^ а б Веттер И.Р., Виттинггофер А (ноябрь 2001 г.). «Переключатель связывания гуаниновых нуклеотидов в трех измерениях». Наука. 294 (5545): 1299–304. Дои:10.1126 / science.1062023. PMID 11701921.
- ^ а б c Лодиш Х, Берк А, Зипурски С.Л., Мацудаира П., Балтимор Д., Дарнелл Дж. (2000). "Глава 25, Рак". Молекулярная клеточная биология (4-е изд.). Сан-Франциско: W.H. Фримен. ISBN 0-7167-3706-X.
- ^ Рокс О, Пейкер А, Бастиэнс П.И. (август 2006 г.). «Пространственно-временное разделение сигналов Ras: одно судно, три якоря, много гаваней». Текущее мнение в области клеточной биологии. 18 (4): 351–7. Дои:10.1016 / j.ceb.2006.06.007. PMID 16781855.
- ^ а б Bos JL (сентябрь 1989 г.). «Онкогены ras при раке человека: обзор». Исследования рака. 49 (17): 4682–9. PMID 2547513.
- ^ Веннерберг К., Россман К.Л., Der CJ (март 2005 г.). «Краткий обзор суперсемейства Ras». Журнал клеточной науки. 118 (Пт 5): 843–6. Дои:10.1242 / jcs.01660. PMID 15731001.
- ^ Роббинс и Котран (2010). Патологическая основа болезни 8-е изд.. п. 282.
- ^ Ротблат Б., Эрлих М., Хаклай Р., Клоог Ю. (2008). «Ингибитор Ras фарнезилтиосалициловая кислота (салиразиб) нарушает пространственно-временную локализацию активного Ras: потенциальное средство для лечения рака». Методы в энзимологии. 439: 467–89. Дои:10.1016 / S0076-6879 (07) 00432-6. ISBN 9780123743114. PMID 18374183.
- ^ Блюм Р., Якоб-Хирш Дж, Амариглио Н., Рехави Дж., Клоог И. (февраль 2005 г.). «Ингибирование Ras в глиобластоме подавляет индуцируемый гипоксией фактор-1альфа, вызывая остановку гликолиза и гибель клеток». Исследования рака. 65 (3): 999–1006. PMID 15705901.
- ^ Reuter CW, Morgan MA, Bergmann L (сентябрь 2000 г.). «Нацеливание на сигнальный путь Ras: рациональное, основанное на механизмах лечение гематологических злокачественных новообразований?». Кровь. 96 (5): 1655–69. Дои:10.1182 / кровь.V96.5.1655. PMID 10961860.
- ^ Омим - гомолог нейробластомы Ras вирусного онкогена; Nras[мертвая ссылка ]
- ^ Лал Р., Харрис Д., Постель-Виней С., де Боно Дж. (Октябрь 2009 г.). «Реовирус: обоснование и обновление клинических испытаний». Современное мнение о молекулярной терапии. 11 (5): 532–9. PMID 19806501.
- ^ Фу, Синьпин; Пригге-Дж, Цай-Р; Сяолиу Чжан. «Мутантный вирус простого герпеса 2 типа, удаленный по протеинкиназному домену гена ICP10, является мощным онколитическим вирусом». Молекулярная терапия. 13 (5): 882–890. Дои:10.1016 / j.ymthe.2006.08.180.
- ^ Тируккумаран К., Моррис Д.Г. (2009). «Онколитическая вирусная терапия с использованием реовируса». Методы молекулярной биологии. Методы молекулярной биологии ™. 542: 607–34. Дои:10.1007/978-1-59745-561-9_31. ISBN 978-1-934115-85-5. PMID 19565924.
- ^ "ClinicalTrials.gov".
- ^ Бунда С., Баррелл К., Наследник П., Зенг Л., Аламсахебпур А., Кано И., Рот Б., Чжан З.Й., Заде Г., Ооо М. (ноябрь 2015 г.). «Ингибирование SHP2-опосредованного дефосфорилирования Ras подавляет онкогенез». Nature Communications. 6: 8859. Дои:10.1038 / ncomms9859. ЧВК 4674766. PMID 26617336.
- ^ Тауб, Бен (2015-12-03). «Ученые нашли способ деактивировать самый распространенный белок, вызывающий рак». IFLScience. Получено 2016-02-20.
- ^ Чавда Б., Арнотт Дж. А., Плани С.Л. (сентябрь 2014 г.). «Нацеливание на пальмитоилирование белков: селективные ингибиторы и последствия для болезни». Мнение эксперта об открытии лекарств. 9 (9): 1005–19. Дои:10.1517/17460441.2014.933802. PMID 24967607.
дальнейшее чтение
- Agrawal AG, Somani RR (июнь 2009 г.). «Ингибитор фарнезилтрансферазы как противораковое средство». Мини-обзоры по медицинской химии. 9 (6): 638–52. Дои:10.2174/138955709788452702. PMID 19519490.
внешние ссылки
- «Открытие опухоли головного мозга дает надежду на новую стратегию, о которой сообщает Канадское онкологическое общество» на сайте ncic.cancer.ca
- «Новое лечение рака получает поддержку NCI» на arstechnica.com
- ras + белки в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- ras + гены в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- Дрозофила Онкоген Ras на 85D - Интерактивная муха
- «Анимация активации ras по EGFR»