ГСК-3 - GSK-3
| Гликогенсинтаза киназа 3, каталитический домен | |
|---|---|
| Идентификаторы | |
| Символ | STKc_GSK3 |
| ИнтерПро | IPR039192 |
| CDD | cd14137 |
| киназа гликогенсинтазы 3 альфа | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Символ | GSK3A | ||||||
| Ген NCBI | 2931 | ||||||
| HGNC | 4616 | ||||||
| OMIM | 606784 | ||||||
| RefSeq | NM_019884 | ||||||
| UniProt | P49840 | ||||||
| Прочие данные | |||||||
| Номер ЕС | 2.7.11.26 | ||||||
| Locus | Chr. 19 q13.2 | ||||||
| |||||||
| киназа гликогенсинтазы 3 бета | |||||||
|---|---|---|---|---|---|---|---|
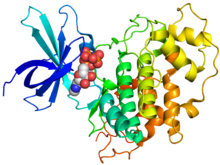 Кристаллографическая структура человеческого GSK-3β (цвета радуги, N-конец = синий, C-конец = красный), связанный с аденилатным эфиром фосфоаминофосфоновой кислоты (сферы).[1] | |||||||
| Идентификаторы | |||||||
| Символ | GSK3B | ||||||
| Ген NCBI | 2932 | ||||||
| HGNC | 4617 | ||||||
| OMIM | 605004 | ||||||
| PDB | 1Q3W Больше структур | ||||||
| RefSeq | NM_002093 | ||||||
| UniProt | P49841 | ||||||
| Прочие данные | |||||||
| Номер ЕС | 2.7.11.26 | ||||||
| Locus | Chr. 3 q13.33 | ||||||
| |||||||
Киназа гликогенсинтазы 3 (ГСК-3) это серин / треониновая протеинкиназа что опосредует добавление фосфат молекулы на серин и треонин аминокислотные остатки. Впервые обнаружена в 1980 году как регуляторная киназа для своего тезки, гликогенсинтаза (GS),[2] ГСК-3 с тех пор был идентифицирован как протеинкиназа для более чем 100 различных белков различными путями.[3][4] У млекопитающих, включая человека, GSK-3 существует в двух изоформы закодировано двумя паралогичный гены GSK-3α (GSK3A ) и GSK-3β (GSK3B ). GSK-3 был предметом многочисленных исследований, поскольку он был вовлечен в ряд заболеваний, включая диабет 2 типа, Болезнь Альцгеймера, воспаление, рак, и биполярное расстройство.
GSK-3 представляет собой серин / треониновую протеинкиназу, которая фосфорилировать либо треонин или же серин, и это фосфорилирование контролирует различные биологические активности, такие как гликоген метаболизм, клеточная сигнализация, сотовый транспорт, и другие.[5] Ингибирование GS с помощью GSK-3β приводит к снижению синтеза гликогена в печени и мышцах, а также к повышению уровня глюкозы в крови или гипергликемии.[6] Вот почему GSK-3β связан с патогенезом и прогрессированием многих заболеваний, таких как сахарный диабет, ожирение, рак,[7] и болезнь Альцгеймера.[8] Он активен в покоящихся клетках и подавляется несколькими гормонами, такими как инсулин, фактор роста эндотелия, и фактор роста тромбоцитов. Инсулин инактивирует его путем фосфорилирования специфических остатков серина Ser21 и Ser9 в изоформах GSK-3 α и β соответственно. В зависимости от фосфатидилинозитол-3-киназы.[нужна цитата ]
По состоянию на 2019 год[Обновить], ГСК-3 - единственный тип киназа гликогенсинтазы названы и признаны. В символы генов для GSK1 и GSK2 были отозваны Комитет по номенклатуре генов HUGO (HGNC), и никаких новых названий для этих «генов» или их местоположения не определено.[9][10]
Механизм
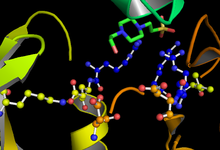
ГСК-3 функционирует фосфорилирование остаток серина или треонина на целевом субстрате. Положительно заряженный карман, примыкающий к активному сайту, связывает «затравочную» фосфатную группу, присоединенную к С-концу серина или треонина с четырьмя остатками целевого сайта фосфорилирования. Активный сайт в остатках 181, 200, 97 и 85 связывает концевой фосфат АТФ и переносит его в целевое место на субстрате (см. Рисунок 1).[11]
Гликоген-синтаза
Гликоген-синтаза является фермент это отвечает в гликоген синтез. Он активируется глюкозо-6-фосфат (G6P) и ингибируется киназы гликогенсинтазы (GSK3 ). Эти два механизма играют важную роль в метаболизме гликогена.[12]
Функция
Фосфорилирование белка GSK-3 обычно подавляет активность его нижерасположенной мишени.[13][14][15] GSK-3 активен в ряде центральных внутриклеточных сигнальных путей, включая клеточную пролиферацию, миграцию, регуляцию глюкозы и апоптоз.
ГСК-3 был первоначально открыт в контексте его участия в регулировании гликогенсинтаза.[2] После грунтовки казеин киназа 2 (CK2), гликогенсинтаза фосфорилируется по кластеру из трех С-концевых остатков серина, снижая ее активность.[16] Помимо своей роли в регуляции гликогенсинтазы, GSK-3 участвует в других аспектах гомеостаза глюкозы, включая фосфорилирование рецептора инсулина. IRS1 [17] и глюконеогенных ферментов фосфоенолпируваткарбоксикиназа и глюкозо-6-фосфатаза.[18] Однако эти взаимодействия не были подтверждены, поскольку эти пути могут подавляться без повышения регуляции GSK-3.[16]
Было также показано, что GSK-3 регулирует иммунные и миграционные процессы. GSK-3 участвует в ряде сигнальных путей врожденного иммунного ответа, включая выработку провоспалительных цитокинов и интерлейкинов.[19][20] Инактивация GSK3B с помощью различных протеинкиназ также влияет на адаптивный иммунный ответ, индуцируя продукцию и пролиферацию цитокинов в наивных CD4 + Т-клетках и Т-клетках памяти.[20] Сообщалось, что в клеточной миграции, неотъемлемом аспекте воспалительных реакций, ингибирование GSK-3 играет противоречивую роль, поскольку было показано, что локальное ингибирование в конусах роста способствует подвижности, в то время как глобальное ингибирование клеточного GSK-3 ингибирует распространение и миграция клеток.[19]
GSK-3 также неразрывно связан с путями пролиферации клеток и апоптоза. Было показано, что GSK-3 фосфорилирует Бета-катенин, таким образом направляя его на деградацию.[21] Таким образом, ГСК-3 является частью канонической Бета-катенин /Wnt путь, который сигнализирует клетке о делении и пролиферации. GSK-3 также участвует в ряде апоптотических сигнальных путей, фосфорилируя факторы транскрипции, которые регулируют апоптоз.[4] GSK-3 может способствовать апоптозу за счет активации проапоптотических факторов, таких как p53 [22] и инактивация факторов, способствующих выживанию, посредством фосфорилирования.[23] Роль GSK-3 в регуляции апоптоза является спорным, однако, так как некоторые исследования показали, что GSK-3 & beta; нокаутный мышей чрезмерно чувствительные к апоптозу и погибают в эмбриональной стадии, в то время как другие показали, что избыточная экспрессия GSK-3 может индуцировать апоптоз .[24] В целом, GSK-3, по-видимому, способствует и ингибирует апоптоз, и эта регуляция варьируется в зависимости от конкретного молекулярного и клеточного контекста.[25]
Регулирование
Из-за его важности для множества клеточных функций активность GSK-3 строго регулируется.
Скорость и эффективность фосфорилирования GSK-3 регулируется рядом факторов. Фосфорилирование определенных остатков GSK-3 может увеличивать или уменьшать его способность связывать субстрат. Фосфорилирование тирозина-216 в GSK-3β или тирозина-279 в GSK-3α усиливает ферментативную активность GSK-3, тогда как фосфорилирование серина-9 в GSK-3β или серина-21 в GSK-3α значительно снижает доступность активного сайта ( см. рисунок 1).[19] Кроме того, GSK-3 необычен среди киназ тем, что для первого фосфорилирования субстрата ему обычно требуется «праймирующая киназа». Фосфорилированный остаток серина или треонина, расположенный на С-конце четырех аминокислот от целевого сайта фосфорилирования, позволяет субстрату связывать карман с положительным зарядом, образованный остатками аргинина и лизина.[16][26]
В зависимости от пути, в котором он используется, GSK-3 может дополнительно регулироваться клеточной локализацией или образованием белковых комплексов. Активность GSK-3 намного выше в ядре и митохондриях, чем в цитозоле корковых нейронов,[27] в то время как фосфорилирование бета-катенина GSK-3 опосредуется связыванием обоих белков с Axin, белок каркаса, позволяющий бета-катенину получать доступ к активному сайту GSK-3.[19]
Актуальность болезни
Из-за своего участия в большом количестве сигнальных путей GSK-3 ассоциирован с множеством высокопрофильных заболеваний. Ингибиторы GSK-3 в настоящее время проходят испытания на терапевтическое действие при Болезнь Альцгеймера, сахарный диабет 2 типа (СД2), некоторые формы рак, и биполярное расстройство.[28]
Теперь было показано, что литий, который используется для лечения биполярное расстройство, действует как стабилизатор настроения, избирательно подавляя GSK-3. Механизм, с помощью которого ингибирование GSK-3 стабилизирует настроение, неизвестен, хотя есть подозрения, что ингибирование способности GSK-3 вызывать воспаление способствует терапевтическому эффекту.[19] Ингибирование GSK-3 также дестабилизирует репрессор транскрипции Rev-ErbA альфа, который играет важную роль в циркадных часах.[29] Элементы циркадных часов могут быть связаны с предрасположенностью к биполярному расстройству настроения.[30]
Активность GSK-3 была связана с обеими патологическими особенностями болезни Альцгеймера, а именно с накоплением амилоид-β (Aβ) отложения и образование нейрофибриллярные сплетения. Считается, что GSK-3 непосредственно способствует продукции Aβ и связан с процессом гиперфосфорилирование из тау-белки, что приводит к путанице.[4][19] Благодаря этой роли GSK-3 в развитии болезни Альцгеймера, ингибиторы GSK-3 могут оказывать положительное терапевтическое воздействие на пациентов с болезнью Альцгеймера и в настоящее время находятся на ранних стадиях тестирования.[31]
Подобным образом целевое ингибирование GSK-3 может оказывать терапевтическое воздействие на определенные виды рака. Хотя было показано, что ГСК-3 продвигает апоптоз в некоторых случаях это также было ключевым фактором в туморогенез при некоторых раках.[32] В поддержку этого утверждения было показано, что ингибиторы GSK-3 вызывают апоптоз в клетках глиомы и рака поджелудочной железы.[24][33]
Ингибиторы GSK-3 также показали себя многообещающими при лечении СД2.[16] Хотя активность GSK-3 в условиях диабета может радикально различаться для разных типов тканей, исследования показали, что введение конкурентных ингибиторов GSK-3 может повысить толерантность к глюкозе у диабетических мышей.[19] Ингибиторы GSK-3 могут также оказывать терапевтическое действие на геморрагическую трансформацию после острого ишемического инсульта.[34] GSK-3 может негативно регулировать путь передачи сигналов инсулина путем ингибирования IRS1 посредством фосфорилирования серина-332,[17] делает рецептор инсулина неспособным активировать IRS1 и далее инициировать канонический путь PI3K / Akt. Роль, которую ингибирование GSK-3 может играть в др. Его сигнальных ролях, еще полностью не изучена.
Ингибирование GSK-3 также опосредует увеличение транскрипции фактора транскрипции Tbet (Tbx21) и ингибирование транскрипции ингибирующего корецептора запрограммированной гибели клеток-1 (PD-1) на Т-клетках.[35] Ингибиторы GSK-3 увеличивали in vivo функцию CTL CD8 (+) OT-I и устранение вирусных инфекций мышиным гамма-герпесвирусом 68 и клоном 13 лимфоцитарного хориоменингита, а также анти-PD-1 в иммунотерапии.
Ингибиторы
Ингибиторы киназы гликогенсинтазы разные хемотипы и имеют различные механизмы действия; они могут быть катионы, из природных источников, синтетические АТФ и не-АТФ-конкурентные ингибиторы и субстрат-конкурентные ингибиторы. GSK3 - это двухлепестковая архитектура с N-концевой и C-терминал, N-конец отвечает за связывание АТФ, а C-конец, который называется цикл активации опосредует активность киназы, тирозин, расположенный на С-конце, необходим для полной активности GSK3.[36]
Преимущества ингибиторов GSK-3β
При диабете ингибиторы GSK-3β повышают чувствительность к инсулину, синтез гликогена и метаболизм глюкозы в скелетных мышцах, а также снижают ожирение, воздействуя на адипогенез процесс.[37] GSK-3β также сверхэкспрессируется при некоторых типах рака, таких как колоректальный, яичник, и рак простаты.[36] Ингибиторы GSK-3β также помогают в лечении Болезнь Альцгеймера,[нужна цитата ] Инсульт,[нужна цитата ] и расстройства настроения, включая биполярное расстройство.[38]
Специфические агенты
К ингибиторам ГСК-3 относятся:[39][40][41]
Катионы металлов
ATP-конкурентоспособный
Полученный из морских организмов
- 6-БИО (IC50= 1,5 мкм)
- Дибромокантареллин (IC50= 3 мкм)
- Гимениалдезин (IC50= 10 нм)
- Индирубин (IC50= 5-50 нм)
- Меридианин
Аминопиримидины
Арилиндолемалеимид
Тиазолы
- AR-A014418 (IC50= 104 нМ)
- АЗД-1080
Пауллонес
IC50= 4-80 нм:
Алоизины
IC50= 0,5-1,5 мкм:
Неконкурентоспособный ATP
Полученный из морских организмов
- Манзамин А (IC50= 1,5 мкм)
- Палинурин (IC50= 4,5 мкМ)
- Трикантин (IC50= 7,5 мкм)
Тиазолидиндионы
- ТДЗД-8 (IC50= 2 мкм)
- NP00111 (IC50= 2 мкм)
- NP031115 (IC50= 4 мкм)
- Тидеглусиб
Галометилкетоны
- ХМК-32 (IC50= 1,5 мкм)
Пептиды
Неизвестный механизм (низкомолекулярные ингибиторы)
Литий
Литий, который используется для лечения биполярное расстройство был первым обнаруженным природным ингибитором GSK-3. Он ингибирует GSK-3 напрямую, конкурируя с ионами магния, и косвенно, путем фосфорилирования и саморегуляции серина. Было обнаружено, что литий оказывает инсулино-подобное действие на метаболизм глюкозы, включая стимуляцию синтеза гликогена в жировых клетках, коже и мышцах. увеличение поглощения глюкозы и активация активности GS. Помимо ингибирования GSK-3, он также ингибирует другие ферменты, участвующие в регуляции метаболизма глюкозы, такие как мио-инозитол-1-монофосфатаза и 1,6-бисфосфатаза. Кроме того, он показал терапевтическую пользу при болезни Альцгеймера и других нейродегенеративных заболеваниях, таких как эпилептическая нейродегенерация.[41]
Напроксен и Кромолин
Напроксен - это нестероидный противовоспалительный препарат в то время как кромолин является противоаллергическим средством, которое действует как тучная клетка стабилизатор. Оба препарата продемонстрировали противоопухолевый эффект в дополнение к гипогликемическому эффекту за счет ингибирования киназы-3β гликогенсинтазы (GSK-3β).
Для подтверждения гипотезы напроксена и кромолина об анти-GSK-3β, в дополнение к измерению уровня глюкозы в сыворотке крови, инсулина сыворотки крови и сравнения их соответствия с известным ингибитором GSK-3β, ARA014418, была проведена стыковка этих двух структур с карманом связывания GSK-3β. сывороточный С-пептид, изменение веса и уровни гликогена в печени для нормальных и диабетических моделей животных натощак для оценки их гипогликемических эффектов in vitro.[нужна цитата ]
Напроксен и кромолин были успешно прикреплены к сайту связывания GSK-3β (оба были помещены в его карман связывания). Они проявляют электростатические, гидрофобные и водородные взаимодействия с ключевыми аминокислотами внутри карман для переплета с профилями связывающего взаимодействия, аналогичными AR-A014418 (известный ингибитор). Отрицательные заряды групп карбоновой кислоты в обоих препаратах электростатически взаимодействуют с положительно заряженной гуанидиновой группой Arg141. Более того, взаимодействия водородных связей между остатками карбоновых кислот кромолина и аммониевыми группами Lys183 и Lys60, в дополнение к π-укладке нафталиновой кольцевой системы напроксена с фенольным кольцом Tyr134.
Антидиабетические эффекты напроксена и кромолина: на моделях нормальных животных оба препарата показали дозозависимое снижение уровня глюкозы в крови и повышение уровня гликогена. В модели хронического диабета II типа уровни глюкозы также снижались, а уровень гликогена и уровни инсулина повышались дозозависимым образом с уменьшением глюкозы в плазме.[нужна цитата ]
Эффекты напроксена и кромолина против ожирения: оба препарата показали значительные эффекты против ожирения, поскольку они снижают массу тела, уровни резистина и глюкозы в зависимости от дозы. Было также обнаружено, что они повышают адипонектин, инсулина и С-пептида дозозависимым образом.[37]
Фамотидин
Фамотидин - специфический, длительного действия Н2 антагонист что снижает секрецию желудочного сока. Он используется при лечении язвенной болезни, ГЭРБ и патологических гиперсекреторных состояний, таких как синдром Золлингера – Эллисона. (14,15) Антагонисты Н2-рецепторов влияют на метаболизм гормонов, но их влияние на метаболизм глюкозы точно не установлено. (16) Исследование показало, что фамотидин снижает уровень глюкозы. Недавно был проведен молекулярный докинг в качестве предварительного скринингового теста in-silico для изучения связывания фамотидина с активным сайтом GSK-3β.[42]
Изучение связывания фамотидина с ферментом показало, что фамотидин может быть закреплен в кармане связывания GSK-3β, обеспечивая значительные взаимодействия с ключевыми точками внутри кармана связывания GSK-3β. Взаимодействие с сильными водородными связями с ключевыми аминокислотами PRO-136 и VAL-135 и потенциальное гидрофобное взаимодействие с LEU-188 были аналогичны тем, которые обнаружены при связывании лиганда с ферментом (AR-A014418).[нужна цитата ]
Кроме того, фамотидин показал высокую аффинность связывания GSK-3β и ингибирующую активность из-за взаимодействий, которые стабилизируют комплекс, а именно водородного связывания гуанидиновой группы в фамотидине с сульфагидрильной частью в CYS-199; и электростатические взаимодействия между той же группой гуанидина с карбоксильной группой в ASP-200, водородная связь между концевой группой NH2, ОН в TYR-143 и гидрофобное взаимодействие атома серы в тиоэфире с ILE-62. Исследования in vitro показали, что фамотидин подавляет активность GSK-3β и увеличивает запасы гликогена в печени в зависимости от дозы. Наблюдалось четырехкратное повышение уровня гликогена в печени при применении максимальной дозы фамотидина (4,4 мг / кг). Кроме того, было показано, что фамотидин снижает уровень глюкозы в сыворотке через 30 и 60 минут после пероральной глюкозной нагрузки у здоровых людей.[43]
Куркумин
Куркумин, входящий в состав куркума пряность, обладает вкусовыми и красящими свойствами.[44] Он имеет две симметричные формы: енол (наиболее распространенные формы) и кетон.[45][46]
Куркумин обладает широким фармакологическим действием: противовоспалительным,[47] антимикробный,[48] гипогликемический, антиоксидантный и ранозаживляющий эффекты.[49] На животных моделях с болезнью Альцгеймера он оказывает анти-деструктивный эффект бета-амилоида в головном мозге,[50] а в последнее время проявляет противомалярийную активность.[51]
Куркумин также обладает химиопрофилактическим и противораковым действием.[цитата не найдена ], и было показано, что он снижает окислительный стресс и почечную дисфункцию у диабетических животных при хроническом употреблении.[52]
Механизм действия куркумина - противовоспалительный; он ингибирует ядерный активатор транскрипции каппа B (NF-KB ), который активируется всякий раз, когда возникает воспалительная реакция.[нужна цитата ]
NF-kB имеет два регулирующих фактора, IkB и GSK-3,[53] что предполагает, что куркумин напрямую связывает и ингибирует GSK-3B. Исследование in vitro подтвердило ингибирование GSK-3B путем моделирования молекулярного стыковки с использованием метода силикона.[54] Концентрация, при которой куркумин ингибирует 50% GK-3B, составляет 66,3 нМ.[54]
Среди двух его форм экспериментальные и теоретические исследования показывают, что енольная форма является предпочтительной формой из-за ее внутримолекулярных водородных связей, а эксперимент ЯМР показывает, что енольная форма существует в различных растворителях.[нужна цитата ]
Оланзапин
Нейролептики лекарства все чаще используются для шизофрения, биполярное расстройство, тревожность и другие психические расстройства[55] Атипичные нейролептики используются чаще, чем нейролептики первого поколения, поскольку они снижают риск экстрапирамидных симптомов, таких как поздняя дискинезия, и имеют лучшую эффективность.[56]
Оланзапин и атипичные нейролептики вызывают увеличение веса за счет увеличения жировых отложений.[57] Он также влияет на метаболизм глюкозы, и несколько исследований показывают, что он может усугубить диабет.[58]
Недавнее исследование показывает, что оланзапин подавляет активность GSK3, что предполагает, что оланзапин способствует синтезу гликогена. Исследование влияния оланзапина на уровни глюкозы и гликогена в крови мышей показало значительное снижение уровня глюкозы в крови и повышение уровня гликогена у мышей, а IC50% оланзапина составляла 91,0 нМ, что считается мощным ингибитором. Исследование также показывает, что субхроническое использование оланзапина приводит к сильному ингибированию GSK3.[38]
Производные пиримидина
Аналоги пиримидина антиметаболиты, которые мешают синтезу нуклеиновых кислот.[59] Было показано, что некоторые из них подходят для АТФ-связывающего кармана GSK-3β для снижения уровня глюкозы в крови и лечения некоторых нейрональных заболеваний.[60]
Смотрите также
Рекомендации
- ^ PDB: 1J1B; Аоки М., Йокота Т., Сугиура И., Сасаки С., Хасэгава Т., Окумура К., Исигуро К., Коно Т., Сугио С., Мацудзаки Т. (март 2004 г.). «Структурное понимание распознавания нуклеотидов в тау-протеинкиназе I / гликогенсинтазе киназе 3 бета». Acta Crystallographica Раздел D. 60 (Pt 3): 439–46. Дои:10.1107 / S090744490302938X. PMID 14993667.
- ^ а б Эмби Н., Рилатт Д. Б., Коэн П. (июнь 1980 г.). «Киназа-3 гликогенсинтазы из скелетных мышц кролика. Отделение от циклической АМФ-зависимой протеинкиназы и фосфорилазы киназы». Европейский журнал биохимии. 107 (2): 519–27. Дои:10.1111 / j.1432-1033.1980.tb06059.x. PMID 6249596.
- ^ Beurel E, Grieco SF, Jope RS (апрель 2015 г.). «Киназа-3 гликогенсинтазы (GSK3): регуляция, действие и заболевания». Фармакология и терапия. 148: 114–131. Дои:10.1016 / j.pharmthera.2014.11.016. ЧВК 4340754. PMID 25435019.
- ^ а б c Джоуп Р.С., Джонсон Г.В. (февраль 2004 г.). «Очарование и мрак киназы-3 гликогенсинтазы». Тенденции в биохимических науках. 29 (2): 95–102. Дои:10.1016 / j.tibs.2003.12.004. PMID 15102436.
- ^ Pandey MK, DeGrado TR (2016). «Гликоген-синтаза-киназа-3 (GSK-3) - таргетированная терапия и визуализация». Тераностика. 6 (4): 571–93. Дои:10.7150 / thno.14334. ЧВК 4775866. PMID 26941849.
- ^ Али А., Хефлих К.П., Вудгетт-младший (август 2001 г.). «Гликоген-синтаза киназа-3: свойства, функции и регуляция». Химические обзоры. 101 (8): 2527–40. Дои:10.1021 / cr000110o. PMID 11749387.
- ^ Эльдар-Финкельман Х (март 2002 г.). «Киназа гликогенсинтазы 3: новая терапевтическая мишень». Тенденции в молекулярной медицине. 8 (3): 126–32. Дои:10.1016 / S1471-4914 (01) 02266-3. PMID 11879773.
- ^ Хупер С., Киллик Р., Лавстон С. (март 2008 г.). «Гипотеза GSK3 болезни Альцгеймера». Журнал нейрохимии. 104 (6): 1433–9. Дои:10.1111 / j.1471-4159.2007.05194.x. ЧВК 3073119. PMID 18088381.
- ^ Гликоген + синтаза + киназа в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- ^ GSK1, GSK2. NCBI Gene.
- ^ Даджани Р., Фрейзер Е., Роу С.М., Янг Н., Гуд В., Дейл Т.С., Перл Л.Х. (июнь 2001 г.). «Кристаллическая структура киназы гликогенсинтазы 3 бета: структурная основа для специфичности и аутоингибирования фосфат-примированного субстрата». Клетка. 105 (6): 721–32. Дои:10.1016 / S0092-8674 (01) 00374-9. PMID 11440715. S2CID 17401752.
- ^ Бускила М., Хантер Р. В., Ибрагим А. Ф., Делаттр Л., Пегги М., ван Дипен Дж. А., Фошол П. Дж., Дженсен Дж., Сакамото К. (ноябрь 2010 г.). «Аллостерическая регуляция гликогенсинтазы контролирует синтез гликогена в мышцах». Клеточный метаболизм. 12 (5): 456–66. Дои:10.1016 / j.cmet.2010.10.006. PMID 21035757.
- ^ Вудгетт-младший (август 1994 г.). «Регуляция и функции подсемейства киназы-3 гликогенсинтазы». Семинары по биологии рака. 5 (4): 269–75. PMID 7803763.
- ^ Вудгетт-младший (сентябрь 2001 г.). «Судить о протеине не только по его названию: GSK-3». STKE науки. 2001 (100): re12. Дои:10.1126 / stke.2001.100.re12. PMID 11579232. S2CID 19052833.
- ^ Али А., Хефлих К.П., Вудгетт-младший (август 2001 г.). «Гликогенсинтаза киназа-3: свойства, функции и регуляция». Химические обзоры. 101 (8): 2527–40. Дои:10.1021 / cr000110o. PMID 11749387. S2CID 12925005.
- ^ а б c d Райасам Г.В., Туласи В.К., Содхи Р., Дэвис Дж. А., Луч А. (март 2009 г.). «Гликоген-синтазокиназа 3: больше, чем однофамилец». Британский журнал фармакологии. 156 (6): 885–98. Дои:10.1111 / j.1476-5381.2008.00085.x. ЧВК 2697722. PMID 19366350.
- ^ а б Либерман З., Эльдар-Финкельман Х (февраль 2005 г.). «Фосфорилирование серина 332 субстрата-1 рецептора инсулина с помощью киназы-3 гликогенсинтазы ослабляет передачу сигналов инсулина». Журнал биологической химии. 280 (6): 4422–8. Дои:10.1074 / jbc.M410610200. PMID 15574412.
- ^ Локхед ПА, Коглан М., Райс С.К., Сазерленд С. (май 2001 г.). «Ингибирование GSK-3 избирательно снижает экспрессию генов глюкозо-6-фосфатазы и фосфатазы и фосфоенолипируваткарбоксикиназы». Сахарный диабет. 50 (5): 937–46. Дои:10.2337 / диабет.50.5.937. PMID 11334436.
- ^ а б c d е ж грамм Джоуп Р.С., Юскайтис К.Дж., Бурел Э. (апрель – май 2007 г.). «Киназа-3 гликогенсинтазы (GSK3): воспаление, заболевания и терапия». Нейрохимические исследования. 32 (4–5): 577–95. Дои:10.1007 / s11064-006-9128-5. ЧВК 1970866. PMID 16944320.
- ^ а б Ван Х, Браун Дж, Мартин М. (февраль 2011 г.). «Киназа гликоген-синтазы 3: точка конвергенции воспалительного ответа хозяина». Цитокин. 53 (2): 130–40. Дои:10.1016 / j.cyto.2010.10.009. ЧВК 3021641. PMID 21095632.
- ^ Миллс С.Н., Новшин С., Боннер Дж. А., Ян Э. С. (2011). «Новые роли киназы гликогенсинтазы 3 в лечении опухолей головного мозга». Границы молекулярной неврологии. 4: 47. Дои:10.3389 / фнмол.2011.00047. ЧВК 3223722. PMID 22275880.
- ^ Watcharasit P, Bijur GN, Zmijewski JW, Song L, Zmijewska A, Chen X, Johnson GV, Jope RS (июнь 2002 г.). «Прямое активирующее взаимодействие между киназой-3бета гликогенсинтазы и р53 после повреждения ДНК». Труды Национальной академии наук Соединенных Штатов Америки. 99 (12): 7951–5. Bibcode:2002PNAS ... 99,7951 Вт. Дои:10.1073 / pnas.122062299. ЧВК 123001. PMID 12048243.
- ^ Grimes CA, Jope RS (сентябрь 2001 г.). «Активность связывания ДНК CREB ингибируется киназой-3 бета гликогенсинтазы и облегчается литием». Журнал нейрохимии. 78 (6): 1219–32. Дои:10.1046 / j.1471-4159.2001.00495.x. ЧВК 1947002. PMID 11579131.
- ^ а б Котлярова С., Пасторино С., Ковелл Л.С., Котляров Ю., Сонг Х., Чжан В., Бейли Р., Марич Д., Зенклусен Дж. К., Ли Дж., Файн Х.А. (август 2008 г.). «Ингибирование киназы-3 гликоген-синтазы вызывает гибель клеток глиомы за счет c-MYC, ядерного фактора-каппаB и регуляции глюкозы». Исследования рака. 68 (16): 6643–51. Дои:10.1158 / 0008-5472.CAN-08-0850. ЧВК 2585745. PMID 18701488.
- ^ Джейкобс К.М., Бхаве С.Р., Ферраро Д.Дж., Джабоин Д.Дж., Халлахан Д.Е., Тхотала Д. (май 2012 г.). «GSK-3β: бифункциональная роль в путях клеточной смерти». Международный журнал клеточной биологии. 2012: 930710. Дои:10.1155/2012/930710. ЧВК 3364548. PMID 22675363.
- ^ Doble BW, Woodgett JR (апрель 2003 г.). «ГСК-3: хитрости для многозадачной киназы». Журнал клеточной науки. 116 (Pt 7): 1175–86. Дои:10.1242 / jcs.00384. ЧВК 3006448. PMID 12615961.
- ^ Биджур Г. Н., Джоп Р. С. (декабрь 2003 г.). «Киназа-3 бета гликоген-синтазы высокоактивна в ядрах и митохондриях». NeuroReport. 14 (18): 2415–9. Дои:10.1097/00001756-200312190-00025. PMID 14663202. S2CID 43633965.
- ^ Сарасвати А.П., Али Хусайни С.М., Кришна Н.Х., Бабу Б.Н., Камаль А. (январь 2018 г.). «Киназа-3 гликогенсинтазы и ее ингибиторы: потенциальная мишень для различных терапевтических состояний». Европейский журнал медицинской химии. 144: 843–858. Дои:10.1016 / j.ejmech.2017.11.103. PMID 29306837.
- ^ Инь Л., Ван Дж, Кляйн П.С., Лазар М.А. (февраль 2006 г.). «Ядерный рецептор Rev-erbalpha - критический литий-чувствительный компонент циркадных часов». Наука. 311 (5763): 1002–5. Bibcode:2006Научный ... 311.1002Y. Дои:10.1126 / science.1121613. PMID 16484495. S2CID 11240826.
- ^ Рыбаковский Дж. К., Дмитрзак-Вегларц М., Дембинская-Краевская Д., Хаузер Дж., Акискал К. К., Акискал Х. Х. (апрель 2014 г.). «Полиморфизм генов циркадных часов и темпераментных измерений TEMPS-A при биполярном расстройстве». Журнал аффективных расстройств. 159: 80–4. Дои:10.1016 / j.jad.2014.02.024. PMID 24679394.
- ^ Ху С., Бегум А.Н., Джонс М.Р., О М.С., Бук В.К., Бук Б.Х., Ян Ф., Чен П., Убеда О.Дж., Ким П.С., Дэвис П., Ма К., Коул Г.М., Frautschy SA (февраль 2009 г.). «Ингибиторы GSK3 показывают преимущества в модели нейродегенерации при болезни Альцгеймера (БА), но побочные эффекты у контрольных животных». Нейробиология болезней. 33 (2): 193–206. Дои:10.1016 / j.nbd.2008.10.007. ЧВК 4313761. PMID 19038340.
- ^ Ван З., Смит К.С., Мерфи М., Пилото О., Сомервейл Т.С., Клири М.Л. (октябрь 2008 г.). «Киназа гликогенсинтазы 3 в поддержании и таргетной терапии лейкоза MLL». Природа. 455 (7217): 1205–9. Bibcode:2008 Натур.455.1205W. Дои:10.1038 / природа07284. ЧВК 4084721. PMID 18806775.
- ^ Marchand B, Tremblay I, Cagnol S, Boucher MJ (март 2012 г.). «Ингибирование активности киназы-3 гликогенсинтазы запускает апоптотический ответ в раковых клетках поджелудочной железы через JNK-зависимые механизмы». Канцерогенез. 33 (3): 529–37. Дои:10.1093 / carcin / bgr309. PMID 22201186.
- ^ Ван В., Ли М, Ван И, Ли Кью, Дэн Дж, Ван Дж, Ян Кью, Чен Кью, Ван Дж (декабрь 2016 г.). «Ингибитор GSK-3β TWS119 ослабляет геморрагическую трансформацию, вызванную rtPA, и активирует путь передачи сигналов Wnt / β-катенин после острого ишемического инсульта у крыс». Молекулярная нейробиология. 53 (10): 7028–7036. Дои:10.1007 / s12035-015-9607-2. ЧВК 4909586. PMID 26671619.
- ^ Тейлор А., Харкер Дж. А., Чантонг К., Стивенсон П. Г., Зунига Е. И., Радд К. Э. (февраль 2016 г.). «Инактивация киназы гликоген-синтазы 3 вызывает опосредованное T-bet подавление корецептора PD-1 для усиления CD8 (+) цитолитических Т-клеточных ответов». Иммунитет. 44 (2): 274–86. Дои:10.1016 / j.immuni.2016.01.018. ЧВК 4760122. PMID 26885856.
- ^ а б Sayas CL, Ariaens A, Ponsioen B, Moolenaar WH (апрель 2006 г.). «GSK-3 активируется тирозинкиназой Pyk2 во время LPA1-опосредованного ретракции нейритов». Молекулярная биология клетки. 17 (4): 1834–44. Дои:10.1091 / mbc.E05-07-0688. ЧВК 1415316. PMID 16452634.
- ^ а б Motawi TM, Bustanji Y, El-Maraghy SA, Taha MO, Al Ghussein MA (сентябрь 2013 г.). «Напроксен и кромолин как новые ингибиторы киназы гликогенсинтазы 3β для облегчения диабета и ожирения: исследование путем моделирования стыковки и последующей биохимической оценки in vitro / in vivo». Журнал биохимической и молекулярной токсикологии. 27 (9): 425–36. Дои:10.1002 / jbt.21503. PMID 23784744. S2CID 46597394.
- ^ а б Мохаммад М.К., Аль-Масри И.М., Таха М.О., Аль-Гуссейн М.А., Алхатиб Х.С., Наджар С., Бустанджи Й. (апрель 2008 г.). «Оланзапин ингибирует киназу-3бета гликогенсинтазы: исследование путем моделирования стыковки и экспериментальной проверки». Европейский журнал фармакологии. 584 (1): 185–91. Дои:10.1016 / j.ejphar.2008.01.019. PMID 18295757.
- ^ Noori MS, Bhatt PM, Courreges MC, Ghazanfari D, Cuckler C, Orac CM, McMills MC, Schwartz FL, Deosarkar SP, Bergmeier SC, McCall KD, Goetz DJ (декабрь 2019 г.). «Идентификация нового селективного и мощного ингибитора киназы-3 гликогенсинтазы». Американский журнал физиологии. Клеточная физиология. 317 (6): C1289 – C1303. Дои:10.1152 / ajpcell.00061.2019. ЧВК 6962522. PMID 31553649.
- ^ Лихт-Мурава А., Паз Р., Вакс Л., Аврахами Л., Плоткин Б., Эйзенштейн М., Эльдар-Финкельман Х. (ноябрь 2016 г.). «Уникальный вид ингибитора GSK-3 открывает новые возможности для клиники». Научная сигнализация. 9 (454): ra110. Дои:10.1126 / scisignal.aah7102. PMID 27902447. S2CID 34207388.
- ^ а б Эльдар-Финкельман Х, Мартинес А (2011). «Ингибиторы GSK-3: доклинические и клинические исследования ЦНС». Границы молекулярной неврологии. 4: 32. Дои:10.3389 / fnmol.2011.00032. ЧВК 3204427. PMID 22065134.
- ^ Дас П.К., Мостофа М., Хасан К., Шил Г.К., Ислам М.Р. (2003). «Влияние ранитидина, фамотидина и омепразола на некоторые гематобиохимические параметры у мышей». Журнал достижений животных и ветеринарии. 2: 321–6.
- ^ Мохаммад М., Аль-Масри И.М., Исса А., Аль-Гусейн М.А., Фарарджех М., Алхатиб Х., Таха М.О., Бустанджи Ю. (август 2013 г.). «Фамотидин ингибирует киназу-3β гликогенсинтазы: исследование путем моделирования стыковки и экспериментальной проверки». Журнал ингибирования ферментов и медицинской химии. 28 (4): 690–4. Дои:10.3109/14756366.2012.672413. PMID 22512725. S2CID 11890710.
- ^ Махешвари Р.К., Сингх А.К., Гаддипати Дж., Srimal RC (март 2006 г.). «Множественные биологические активности куркумина: краткий обзор». Науки о жизни. 78 (18): 2081–7. Дои:10.1016 / j.lfs.2005.12.007. PMID 16413584.
- ^ Баласубраманян К. (май 2006 г.). «Молекулярная орбитальная основа для предотвращения болезни Альцгеймера желтым карри куркумином». Журнал сельскохозяйственной и пищевой химии. 54 (10): 3512–20. Дои:10.1021 / jf0603533. PMID 19127718.
- ^ Пэйтон Ф., Сандаски П., Олворт В.Л. (февраль 2007 г.). «ЯМР-исследование структуры раствора куркумина». Журнал натуральных продуктов. 70 (2): 143–6. Дои:10.1021 / np060263s. PMID 17315954.
- ^ Кохли К., Али Дж., Ансари М.Дж., Рахеман З. (2005). «Куркумин: натуральное противовоспалительное средство». Индийский журнал фармакологии. 37 (3): 141. Дои:10.4103/0253-7613.16209.
- ^ Неги П.С., Джаяпракаша Г.К., Джаган Мохан Рао Л., Сакария К.К. (октябрь 1999 г.). «Антибактериальная активность масла куркумы: побочный продукт производства куркумина». Журнал сельскохозяйственной и пищевой химии. 47 (10): 4297–300. Дои:10.1021 / jf990308d. PMID 10552805.
- ^ Сидху Г.С., Сингх А.К., Тхалор Д., Банаудха К.К., Патнаик Г.К., Шримал Р.К., Махешвари Р.К. (1998). «Улучшение заживления ран куркумином у животных». Ремонт и регенерация ран. 6 (2): 167–77. Дои:10.1046 / j.1524-475X.1998.60211.x. PMID 9776860. S2CID 21440334.
- ^ Ян Ф., Лим Г.П., Бегум А.Н., Убеда О.Дж., Симмонс М.Р., Амбегаокар С.С., Чен П.П., Кайед Р., Глабе К.Г., Фраутши С.А., Коул Г.М. (февраль 2005 г.). «Куркумин подавляет образование бета-амилоидных олигомеров и фибрилл, связывает бляшки и снижает уровень амилоида in vivo». Журнал биологической химии. 280 (7): 5892–901. Дои:10.1074 / jbc.M404751200. PMID 15590663.
- ^ Мишра С., Кармодия К., Суролия Н., Суролия А (март 2008 г.). «Синтез и исследование новых аналогов куркумина в качестве противомалярийных агентов». Биоорганическая и медицинская химия. 16 (6): 2894–902. Дои:10.1016 / j.bmc.2007.12.054. PMID 18194869.
- ^ Шарма С., Кулкарни С.К., Чопра К. (октябрь 2006 г.). «Куркумин, активное начало куркумы (Curcuma longa), улучшает диабетическую нефропатию у крыс». Клиническая и экспериментальная фармакология и физиология. 33 (10): 940–5. Дои:10.1111 / j.1440-1681.2006.04468.x. PMID 17002671. S2CID 25193929.
- ^ Демарчи Ф., Бертоли С., Сэнди П., Шнайдер С. (октябрь 2003 г.). «Киназа-3 бета гликоген-синтазы регулирует стабильность NF-каппа B1 / p105». Журнал биологической химии. 278 (41): 39583–90. Дои:10.1074 / jbc.M305676200. PMID 12871932.
- ^ а б Бустанджи Й., Таха М.О., Алмасри И.М., Аль-Гуссейн М.А., Мохаммад М.К., Алхатиб Х.С. (июнь 2009 г.). «Ингибирование киназы гликогенсинтазы куркумином: исследование путем моделирования молекулярного стыковки и последующей оценки in vitro / in vivo». Журнал ингибирования ферментов и медицинской химии. 24 (3): 771–8. Дои:10.1080/14756360802364377. PMID 18720192. S2CID 23137441.
- ^ «Антипсихотики от А до Я». Mind.org.uk. 2018 г.[ненадежный медицинский источник? ]
- ^ «Антипсихотические препараты от биполярного расстройства». WebMD.
- ^ Гоуди А.Дж., Смит Дж. А., Халфорд Дж. С. (декабрь 2002 г.). «Характеристика увеличения веса крыс, вызванного оланзапином». Журнал психофармакологии. 16 (4): 291–6. Дои:10.1177/026988110201600402. PMID 12503827. S2CID 23589812.
- ^ Ди Лоренцо Р., Брогли А. (сентябрь 2010 г.). «Профиль инъекции оланзапина пролонгированного действия для поддерживающего лечения взрослых пациентов с шизофренией». Психоневрологические заболевания и лечение. 6: 573–81. Дои:10.2147 / NDT.S5463. ЧВК 2938306. PMID 20856920.
- ^ Мерфи Ф, Миддлтон (2012). «Цитостатические и цитотоксические препараты». Ежегодный всемирный обзор новых данных о побочных реакциях и взаимодействиях с лекарственными средствами. Побочные эффекты лекарств Ежегодник. 34. С. 731–747. Дои:10.1016 / B978-0-444-59499-0.00045-3. ISBN 9780444594990.
- ^ Крамер Т., Шмидт Б., Ло Монте Ф (2012). «Ингибиторы малых молекул GSK-3: структурные идеи и их применение в моделях болезни Альцгеймера». Международный журнал болезни Альцгеймера. 2012: 381029. Дои:10.1155/2012/381029. ЧВК 3408674. PMID 22888461.
внешняя ссылка
- Гликоген-синтаза-киназа 3 в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)